Structural and functional studies of the EGF20-27 region reveal new features of the human Notch receptor important for optimal activation
Zhihan Bo, Thomas Rowntree, Steven Johnson, Hilman Nurmahdi, Richard J. Suckling, Johan Hill, Boguslawa Korona, Philip C. Weisshuhn, Devon Sheppard, Yao Meng, Shaoyan Liang, Edward D. Lowe, Susan M. Lea, Christina Redfield, Penny A. Handford

TL;DR
This study reveals that a specific region of the Notch receptor relies on calcium to maintain a rigid structure, which is important for proper signaling.
Contribution
The study identifies a Ca2+-dependent structural organization in the EGF20-27 region of Notch-1 and its functional role in receptor activation.
Findings
The EGF20-27 region of Notch-1 is rigid and elongated with high calcium affinity in most domains.
Mutations in Ca2+-binding sites reduce trans-activation and cis-inhibition, showing the importance of calcium-stabilized structure.
The Ax region's Ca2+-dependent structure influences Notch signaling efficiency and cis-inhibition balance.
Abstract
The Notch receptor is activated by the Delta/Serrate/Lag-2 (DSL) family of ligands. The organization of the extracellular signaling complex is unknown, although structures of Notch/ligand complexes comprising the ligand-binding region (LBR), and negative regulatory region (NRR) region, have been solved. Here, we investigate the human Notch-1 epidermal growth factor-like (EGF) 20-27 region, located between the LBR and NRR, and incorporating the Abruptex (Ax) region, associated with distinctive Drosophila phenotypes. Our analyses, using crystallography, NMR and small angle X-ray scattering (SAXS), support a rigid, elongated organization for EGF20-27 with the EGF20-21 linkage showing Ca2+-dependent flexibility. In functional assays, Notch-1 variants containing Ax substitutions result in reduced ligand-dependent trans-activation. When cis-JAG1 was expressed, Notch activity differences…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
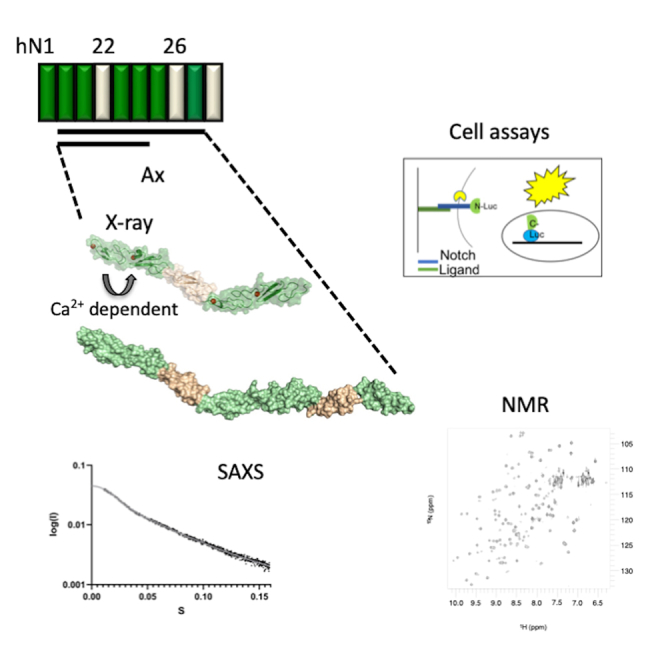 Figure 1
Figure 1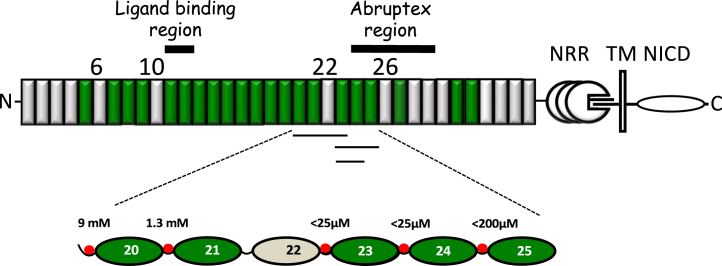 Figure 2
Figure 2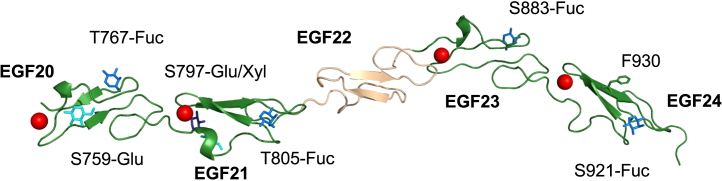 Figure 3
Figure 3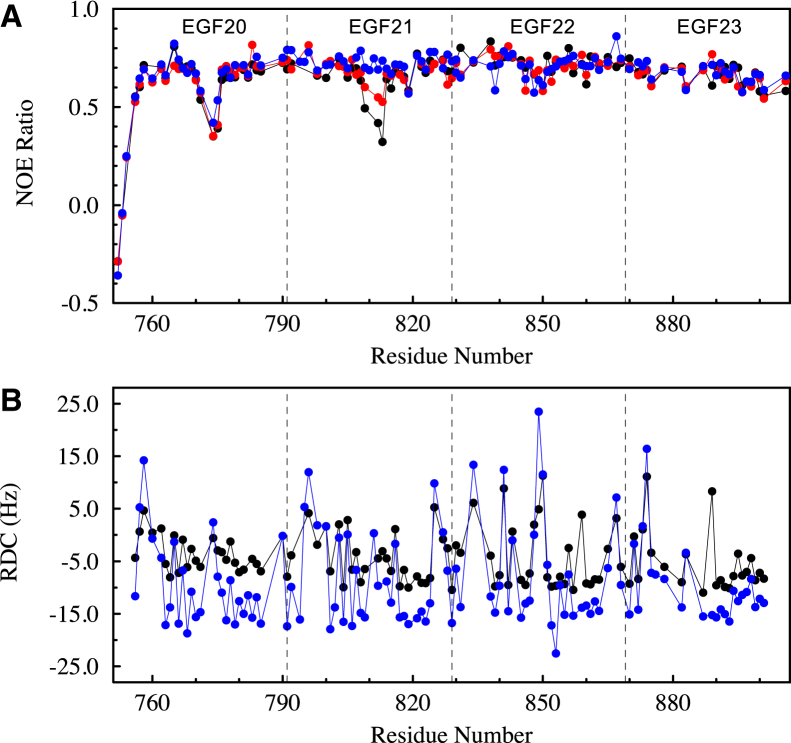 Figure 4
Figure 4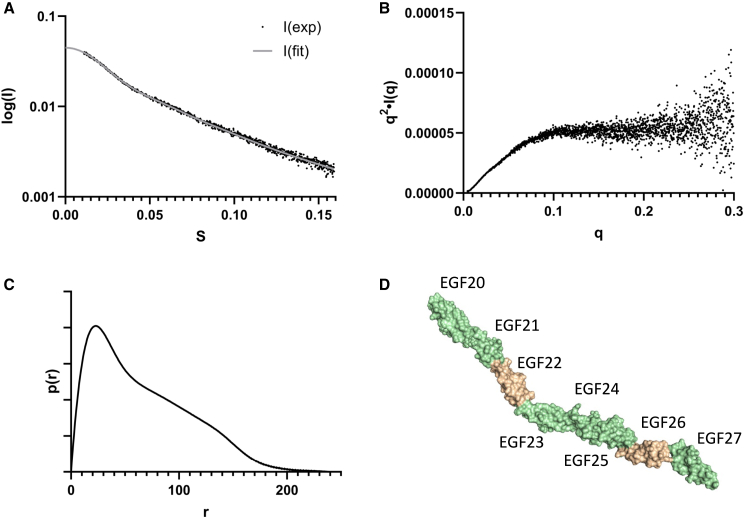 Figure 5
Figure 5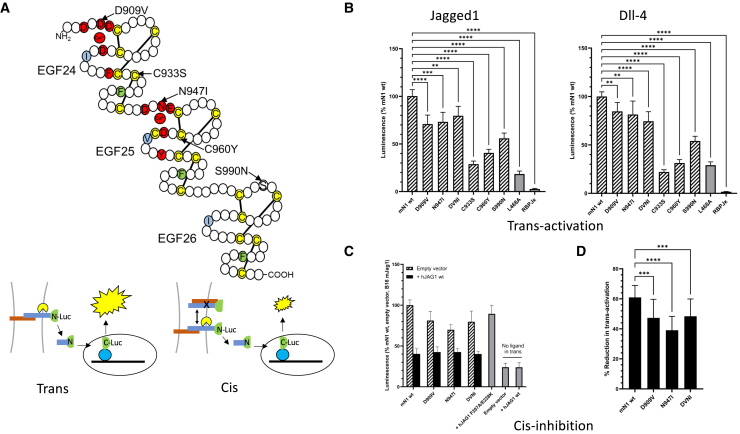 Figure 6
Figure 6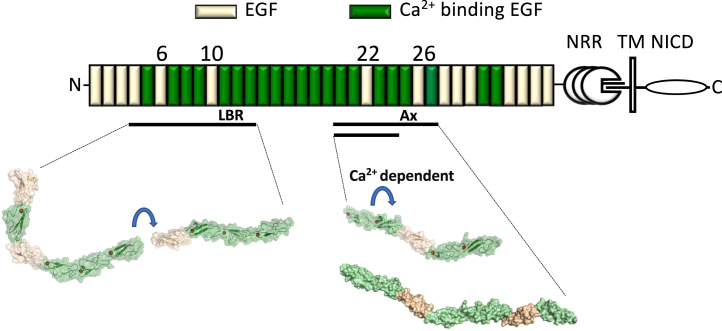 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProtein Tyrosine Phosphatases · Developmental Biology and Gene Regulation · Congenital heart defects research
