Nationwide seroprevalence of Mycobacterium bovis and Mycobacterium avium in domestic sows and wild boars in Korea under a one health framework
Seon Jae Moon, Da-Yun Bae, Yun-Chae Cho, Dae Sung Yoo, Yeonsu Oh, Ho-Seong Cho

TL;DR
This study estimates the spread of Mycobacterium bovis and Mycobacterium avium in domestic pigs and wild boars across Korea, highlighting the need for integrated health monitoring.
Contribution
The study provides the first nationwide seroprevalence data for mycobacteria in Korean suids under a One Health framework.
Findings
M. bovis seroprevalence was 4.54% in domestic sows and 5.91% in wild boars.
M. avium seroprevalence was higher in domestic sows (10.10%) than in wild boars (7.71%).
Significant provincial variation was observed only for M. avium in domestic sows.
Abstract
Tuberculosis caused by Mycobacterium bovis and infections due to nontuberculous mycobacteria, particularly the Mycobacterium avium complex (MAC), are increasingly recognized at the livestock–wildlife–human interface. In the Republic of Korea, bovine tuberculosis remains endemic in cattle, yet nationwide data on mycobacterial exposure in suids are lacking. Between February 2023 and November 2024, serum samples from 1,366 domestic sows and 1,168 wild boars collected across nine administrative provinces were analyzed using validated commercial ELISAs to estimate apparent seroprevalence. Apparent seroprevalence of M. bovis was 4.54% (95% CI: 3.56%–5.78%) in domestic sows and 5.91% (95% CI: 4.69%–7.41%) in wild boars. Apparent seroprevalence of M. avium was 10.10% (95% CI: 8.61%–11.81%) in domestic sows and 7.71% (95% CI: 6.31%–9.38%) in wild boars. Significant provincial variation was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
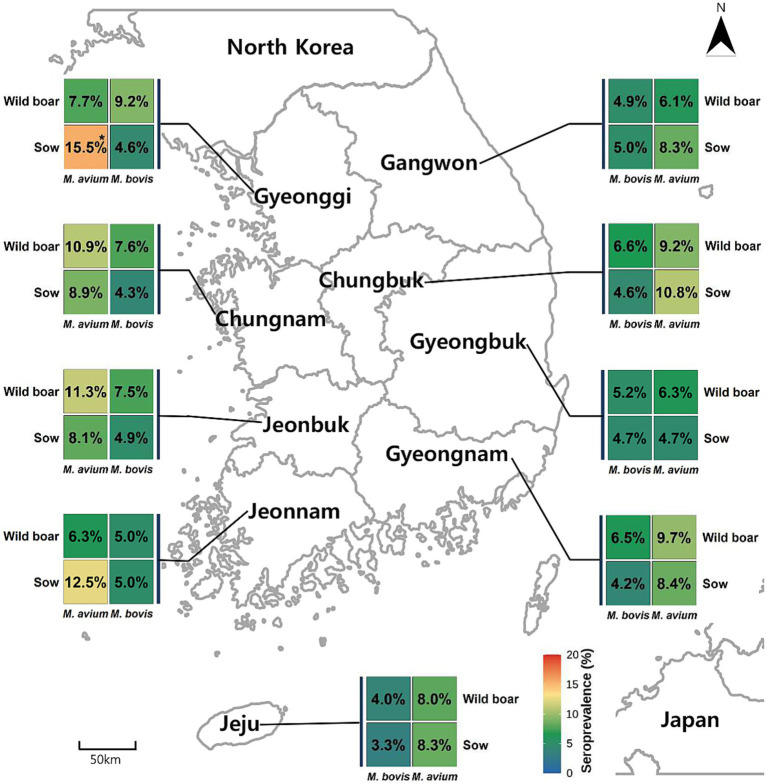 Figure 1
Figure 1| Provinces | Domestic sows | Wild boars | ||
|---|---|---|---|---|
| Minimum sample size | Estimated population | Minimum sample size | Estimated population | |
| Gyeonggi | 171 | 156,105 | 51 | 3,573 |
| Gangwon | 46 | 42,343 | 231 | 16,157 |
| Chungbuk | 57 | 52,371 | 65 | 4,566 |
| Chungnam | 226 | 207,557 | 83 | 5,820 |
| Jeonbuk | 127 | 115,733 | 63 | 4,381 |
| Jeonnam | 116 | 105,979 | 149 | 10,418 |
| Gyeongbuk | 140 | 127,697 | 262 | 18,246 |
| Gyeongnam | 133 | 121,241 | 145 | 10,142 |
| Jeju | 50 | 45,974 | 3 | 221 |
| Total | 1,066 | 975,000 | 1,052 | 73,523 |
| Region (Province) | No. of Positive sows / tested | Apparent prevalence (%) (95% CI) | No. of positive pig farms / tested | Apparent prevalence (%) (95% CI) | No. of positive wild boars / tested | Apparent prevalence (%) (95% CI) |
|---|---|---|---|---|---|---|
| Gyeonggi | 14/303 | 4.62 (2.77–7.61) | 11/92 | 11.96 (6.81–20.15) | 6/65 | 9.23 (4.30–18.71) |
| Gangwon | 3/60 | 5.00 (1.71–13.70) | 2/18 | 11.11 (3.10–32.80) | 12/245 | 4.90 (2.82–8.36) |
| Chungbuk | 3/65 | 4.62 (1.58–12.71) | 2/19 | 10.53 (2.94–31.39) | 5/76 | 6.58 (2.84–14.49) |
| Chungnam | 12/280 | 4.29 (2.47–7.34) | 9/78 | 11.54 (6.19–20.50) | 7/92 | 7.61 (3.73–14.88) |
| Jeonbuk | 9/185 | 4.86 (2.58–8.99) | 6/53 | 11.32 (5.28–22.34) | 6/80 | 7.50 (3.48–15.41) |
| Jeonnam | 6/120 | 5.00 (2.31–10.48) | 4/35 | 11.43 (4.54–25.95) | 8/160 | 5.00 (2.56–9.56) |
| Gyeongbuk | 7/150 | 4.67 (2.28–9.32) | 5/46 | 10.87 (4.74–23.06) | 14/270 | 5.19 (3.11–8.51) |
| Gyeongnam | 6/143 | 4.20 (1.94–8.85) | 5/45 | 11.11 (4.87–23.63) | 10/155 | 6.45 (3.54–11.47) |