Introduction
The Notch receptor is part of a core metazoan signal transduction pathway which performs crucial roles during development and in the adult organism including cell-fate determination, cell proliferation, and apoptosis and has an impact on most tissues and organs.1^,^2 In adults, Notch has key roles in tissue homeostasis by regulating stem cell maintenance and function, immune system activation, and angiogenesis. Dysregulation of the Notch pathway results in both inherited and acquired disease, including many cancers, but also occurs during aging in normal esophageal epithelial tissue where hNotch-1 mutations drive clonal expansion and appear protective against tumorigenesis.3^,^4
Canonical Notch signaling requires cell-surface expression of a hetero-dimeric trans-membrane receptor which is extensively modified by O-glucosylation and O-fucosylation.5^,^6^,^7^,^8 Ligand binding by one of the Jagged/Serrate or Delta (DLL) families, followed by ligand endocytosis, exposes the S2 site within the negative regulatory region (NRR) of Notch to proteolytic cleavage by ADAM10. Following this, γ secretase cleaves at the intramembrane S3 site releasing the intracellular domain of Notch (NICD).9^,^10 Subsequently, NICD translocates to the nucleus, and in the presence of mastermind-like (MAML) binds to a transcription factor of the CBF1, suppressor of hairless, Lag-1 (CSL/RBPJk) family, and relieves repression of genes of the HES and Hey families.11 Notch ligand activity is sensitive to the modification of O-fucosylated Notch by Fringe.12^,^13^,^14^,^15 This can modulate signaling by different ligands important in controlling embryonic patterning and boundary formation between adjacent developmental compartments.
Although the downstream consequences of Notch receptor activation have been extensively studied, there is less knowledge about the initial cell-cell mediated extracellular Notch/ligand binding event and how this is converted into an activating signal. Identification of an N-terminal C2 domain in the canonical ligands suggested lipid binding at the cell surface may be important for optimal activation.16^,^17^,^18 Different activation dynamics which result in opposing effects on myogenic cell fate were shown to be mediated by DLL1 and DLL4 signaling.19 Data have also shown cis-activation to be an additional mode of Notch signaling together with cis-inhibition and trans-activation.20 These and other studies underscore the importance of understanding the pleiotropy of receptor/ligand structures and interactions at the cell surface and their subsequent fate.
High-resolution structures of receptor and ligand fragments, both in isolation and in complex, have been determined giving insight into the core interaction surfaces and the role of O-glycans.16^,^18^,^21^,^22^,^23^,^24^,^25^,^26 Further in vivo and in vitro work has begun to decipher the complex Fringe code and how this regulates Notch EGF domain sensitivity to different ligands.15^,^27 The structure of the NRR has been determined establishing its mechanosensory role in Notch signal activation which regulates S2 cleavage.28^,^29 However, there has been much less structural information available for other regions of the extracellular portion of Notch, which are mainly comprised of multiple tandem repeats of EGF domains (Figure 1). We previously identified both flexible and rigid EGF-like domain interfaces in the N-terminal EGF4-13 extracellular region of hNotch-1 which binds ligand.30 These latter data reveal that this portion of the receptor does not simply extend from the cell surface in a near-linear rigid fashion, as once predicted, but has the potential to adopt a range of conformations. This is supported by a mass spectrometry/small angle X-ray scattering (SAXS) study which showed that Notch and ligand ectodomains can form novel intra- and inter-molecular interactions, incompatible with a linear rod-like structure.31Figure 1. Modular organization of the extracellular domain of hNotch-1 and overview of Ca^2+^ dissociation constants(Upper panel) The negative regulatory region (NRR) and transmembrane domain (TM) of hNotch-1 are indicated. Individual domains belonging to the Notch intracellular domain (NICD) are not indicated separately. Ca^2+^-binding and non-Ca^2+^-binding EGF domains are indicated in green and wheat, respectively. The thick horizontal black lines highlight the ligand-binding region, EGF11-13, and the Abruptex region, EGF24-29. The thin black lines indicate the constructs used for Ca^2+^ dissociation constant determination (EGF20-23, EGF23-25, EGF23-24).(Lower panel) The measured Ca^2+^ dissociation constants at pH 7.5 and I = 0.15 for the EGF20-25 region are shown. K_D_ values were determined by NMR. Ca^2+^ is indicated by a red circle at the N terminus of each Ca^2+^-binding EGF domain. The weak affinity for EGF20 reflects its non-native context in EGF20-23 where it lacks a preceding EGF domain which would be expected to stabilize the Ca^2+^-bound state (also see Figures S1–S4).