| Jeju | 2/60 | 3.33 (0.92–11.36) | 2/20 | 10.00 (2.79–30.10) | 1/25 | 4.00 (0.71–19.54) |
| Total | 62/1,366 | 4.54 (3.56–5.78) | 46/406 | 11.33 (8.62–14.77) | 69/1,168 | 5.91 (4.69–7.41) |
| Region (Province) | No. of positive Sows / Tested | Apparent prevalence (%) (95% CI) | No. of positive pig farms / tested | Apparent prevalence (%) (95% CI) | No. of positive wild boars / tested | Apparent prevalence (%) (95% CI) |
|---|---|---|---|---|---|---|
| Gyeonggi | 47/303 | 15.51* (11.87–20.02) | 24/92 | 26.09 (18.23–35.86) | 5/65 | 7.69 (3.33–16.78) |
| Gangwon | 5/60 | 8.33 (3.61–18.07) | 4/18 | 22.22 (9.00–45.23) | 15/245 | 6.12 (3.75–9.85) |
| Chungbuk | 7/65 | 10.77 (5.32–20.06) | 4/19 | 21.05 (8.55–42.42) | 7/76 | 9.21 (4.53–17.81) |
| Chungnam | 25/280 | 8.93 (6.12–12.85) | 14/78 | 17.95 (11.01–27.83) | 10/92 | 10.87 (6.01–18.86) |
| Jeonbuk | 15/185 | 8.11 (4.97–12.95) | 8/53 | 15.09 (7.84–26.90) | 9/80 | 11.25 (6.03–20.02) |
| Jeonnam | 15/120 | 12.50 (7.72–19.60) | 6/35 | 17.14 (8.09–32.66) | 10/160 | 6.25 (3.43–11.12) |
| Gyeongbuk | 7/150 | 4.67 (2.28–9.32) | 7/46 | 15.22 (7.61–28.35) | 17/270 | 6.30 (3.97–9.85) |
| Gyeongnam | 12/143 | 8.39 (4.87–14.09) | 8/45 | 17.78 (9.16–31.83) | 15/155 | 9.68 (5.95–15.35) |
| Jeju | 5/60 | 8.33 (3.61–18.07) | 3/20 | 15.00 (5.23–36.03) | 2/25 | 8.00 (2.22–24.97) |
| Total | 138/1,366 | 10.10 (8.61–11.81) | 78/406 | 19.21 (15.71–23.25) | 90/1,168 | 7.71 (6.31–9.38) |
| Country | Year | Sampling time | No. positives | No. tested | Prevalence | Detection method | Species |
|---|---|---|---|---|---|---|---|
| Korea (present study) | 2024 | 2023.02–2024.11 | 62 (sows); 69 (wild boars) | 1,366 (sows); 1,168 (wild boars) | 0.045/0.059 | ELISA | Pigs / Wild boars |
| Africa | |||||||
| Burkina Faso ( | 2021 | 2017.08–2017.12 | 8 | 2,430 | 0.003 | Bacteriology | Pigs |
| Egypt ( | 2009 | 2004.04–2005.10 | 30 | 745 | 0.040 | PCR | Pigs |
| Ethiopia ( | 2013 | 2011.03–2011.09 | 12 | 841 | 0.014 | Bacteriology | Pigs |
| Ethiopia ( | 2020 | 2016.09–2017.12 | 10 | 329 | 0.030 | Tuberculin skin test | Pigs |
| Morocco ( | 2016 | 2009–2011 | 6 | 43 | 0.140 | PCR | Wild boars |
| South Africa ( | 2018 | 2013–2015 | 64 | 170 | 0.376 | ELISA | Warthogs |
| South Africa ( | 2021 | 1999.05–2016.08 | 40 | 100 | 0.400 | ELISA | Warthogs |
| South Africa ( | 2024 | Unclear | 0 | 90 | 0 | PCR | Pigs |
| Uganda ( | 2010 | 2008.09–2009.02 | 32 | 997 | 0.032 | Bacteriology | Pigs |
| Asia | |||||||
| China ( | 2024 | 2021.02–2021.06 | 3 | 1,379 | 0.002 | ELISA | Pigs |
| India ( | 2011 | 2008.05–2010.02 | 9 | 108 | 0.083 | Bacteriology | Pigs |
| India ( | 2021 | 2017.04–2020.02 | 3 | 42 | 0.071 | Bacteriology | Pigs |
| Malaysia ( | 2021 | Unclear | 5 | 30 | 0.167 | ELISA | Wild boars |
| Malaysia ( | 2021 | 2019.04–2020.08 | 9 | 30 | 0.300 | PCR | Wild boars |
| Europe | |||||||
| France ( | 2019 | 2014–2016 | 8 | 495 | 0.016 | Bacteriology | Wild boars |
| Italy ( | 2012 | 2009 | 4 | 119 | 0.034 | PCR | Pigs |
| Italy ( | 2017 | 2013 | 24 | 299 | 0.080 | PCR | Pigs |
| Italy ( | 2020 | 2016–2017 | 46 | 434 | 0.106 | ELISA | Wild boars |
| Poland ( | 2020 | 2011–2017 | 21 | 55 | 0.382 | PCR | Wild boars |
| Poland ( | 2023 | 2013–2020 | 46 | 104 | 0.442 | PCR | Wild boars |
| Portugal ( | 2009 | 2005–2007 | 18 | 162 | 0.111 | Bacteriology | Wild boars |
| Portugal ( | 2017 | 2011.06.01–2014.05.31 | 191 | 2,191 | 0.087 | PCR | Wild boars |
| Portugal ( | 2018 | 2006–2013 | 16 | 678 | 0.024 | ELISA | Wild boars |
| Portugal ( | 2024 | Unclear | 13 | 211 | 0.062 | PCR | Wild boars |
| Slovenia ( | 2024 | 2016–2017 | 0 | 676 | 0.000 | Bacteriology | Wild boars |
| 2018–2019 | 0 | 132 | 0.000 | ELISA | |||
| Spain ( | 2013 | 2008–2012 | 33 | 1,275 | 0.026 | PCR | Wild boars |
| 22 | 1,057 | 0.021 | ELISA | ||||
| Spain ( | 2014 | 2002.10–2003.2, 2003.10–2004.2, 2004.10–2005.2, 2005.10–2006.2, 2006.10–2007.2, 2008.10–2009.2, 2009.10–2010.2 | 280 | 1,102 | 0.254 | PCR | Wild boars |
| Spain ( | 2014 | 2004–2012 | 103 | 745 | 0.138 | PCR | Wild boars |
| Spain ( | 2017 | 2011–2015 | 329 | 7,676 | 0.042 | PCR | Wild boars |
| Spain ( | 2018 | 2015–2017 | 82 | 3,622 | 0.023 | ELISA | Pigs |
| Spain ( | 2020 | 2015.09–2016.03, 2016.09–2017.03, 2017.09–2018.03, 2018.09–2019.03, 2019.09–2020.03 | 17 | 278 | 0.061 | PCR | Wild boars |
| Spain ( | 2020 | 2010–2016 | 326 | 1902 | 0.171 | ELISA | Wild boars |
| Spain ( | 2021 | 2010–2019 | 10 | 894 | 0.011 | PCR | Wild boars |
| Switzerland ( | 2021 | 2017–2018 | 5 | 176 | 0.028 | PCR | Wild boars |
| North America | |||||||
| U.S. ( | 2011 | 2010.06–2010.09 | 0 | 98 | 0.000 | Histopathology, Bacteriology | Wild boars |
| U.S. ( | 2017 | 2013.10.01–2014.09.30 | 1 | 2,725 | 0.000 | ELISA | Wild boars |
| South America | |||||||
| Brazil ( | 2017 | Unclear | 25 | 80 | 0.313 | PCR | Wild boars |
| Argentina ( | 2024 | Unclear | 34 | 311 | 0.109 | PCR | Wild boars |
| New Zealand ( | 2011 | 1996–2003, 2003–2007 | 293 | 785 | 0.373 | Bacteriology | Wild boars |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTuberculosis Research and Epidemiology · Mycobacterium research and diagnosis · Infectious Diseases and Tuberculosis
Introduction
1
Tuberculosis (TB) caused by Mycobacterium bovis and other members of the Mycobacterium tuberculosis complex (MTC) remains a persistent challenge at the livestock–wildlife–human interface. Although long-standing eradication programs have substantially reduced bovine TB in many regions, MTC continues to circulate in wildlife reservoirs and may spill over into domestic animals, including pigs. Under certain ecological conditions, pigs can function as spillover or bridge hosts within multi-host systems, thereby contributing to environmental persistence and transmission opportunities (Di Marco et al., 2012; Cano-Terriza et al., 2018). Evidence from Mediterranean ecosystems, including Spain and Italy, has demonstrated MTC infection in domestic and wild swine populations, highlighting their epidemiological relevance where livestock, wildlife, and shared environments overlap (Di Marco et al., 2012; Cano-Terriza et al., 2018; Muñoz-Mendoza et al., 2013; Iovane et al., 2020). In contrast, countries dominated by intensive indoor pig production systems typically report low prevalence, emphasizing the influence of production ecology and wildlife contact on exposure risk (Campbell et al., 2011; Pedersen et al., 2017; Sun et al., 2024).