In this study, we have used X-ray crystallography, NMR spectroscopy, and SAXS to investigate the structure and flexibility of the EGF20-27 region of the hNotch-1 ectodomain by analyzing a series of limited fragments (Figure 1). This region is homologous to Drosophila Notch where Abruptex (Ax) missense mutations in the EGF24-29 region result in specific phenotypes which are distinct from Notch null or loss of function phenotypes.32^,^33^,^34^,^35 We report crystal structures of EGF21-23 and EGF20-24, where the domain interface formed between the non-Ca^2+^-binding domain, EGF22, and its preceding domain introduces a near-linear conformation to the region. Ca^2+^-binding measurements identify high-affinity sites (K_D_<25 μM) for EGF23 and 24 domains in the 20–24 region and an unexpectedly low affinity site in EGF21 (K_D_∼1.3 mM). The dynamic behavior of the EGF20-21 interface has been probed as a function of Ca^2+^ concentration using {^1^H}-^15^N heteronuclear Nuclear Overhauser Effect (NOE) measurements and residual dipolar couplings; these data suggest that changes in local Ca^2+^ concentrations may modulate the flexibility of this region. Data from SAXS analysis of the EGF20-27 construct have been used, together with the EGF20-24 X-ray structure, to model the overall topology of this region which appears rigid and elongated. In parallel, functional studies of full-length Notch-1 variants revealed lower ligand-mediated Notch trans-activation in cellular assays either due to defective Ca^2+^ binding to EGF24 and EGF25 or misfolding. Observed differences in ligand-dependent activation of Ca^2+^ binding Notch variants and WT were suppressed when JAG1 was expressed in cis, within the same cell as receptor. These data are consistent with an important role for the Ca^2+^-dependent structure of the EGF20-27 region in facilitating ligand-dependent activation and cis-inhibition and provide a plausible explanation for a subset of unusual Abruptex phenotypes.
Results
Crystal structures of hNotch-1 EGF21-23 and EGF20-24 reveal a near-linear conformation
Structures of hNotch-1 EGF21-23 and hNotch-1 EGF20-24 were determined using X-ray crystallography to resolutions of 1.55 Å and 1.50 Å respectively (Table 1). The structures are elongated, with each domain within the construct displaying a canonical EGF fold (Figure 2). Tilt and twist angles between adjacent domains pairs in hNotch-1 EGF20-24 are listed in Table 2, and hNotch-1 EGF21-23 superposes onto hNotch-1 EGF20-24 with an r.m.s.d < 1 Å (over 111 C_α_ atoms).Table 1. Crystallization and structure determinationhNotch-1 EGF21-23hNotch-1 EGF20-24Data collectionBeamlineDiamond I04-1Diamond I03Space groupP4_1_P1Wavelength (Å)0.920.9763Cell dimensions (Å)a, b, c (Å)41.9, 41.9, 61.228.0, 35.1, 63.9α, β, γ (°)90.0, 90.0, 90.099.7, 92.4, 96.9Resolution range (Å)a41.92–1.55 (1.59–1.55)62.9–1.50 (1.53–1.50)Unique reflections15359 (1109)36167 (2764)R_merge_a^,^b0.022 (0.497)0.056 (0.483)R_meas_a^,^c0.040 (0.667)0.068 (0.602)CC_1/2_a^,^d0.999 (0.834)0.996 (0.884)Mean I/σIa27.1 (2.5)7.7 (1.7)Completeness (%)a99.7 (99.5)95.2 (94.4)Multiplicitya4.7 (4.7)2.6 (2.6)Anomalous completeness93.8 (93.1)–Anomalous multiplicity2.2 (2.1)–Anomalous correlation0.721 (0.028)–Wilson <B> (Å^2^)27.720.7RefinementResolution range (Å)a41.9–1.55 (1.67–1.55)62.9–1.50 (1.54–1.50)No. of reflections1533136167R_work_/R_free_a0.195/0.218 (0.316/0.366)0.175/0.197 (0.294/0.319)Number of atoms/B-factors (Å^2^)Protein841/45.61439/32.6Ligand/ion2/24.083/42.3Water93/50.4288/42.5Rmsd from ideal valuesBond lengths (Å)0.0040.014Bond angles (°)0.6701.46Ramachandran plotFavored region (%)99.197.9Allowed (%)100.0100.0Outliers (%)00Rotamer outliers (%)1.00.6C-beta outliers00PDB ID code9B3G9B3NaValues in parentheses are for the highest resolution shell.bRmerge = Σ(Ihl - <Ih>)/Σ(Ihl) where <Ih> is the mean intensity of unique reflection h, summed over all reflections for each observed intensity Ihl.cRmeas = Σ(n/n – 1)^1/2^ (Ihl - <Ih>)/Σ(Ihl) where n is the number of observations for unique reflection h with mean intensity <Ih>, summed over all reflections for each observed intensity Ihl.dCC_1/2_ is the correlation coefficient on <I> between random halves of the dataset.Figure 2. Structure of hNotch-1 EGF20–24 reveals an extended conformationX-ray structure of EGF20–24 is shown in a cartoon representation. Ca^2+^-binding EGF domains are shown in green, while the non-Ca^2+^-binding EGF domain is shown in wheat. The Ca^2+^ ions bound in EGF20, EGF21, EGF23, and EGF24 are shown as red spheres. Ser/Thr residues glycosylated in the S2 expression system are labeled; glucose, fucose and xylose are shown in cyan, mid-blue and dark-blue respectively. The unusual solvent exposed position of F930 in EGF24 is shown in green.Table 2. Interdomain tilt and twist angles observed in the X-ray structure of hNotch-1 EGF20-24Domain PairTilt AngleaTwist Anglea20–215.5^o^135.2^o^21–2213.4^o^51.1^o^22–2343.1^o^171.1^o^23–2432.1^o^156.3^o^aThe program mod262^,^63 was used to measure the tilt and twist angles for each pair of EGF domains. The tilt angle informs on the linearity of the domain pair; a value close to 0° is expected for a linear and elongated domain pair while a value close to 90° is expected for a pair oriented at right angles to each other. The twist angle informs on the rotation along the interdomain axis of one domain relative to the other and can be most easily visualized by the relative rotation of the anti-parallel β sheets in the two domains.
As expected, EGF20, EGF21, EGF23, and EGF24 each have a single Ca^2+^ bound and show modification by O-fucose and O-glucose. Ca^2+^ occupancy of the EGF20 site was only 45%; this is likely due to the weak affinity of this non-native N-terminal site (see in the following text).36^,^37^,^38 EGF24 is unusual since the consensus aromatic residue F930, located between the 3^rd^ and 4^th^ cysteine residues and usually involved in stabilizing intramolecular structure, is shifted by one residue (+1) and points out into solvent. It may therefore be available for intermolecular interactions. This feature is specific to mammalian Notch-1 (human, mouse, and rat) and is not seen in Notch-2 (human, mouse, and rat) or Drosophila Notch, all of which have the same conserved 36 EGF domain modular organization. EGF22 does not contain a Ca^2+^-binding consensus sequence. The structure shows a rigid and extended relative orientation of the Ca^2+^-binding EGF21 and non-Ca^2+^-binding EGF22 with a tilt angle of only 13.4^o^. This is in contrast to other Notch Ca^2+^-binding EGF-non-Ca^2+^-binding EGF pairs (∼90^o^ bent structure observed previously for EGF5-6, and the flexible interface observed in solution between EGF9-1030).
Ca2+-binding measurements reveal rigid interfaces for Ca2+-binding EGF domains in the EGF22-25 region of hNotch-1 but a flexible interface at EGF20-21
Ca^2+^ affinities for Ca^2+^-binding (cb) EGF domains of the EGF20-25 region of human Notch-1 were measured to gain insight into the rigidity of interdomain interfaces in this region in solution. In EGF domains, a consensus sequence of D/N/E-x-D/N-D/N/E/Q-x_m_-D/N/Q^∗^-x_n_-Y/F (where ^∗^ indicates possible β-hydroxylation, and m/n are variable) is predictive for Ca^2+^ binding36^,^37^,^38^,^39^,^40 and a high affinity for Ca^2+^-binding is correlated with a well-defined and rigid domain interface.30^,^41 Ca^2+^-binding EGF domains 20, 21, 23, and 25 show the consensus Ca^2+^ binding sequence and the aromatic residue in the preceding domains which contributes to a hydrophobic packing interaction seen in many cbEGF/cbEGF and EGF/cbEGF rigid interfaces.