In the Republic of Korea, bovine TB has remained endemic for more than a century, with recent outbreak continuing despite sustained national efforts. In parallel, MTC infections have been documented in suids (Ku et al., 2016; Seo et al., 2017). M. bovis isolated from a wild sow (Sus scrofa) in 2012, exhibited a genotype distinct from strains circulating in cattle and deer, suggesting an independent or previously unrecognized source of exposure (Ku et al., 2016). In the same year, M. tuberculosis infection was confirmed in a domesticated wild boar through PCR and immunohistochemistry (Seo et al., 2017). Although such reports are sporadic, they indicate that domestic pigs and wild boars in Korea may be exposed to MTC and could participate as bridge hosts at the livestock–wildlife interface – an issue central to One Health surveillance.
Beyond MTC, the Mycobacterium avium complex (MAC), comprising M. avium and M. intracellulare, presents additional challenges for veterinary and public health sectors. MAC organisms are widespread in the environment, and pigs are susceptible to infection, often developing granulomatous lesions in mesenteric lymph nodes detected at slaughter. Although infections are frequently subclinical, MAC is associated with carcass condemnation and economic loss and may carry zoonotic implications, particularly for immunocompromised individuals (Hulinova Stromerova and Faldyna, 2018). In Korea, the incidence of MAC pulmonary disease in humans has increased, and M. avium is now frequently isolated in clinical settings (Kwon et al., 2019). Despite these trends, MAC exposure in Korean swine remains poorly characterized, limiting understanding of potential environmental or cross-species exposure routes. Notably, MAC, especially M. avium subsp. hominissuis, has been frequently detected in slaughter pigs and linked to lymph node lesions, underscoring its relevance to herd health and the food chain (Gcebe et al., 2023).
International studies report wide variability in mycobacterial prevalence in domestic pigs and wild suids, reflecting differences in production systems, wildlife reservoirs, and diagnostic approaches. Higher prevalence has been reported in wildlife-rich ecosystems and in settings with outdoor husbandry or established maintenance hosts, whereas low prevalence is typically observed in intensive indoor systems. However, direct comparisons among studies are constrained by methodological heterogeneity, including differences in the use of serology, PCR, culture, lesion inspection, or skin testing.
Taken together, the persistence of bovine TB, the occurrence of MTC infections in suids, and the growing public health significance of NTM – including MAC – highlight the need for integrated surveillance across animal species. Yet, no nationwide serological survey of M. bovis or M. avium exposure in swine has been conducted in Korea. This knowledge gap hampers risk assessment at the livestock–wildlife–human interface.
This study therefore aimed to (i) provide the first nationwide estimates of apparent seroprevalence of M. bovis and M. avium in domestic sows and wild boars in Korea, (ii) evaluate province-level heterogeneity while avoiding inference about spatial clustering beyond the resolution of the data, and (iii) position Korean swine mycobacterial exposure within a global One Health context.
Materials and methods
2
Sample size determination and sample collection
2.1
Minimum sample sizes for domestic sows and wild boars (Sus scrofa) were calculated using population estimates from the 2024 Q2 Livestock Trend Survey (Statistics Korea, 2025) and the 2023 Wildlife Survey (National Institute of Biological Resources, 2023), respectively. The source population was estimated at approximately 975,000 domestic sows and 73,523 wild boars (Table 1).
Sample size calculations were performed in R version 4.3.2 (R Studio 2023.09.01), assuming an expected prevalence (P) of 0.5, a 95% confidence level (Z = 1.96), and a margin of error (d) of 0.03. A stricter margin of error than the conventional 0.05 was selected to increase the minimum required sample size and thereby improve the precision of national prevalence estimates. At the study design stage, diagnostic sensitivity and specificity were not incorporated into the sample size formula, as reliable field-validated estimates for the commercial ELISA assays in domestic pigs and wild boars under low-prevalence conditions are currently limited.
An initial sample size assuming an infinite population was calculated as:
The final required sample size (n) for each species was then obtained using finite population correction:
where N denotes the estimated source population size. Under these assumptions, the minimum required sample size was 1,066 domestic sows and 1,052 wild boars. Final target sample sizes were proportionally allocated across nine administrative provinces according to regional population distributions (Table 1).
Between February 2023 and November 2024, serum samples were obtained from 1,366 domestic sows and 1,168 wild boars across nine provinces. For domestic sows, approximately 5 mL of blood was collected via jugular venipuncture into serum separator tubes (Ampulab Serum Separation Gel Tube; Soyagreentec, Seoul, Korea). Samples were transported on ice to the Swine Disease Laboratory at the College of Veterinary Medicine, Jeonbuk National University within a maximum of 24 h after collection, centrifuged at 3,000 × g for 10 min, and stored at −20 °C until analysis. Domestic sow samples were collected from breeding herds maintained under intensive indoor production systems. Wild boar samples were obtained post-mortem from animals harvested through routine population management and regulated hunting programs coordinated by local authorities. Blood samples were collected immediately after harvest and processed using the same laboratory procedures as domestic sow samples.
ELISA-based serological detection
2.2
All sera were tested for antibodies to M. bovis and M. avium using commercial ELISA kits (ID Screen® Porcine Tuberculosis Indirect and ID Screen® Mycobacterium avium Indirect Multi-species; Innovative Diagnostics, Grabels, France), following the manufacturer’s instructions. Samples were diluted 1:100 in dilution buffer, and optical densities (OD) were measured at 450 nm.
The sample-to-positive (S/P) ratios were calculated as:
Following the manufacturer’s instructions, S/P ratios ≥ 40 (for M. bovis) or ≥ 50 (for M. avium) were interpreted as positive. Positive and negative control sera supplied with the kit were included on each plate to validate assay performance according to the manufacturer’s quality criteria (OD ratio > 3.0 between controls).
Statistical analyses
2.3
Apparent seroprevalence was calculated as the proportion of ELISA-positive samples among those tested. Exact two-sided 95% confidence intervals (CIs) were computed using the Clopper–Pearson method (binom.test in R), which is conservative and suitable for low-prevalence settings.
Provincial differences were assessed using Fisher’s exact test when expected cell counts were sparse (<5), and Pearson’s chi-square test when assumptions were met. Specifically, Fisher’s exact test was applied for M. bovis in sows and for both antigens in wild boars, whereas Pearson’s chi-square test was used for M. avium in sows. When global testing indicated significance (α = 0.05, two-sided), post-hoc cellwise diagnostics were performed using standardized Pearson residuals; |z| ≥ 1.96 was considered noteworthy. p-values were adjusted across provinces (m = 9) using Holm’s method.
Samples with missing or indeterminate ELISA results were excluded under complete-case analysis; however, no samples were excluded for this reason (0 of 2,534 samples). A farm was classified as seropositive if ≥1 sampled sow from that farm tested ELISA-positive. As supplementary summaries, province-specific prevalence ratios with 95% CIs were calculated. True prevalence was not estimated because robust sensitivity and specificity values for these ELISAs in domestic pigs and wild boars under low-prevalence field conditions were not consistently available.
Results
3
Apparent seroprevalence of Mycobacterium bovis antibodies
3.1
Among 1,366 sow sera, 62 tested positive for M. bovis antibodies, yielding an apparent seroprevalence of 4.54% (95% CI: 3.56%–5.78%) (Table 2). Provincial seroprevalence values ranged from 3.33% to 5.00%, with no statistically significant differences detected among provinces (Fisher’s exact test, p > 0.05). The provincial distribution is shown in Figure 1.