NMR titrations using ^15^N-labeled EGF20-23 and EGF23-24 were used to measure K_D_ values for the EGF20, EGF21, EGF23, and EGF24 sites (Figure S1).30^,^42 1D ^1^H NMR was used to estimate K_D_ values for EGF25 in the EGF23-25 construct. The measured Ca^2+^ affinities are summarized in Figure 1. The K_D_ values for EGF domains 23, 24, and 25 are in the range of ∼25–∼200 μM and under the conditions of extracellular free Ca^2+^ concentration (∼1.5 mM) and physiological ionic strength (I = 0.15) at pH 7.4 these sites will be saturated to >∼95%. EGF21 has a significantly weaker affinity for Ca^2+^ (K_D_ ∼1.3 mM), despite its native context; this site will only be occupied in ∼50% of molecules. The high Ca^2+^ affinity observed for all the cbEGF domains except EGF21 suggests that the cbEGF domains from 23 to 25 form a packing interaction in solution with the preceding domain leading to a rigid interdomain interface.
Two Ax missense mutations identified in EGF24 (D948V) and EGF25 (N986I) of Drosophila Notch were investigated for their effects on Ca^2+^ binding in EGF pair or triple domain constructs (Figures S2 and S3). Each substitution reduced the Ca^2+^ binding affinity of the mutant domain without impacting the affinity of the neighboring domain. To test the impact of homologous mutations in hNotch-1, D909V was introduced into EGF24 in EGF23-24 and EGF23-25 constructs and reduced the Ca^2+^ binding affinity of the mutant domain but not adjacent domains (Figure S4). Each of these mutations therefore appears to act by introducing flexibility at the 23–24 or 24–25 interfaces.
Dynamics in solution
Heteronuclear NOE shows Ca2+-dependent dynamics in EGF21
The {^1^H}-^15^N heteronuclear NOE provides a method for identifying regions of the polypeptide backbone that undergo fast timescale dynamics (ps/ns).43 Data for the EGF20-23 construct at three different Ca^2+^ concentrations (1.4 mM, 2.8 mM, and 40 mM CaCl_2_) are shown in Figure 3A; under these conditions the occupancy of each Ca^2+^ binding site will differ. Reduced values of the heteronuclear NOE, characteristic of mobile residues, were observed for the first few residues at the N terminus of EGF20 and for several residues between the 3^rd^ and 4^th^ cysteine of EGF20. These residues are involved in Ca^2+^ binding and their mobility reflects the weak affinity of this domain for Ca^2+^ in its non-native context. Interestingly, EGF21, which is placed in a native context, showed similar reduced NOE values for some residues in the region between the 3rd and 4th cysteines at 1.4 mM CaCl_2_, when the domain does not have Ca^2+^ bound (∼23% saturated). The flexibility of these residues decreased in 2.8 mM Ca^2+^ when the site is ∼55% occupied. These residues became rigid in 40 mM Ca^2+^ when the domain is fully bound with Ca^2+^. As expected EGF23 had a rigid backbone, consistent with full occupancy of its Ca^2+^ binding site at each concentration. EGF22, which lacks a Ca^2+^ binding site, also had a rigid backbone under all conditions studied. With a K_D_ of 1.3 mM, the backbone flexibility observed in EGF21 could be functionally relevant at physiological Ca^2+^ concentrations.Figure 3NMR measurements of Ca^2+^-dependent dynamics in EGF20-23(A) {^1^H}-^15^N heteronuclear NOE data for hNotch-1 EGF20-23 measured at Ca^2+^ concentrations of 1.4 mM (black circle), 2.8 mM (red circle) and 40 mM (blue circle) CaCl_2_. Based on experimental K_D_ measurements (Figure 1), the EGF23 site will be fully occupied with Ca^2+^ at all three concentrations. The EGF21 site will be 23%, 55%, and 100% occupied, while the EGF20 site will be 4%, 14%, and 75% occupied. Reduced NOE ratios, characteristic of significant mobility on a nanosecond to picosecond timescale, are observed at the N terminus of EGF20 and for several residues between the 3^rd^ and 4^th^ cysteine of EGF20. EGF21 shows Ca^2+^-dependent NOE values for some residues in the region between the 3^rd^ and 4^th^ cysteines. These residues become rigid in 40 mM Ca^2+^ when the domain is fully bound with Ca^2+^. EGF23, which has Ca^2+^ bound in all three experiments has a rigid backbone. The dashed vertical lines represent the boundary between EGF domains.(B) Residual dipolar couplings (RDCs) measured for 540 μM hNotch-1 EGF20-23 at low (0.84 mM) (black circle) and high (40 mM) (blue circle) Ca^2+^ concentrations. Based on experimental K_D_ measurements (Figure 1), EGF23 will be fully occupied at both concentrations, EGF21 will be 15% and 100% occupied and EGF20 will be 2% and 75% occupied. The magnitude of the RDCs observed in all EGF domains increases as the Ca^2+^ concentration increases from 0.84 mM to 40 mM. This is demonstrated by the large and similar values of 15.4, 16.9, 14.1, and 14.8 obtained for D_a_, the axial component of the alignment tensor, for EGF20, EGF21, EGF22, and EGF23, respectively, at 40 mM CaCl_2_ which indicate an elongated and rigid structure. At 0.84 mM CaCl_2_, D_a_ values of 6.2, 9.0, 9.5, and 9.2 are obtained for EGF20, EGF21, EGF22, and EGF23, respectively. The lower values observed indicate a less elongated structure. The significantly lower value for EGF20 reflects the flexibility that exists between the EGF20 and EGF21 domains when the Ca^2+^-binding site in EGF21 is not fully occupied.