Provincial seroprevalence of Mycobacterium bovis and Mycobacterium avium antibodies in domestic sows and wild boars across Korea (2023.02–2024.11). Choropleth maps illustrate the apparent seroprevalence (%) of M. bovis and M. avium in (A) domestic sows (n = 1,366) and (B) wild boars (n = 1,168) across nine administrative provinces of Korea, based on commercial ELISA results (ID Screen® Porcine Tuberculosis Indirect; ID Screen® M. avium Indirect Multi-species). Provincial differences were evaluated using Pearson’s chi-square test or Fisher’s exact test, as appropriate. Significant regional variation was observed only for M. avium seropositivity in domestic sows, with higher-than-expected prevalence in the Gyeonggi region (Holm-adjusted p < 0.01). Provinces with limited wild boar sample sizes (e.g., Jeju) should be interpreted with caution.
Of the 1,168 wild boar samples analyzed, 69 were positive for M. bovis antibodies, resulting in an apparent seroprevalence of 5.91% (95% CI: 4.69%–7.41%) (Table 2). Provincial estimates ranged from 4.00% (Jeju) to 9.23% (Gyeonggi), but no statistically significant provincial differences were observed (Fisher’s exact test, p > 0.05).
Apparent seroprevalence of Mycobacterium avium antibodies
3.2
A total of 138 of 1,366 sows were seropositive for M. avium, corresponding to an apparent seroprevalence of 10.10% (95% CI: 8.61%–11.81%) (Table 3). Chi-square testing revealed significant provincial heterogeneity (χ^2^ = 18.3, df = 8, Holm-adjusted p < 0.05). The Gyeonggi Province exhibited significantly higher seropositivity (15.51%), as supported by standardized residual diagnostics (Holm-adjusted p = 0.007).
In wild boars, 90 of 1,168 samples were seropositive for M. avium, resulting in 7.71% (95% CI: 6.31–9.38%) (Table 3). Provincial prevalence ranged from 6.12% (Gangwon) to 11.25% (Jeonbuk), with no statistically significant provincial variation (Fisher’s exact test, p > 0.05).
Contextual comparison with global prevalence data
3.3
For broader epidemiological context, Table 4 compares prevalence estimates from the present study with reported swine tuberculosis prevalence worldwide, derived using diverse diagnostic methods including ELISA, PCR, bacteriology, and skin testing. Seroprevalence estimates for Korean sows and wild boars fall within the lower-to-middle range of reported international values; however, comparisons should be interpreted cautiously due to methodological heterogeneity across studies.
Discussion
4
This study provides the first nationwide serological assessment of exposure to M. bovis and M. avium in domestic sows and wild boars in Korea. By collecting samples across nine administrative provinces and applying validated ELISA assays, we establish baseline data that contribute to understanding mycobacterial exposure at the livestock–wildlife interface and provide a reference point for future surveillance efforts.
The apparent seroprevalence of M. bovis observed in domestic sows (4.54%) exceeds levels reported from several regions characterized by intensive indoor or semi-intensive pig production systems, including southern Spain (Cano-Terriza et al., 2018). Similarly, the seroprevalence detected in wild boars (5.91%) was higher than estimates reported from Portugal (Santos et al., 2018), the United States (Pedersen et al., 2017), Iberian Atlantic Spain (Muñoz-Mendoza et al., 2013), and Slovenia (Pate et al., 2024). In contrast, the Korean estimates were lower than those reported from multi-host systems in Campania, Italy (Iovane et al., 2020), northern Spain (Varela-Castro et al., 2020), and Malaysia (Lekko et al., 2021), where wildlife–livestock interfaces are known to support sustained transmission. Substantially, higher seroprevalence has been documented in African ecosystems, where warthogs (Phacochoerus africanus) function as maintenance hosts, with reported values reaching 37.6%–40.0% in South Africa (Roos et al., 2018; Neiffer et al., 2021). Collectively, these comparisons suggest that Korea occupies an intermediate epidemiological position between intensive production systems with limited wildlife contact and ecosystems characterized by persistent wildlife reservoirs. Nevertheless, such comparisons should be interpreted cautiously, as reported prevalence is strongly influenced by host populations, ecological context, and diagnostic methodology.
When placed in a broader international context, detection frequencies for M. bovis in suids have generally been low but variable across regions. For example, individual seroprevalence in Gansu Province, China, was reported at 0.22% (Sun et al., 2024), whereas higher detection rates have been described in Mediterranean settings, including 3.4% of slaughtered pigs in Sicily, Italy (Di Marco et al., 2012), and 2.3% seroprevalence among Iberian pigs in southern Spain (Cano-Terriza et al., 2018). For the MAC, reported frequencies likewise vary, ranging from 2.4% in slaughtered pigs in Egypt (Mohamed et al., 2009) to 4.6% in wild boars from Spain’s Iberian Atlantic region (Muñoz-Mendoza et al., 2013) and 1.9% in wild boars from southern Spain (García-Jiménez et al., 2015). Relative to these reports, the apparent seroprevalence observed in Korean domestic sows and wild boars does not indicate unusually high exposure but is consistent with measurable, low-to-moderate exposure levels.
The seroprevalence profile observed in Korea likely reflects the predominance of intensive indoor pig production systems, which limit direct contact between domestic pigs and wildlife, combined with potential environmental exposure arising from other reservoirs. Bovine tuberculosis continues to be reported in Korean cattle herds (Animal and Plant Quarantine Agency, 2025; Yoo et al., 2025), and prior documentation of M. bovis infection in a wild sow (Ku et al., 2016) and M. tuberculosis detection in a wild boar (Seo et al., 2017) supports the plausibility of multi-host exposure pathways. However, serological data alone cannot identify sources of infection or infer transmission routes, and the present findings should therefore be interpreted as evidence of exposure rather than active transmission.
Exposure to M. avium was more common than exposure to M. bovis in both domestic sows and wild boars, with apparent seroprevalence reaching 10.10% and 7.71%, respectively. Direct quantitative comparisons with previous studies are constrained by methodological heterogeneity, as many investigations of MAC in pigs and wild suids rely on culture or molecular detection rather than serology. Nonetheless, international studies demonstrate considerable variability in MAC detection in slaughter pigs and wild boars (Muñoz-Mendoza et al., 2013; Mohamed et al., 2009; García-Jiménez et al., 2015). The veterinary and public health relevance of MAC infections has been increasingly recognized, given their association with carcass condemnation in pigs and the rising incidence of MAC pulmonary disease in humans, including in Korea (Hulinova Stromerova and Faldyna, 2018; Kwon et al., 2019).
In the present study, significant regional variation in M. avium exposure was observed only among domestic sows, with the highest apparent seroprevalence detected in the Gyeonggi region. Studies addressing MAC exposure in domestic pigs or wild boars in Korea and East Asia remain extremely limited, underscoring the lack of regional baseline data. This finding should be interpreted cautiously, as statistical significance was driven by a single province. Gyeonggi Province encompasses the highest density of large-scale swine operations and complex peri-urban interfaces, suggesting that environmental factors such as water systems, bedding materials, or bioaerosols may contribute to elevated exposure risk (Hulinova Stromerova and Faldyna, 2018; Kwon et al., 2019). In contrast, wild boars exhibited no significant regional variation in MAC seroprevalence, consistent with diffuse, low-level environmental exposure rather than localized exposure foci, as reported in European wildlife studies (Muñoz-Mendoza et al., 2013; Richomme et al., 2019; Ghielmetti et al., 2021).
The concurrent detection of antibodies to both MTC and MAC in domestic sows and wild boars highlights several One Health considerations. Environmental and indirect transmission pathways may be sufficient to sustain low-level endemic exposure even in the absence of widespread outdoor pig farming. Korea’s high baseline prevalence of human tuberculosis and the increasing incidence of MAC pulmonary disease may further contribute to shared peri-urban exposure environments (Kwon et al., 2019; Yoo et al., 2025). In addition, other wildlife or meso-mammalian species, including rodents and small carnivores, may participate in local transmission networks, analogous to the role of brushtail possums in New Zealand’s bovine tuberculosis system (Nugent et al., 2012; Nugent et al., 2015). Evidence of mycobacterial infections in Korean companion animals and wildlife—including multidrug-resistant M. bovis isolated from a dog (Cho et al., 2022), M. avium infections in birds (Lee et al., 2010), and disseminated MAC infection in a captive tiger (Cho et al., 2006)—further underscores the relevance of integrated, multi-species surveillance.
Several methodological considerations warrant acknowledgment. ELISA-based assays are well suited for large-scale screening and have been widely applied in livestock and wildlife surveillance (Santos et al., 2018; Richomme et al., 2019; Cardoso et al., 2024); however, they primarily reflect prior exposure rather than active infection and may under-detect early or low-antibody responses. Accordingly, the apparent seroprevalence reported here should be interpreted as indicative of exposure rather than infection status. Reliable estimation of true prevalence would require robust, species- and context-specific diagnostic performance parameters that are currently unavailable for ELISA-based assays in suids. Complementary diagnostic approaches, including PCR, bacterial culture, interferon-gamma release assays, and histopathology, would improve epidemiological resolution (Mohamed et al., 2009; Cardoso et al., 2024).
The absence of demographic information for wild boars, such as age and sex, precluded assessment of risk factors known to influence infection dynamics in other settings (Muwonge et al., 2010; Arega et al., 2013; Demissie et al., 2020; Sanou et al., 2021). Similarly, individual- or farm-level risk factor analyses were not possible due to limited metadata on herd characteristics and management practices. The lack of spatial analysis further restricted identification of potential geographic clusters or transmission hotspots. Future studies integrating structured metadata, environmental variables, and molecular typing approaches – including spoligotyping, variable-number tandem repeat analysis, or whole-genome sequencing – would be instrumental in clarifying transmission pathways across hosts, as demonstrated in European multi-host systems (Amato et al., 2017; Madeira et al., 2017; Orłowska et al., 2020; Ciaravino et al., 2021; Welz et al., 2023).