Residual dipolar couplings show Ca2+-dependent interdomain orientations
Residual dipolar couplings (RDCs) are a useful NMR parameter for assessing the relative orientations of protein domains in solution and for identifying interdomain dynamics on a wider range of timescales than the heteronuclear NOE.44^,^45^,^46^,^47^,^48 We have used RDCs previously to define interfaces in the EGF4-EGF13 region of hNotch-1.30 ^1^H^N^-^15^N RDCs were measured for EGF20-23 at two Ca^2+^ concentrations (low-0.84 mM and high-40 mM CaCl_2_) using 2% C12E6/n-hexanol as the alignment medium.49 It can be seen in Figure 3B that the magnitude of the RDCs observed in all four EGF domains increases as the Ca^2+^ concentration changes from low to high. This indicates stronger alignment of EGF20-23 resulting from a more elongated and rigid structure at the higher Ca^2+^ concentration. Fits of the RDCs for EGF22 and EGF23 at both Ca^2+^ concentrations, using the method we described previously,30 yield interdomain tilt and twist angles that are consistent with those observed in the X-ray structure indicating a rigid interface between EGF22 and EGF23 in solution. At the lower Ca^2+^ concentration the RDCs observed for EGF20 are of a smaller magnitude than those observed for EGF21, EGF22, and EGF23. This reflects the flexibility that exists between the EGF20 and EGF21 domains when the Ca^2+^-binding site in EGF21 is not fully occupied. The RDC data are consistent with a Ca^2+^-dependent interface between EGF20 and EGF21, as identified previously using the heteronuclear NOE.
SAXS analysis
We were unable to obtain diffraction-quality crystals of hNotch-1 EGF20-27. To determine the overall shape of this region and to assess possible flexibility in solution, SAXS measurements were collected on a sample resolved by SEC in 5mM Tris, pH 7.5, 15 mM Ca^2+^ (Table 3). The shape of the scattering curve (Figure 4A) shows that EGF20-27 is non-globular, as expected for this multi-domain construct, while the Kratky plot (Figure 4B) demonstrates that EGF20-27 has an extended rather than an unfolded structure. The P(r) distribution (Figure 4C) is also consistent with an extended structure for this 8-domain construct and shows a D_max_ of 238 Å for EGF20-27; a completely extended model for an 8-domain construct would be expected to have a D_max_ of ∼240 Å based on a length of ∼30 Å per EGF domain. A model consisting of the EGF20-24 X-ray structure and single EGF domain structures for EGFs 25, 26 and 27 was refined against SAXS data for EGF20-27 using SREFLEX from the ATSAS suite50 and shows an extended and near linear conformation (Figure 4D). A similar SAXS analysis was carried out for a smaller EGF23-27 construct and is also consistent with an extended structure for EGF23-27 with a shorter D_max_ of 160 Å determined from the P(r) distribution (Figure S5). In addition, we have used AlphaFold2 (see STAR Methods) to generate a model for EGF20-27. This shows an extended structure that is similar to that generated by SREFLEX but the latter model gives better agreement with experimental SAXS data (Figure S6).Table 3SAXS data collection and processing parameters for hNotch-1 EGF20-27Data collection parametersInstrumentDiamond Light SourceBeamlineB21Wavelength (Å)0.9537q-range (Å ^−1^)0.0032–0.38Sample-to-detector distance (m)3.6ExperimentSEC-SAXSExposure time per frame (sec)1Temperature (°C)20DetectorEiger 4MStructural parametersI_0_ (cm^−1^) [from Guinier]0.043Rg (Å) [from Guinier]49.61 ± 0.25sRg limits (nm)0.62–1.3Dmax (Å) [Gnome]238I_0_ (cm^−1^) [from p(r)]0.04Rg (Å) [from p(r)]56.10Bayesian Molecular mass (kDa) [Primus]53.1Calculated Mw from sequence (kDa)32.4Figure 4SAXS data for hNotch-1 EGF20-27 reveal an extended shape(A) A scaled, merged, and averaged X-ray scattering curve was collected with purified hNotch-1 EGF20-27 at 2.5 mg/mL in 5mM Tris, pH 7.5, 15 mM CaCl_2_.(B and C) (B) Kratky plot and (C) P(r) distribution derived from the scatter curves collected from purified hNotch-1 EGF20-27.(D) Model for EGF20-27 derived from the SAXS data using SREFLEX refinement (also see Figures S5 and S6). Ca^2+^-binding and non-Ca^2+^-binding domains are indicated in green and wheat respectively.