Although tuberculosis is designated as a “Contagious Animal Disease Type II” under Korean legislation (Yoo et al., 2025), pigs are not currently included in routine national surveillance programs. Given the long production lifespan of breeding sows and the expanding geographic distribution of wild boars in Korea (National Institute of Biological Resources, 2023), integrating serological monitoring of both populations into existing surveillance frameworks could strengthen early detection and risk assessment. Experience from European wildlife–livestock tuberculosis systems demonstrates the value of coordinated surveillance, ecological risk evaluation, and targeted control strategies (Santos et al., 2009; Cano-Manuel et al., 2014; Mentaberre et al., 2014; Gortázar et al., 2017). Expanded environmental sampling and molecular epidemiological investigations would further clarify transmission pathways across species and ecosystems.
In conclusion, domestic sows and wild boars in Korea exhibit measurable exposure to both MTC and MAC. The observed seroprevalence profile lies between that reported for intensive indoor production systems and that observed in outdoor or wildlife-rich ecosystems with established reservoir hosts (Di Marco et al., 2012; Cano-Terriza et al., 2018; Lekko et al., 2021; Roos et al., 2018; Neiffer et al., 2021; Santos et al., 2009; Maciel et al., 2018; Barandiaran et al., 2024). These findings support the importance of sustained One Health surveillance and integrated multi-species monitoring to better understand and mitigate mycobacterial exposure risk in Korea.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amato B. Capucchio T. M. Biasibetti E. Mangano E. Boniotti B. M. Pacciarini L. M. . (2017). Pathology and genetic findings in a rare case of Mycobacterium caprae infection in a sow. Vet. Microbiol. 205, 71–74. doi: 10.1016/j.vetmic.2017.05.010, 28622866 · doi ↗ · pubmed ↗
- 2Animal and Plant Quarantine Agency. Korea animal health integrated system (KAHIS): bovine tuberculosis occurrence status. (2025). Available online at: https://home.kahis.go.kr/home/lkntscrinfo/select Lknts Occrrnc.do [Accessed August 12, 2025].
- 3Arega S. M. Conraths F. J. Ameni G. (2013). Prevalence of tuberculosis in pigs slaughtered at two abattoirs in Ethiopia and molecular characterization of Mycobacterium tuberculosis isolated from tuberculous-like lesions in pigs. BMC Vet. Res. 9:97. doi: 10.1186/1746-6148-9-97, 23647845 PMC 3661388 · doi ↗ · pubmed ↗
- 4Barandiaran S. Marfil M. J. La Sala L. F. Tammone A. Condori W. E. Winter M. . (2024). Tuberculosis in wild pigs from Argentina. Eco Health 21, 71–82. doi: 10.1007/s 10393-024-01681-y, 38727761 · doi ↗ · pubmed ↗
- 5Campbell T. A. Long D. B. Bazan L. R. Thomsen B. V. Robbe-Austerman S. Davey R. B. . (2011). Absence of Mycobacterium bovis in feral swine (Sus scrofa) from the southern Texas border region. J. Wildl. Dis. 47, 974–978. doi: 10.7589/0090-3558-47.4.974, 22102669 · doi ↗ · pubmed ↗
- 6Cano-Manuel F. J. López-Olvera J. Fandos P. Soriguer R. C. Pérez J. M. Granados J. E. (2014). Long-term monitoring of 10 selected pathogens in wild boar (Sus scrofa) in Sierra Nevada National Park, southern Spain. Vet. Microbiol. 174, 148–154. doi: 10.1016/j.vetmic.2014.06.01725261921 · doi ↗ · pubmed ↗
- 7Cano-Terriza D. Risalde M. A. Rodríguez-Hernández P. Napp S. Fernández-Morente M. Moreno I. . (2018). Epidemiological surveillance of Mycobacterium tuberculosis complex in extensively raised pigs in the south of Spain. Prev. Vet. Med. 159, 87–91. doi: 10.1016/j.prevetmed.2018.08.015, 30314795 · doi ↗ · pubmed ↗
- 8Cardoso B. Jiménez-Ruiz S. Perelló Jiménez A. Nóvoa M. Santos J. P. V. Correia-Neves M. . (2024). Performance of post-mortem diagnostic tests for tuberculosis in wild ungulates at low and high prevalence assessed using Bayesian latent class models. Front Vet Sci. 11:1415277. doi: 10.3389/fvets.2024.1415277, 39386245 PMC 11461446 · doi ↗ · pubmed ↗