Ligand-dependent activation of Notch is reduced by substitutions in the EGF24-26 region
The functional importance of the EGF20-27 region was investigated by comparing the JAG1 and DLL4-dependent activity of full-length Notch-1 variants using a well-established luciferase complementation assay.51 These included variants with single Ca^2+^ binding consensus substitutions D909V in EGF24, N947I in EGF25 (homologous to Drosophila Ax suppressor variants) and a DVNI double variant, C933S (EGF24) and C960Y (EGF25) (homologous to Drosophila Ax homozygous lethal variants) and S990N (EGF26) a substitution commonly found in aging esophageal tissue (Figure 5A).4 An EGF12 L468A substitution, which has previously been shown to be critical for ligand binding,52 was also tested and acted as a negative control. Initially both an EGTA-induced ligand-independent activation assay, and direct measurement of cell surface Notch levels were used to demonstrate similar expression levels of mutant receptor on the cell surface compared to WT for D909V, N947I, and DVNI, and the control L468A variant (Figure S7). However C933S, C960Y, and S990N all showed reduced cell surface levels. In ligand-dependent assays, either utilizing cells expressing full-length mouse ligands (Jagged1 or Dll-4) or plate-bound purified human ligand fragments (JAG1 or DLL4), variants D909V, N947I, and DVNI all reduced the ability of Notch-1 to be trans-activated in response to both ligands, (Figure 5B; Figure S7). This reduction in activation was not as pronounced as that observed for L468A (Figure 5B) but nonetheless indicated an important role for the Ca^2+^ dependent structure of this region. As expected C933S and C960Y both showed reduced activity consistent with lower expression levels on the cell surface, most likely as a consequence of misfolding due to loss of a key disulphide bond. Thus the homozygous lethality associated with a subset of Drosophila Ax mutations, may be explained by a substantial quantitative defect of Notch at the cell surface. Variant S990N also showed reduced activity consistent with low cell surface levels. This may be due to defective O-glucosylation since modification of Ser/Thr residues between C1 and C2 within Notch EGF domains is known to facilitate folding (although S990, located at +3 from C1 within this loop region does not conform to the defined consensus).53 However the asparagine substitution at residue 990 creates an NxC motif which can be associated with N-glycosylation.54 Thus it is also possible that incorrect addition of an N-glycan to EGF26 impedes folding. The quantitative defect associated with this substitution further supports the hypothesis that loss of function Notch mutations accumulate in aging esophageal tissue.4Figure 5. Luciferase reporter assays demonstrate Notch-1 variants from EGF24-26 region show defects in trans-activation and cis-inhibition(A) (Upper panel) Amino acid substitutions from the Notch-1 EGF24-26 region studied. Conserved Cys residues indicated in yellow, Ca^2+^-binding consensus residues shown in red, residues which form interdomain packing interactions shown in blue and green. (Lower panel) Cartoon of trans-activation and cis-inhibition cellular assays using the Notch luciferase reporter is shown. Ligand in red, Notch in blue. Cis-inhibition reduces the observed luminescence.(B) Ligand-dependent trans-activation assays were performed with Notch-1 variant lines co-cultured with B16 cells expressing mJag1 (Jagged1) or mDll4 (Dll-4). Graphs show the combined results of four independent plates, each with five repeat wells. Signal in each case is shown relative to the WT Notch-1 (mN1 wt) line co-cultured with ligand. Data presented as mean ± SD. Statistical significance was determined by Kruskal-Wallis test and Dunn’s post hoc test (^∗∗^ = p < 0.01, ^∗∗∗^ = p < 0.001, ^∗∗∗∗^ = p < 0.0001).(C) Cis-inhibition of Notch-1 variant cells transfected with pcDNA3.1/full-length WT hJAG1. Controls were empty pcDNA3.1 or pcDNA3.1/hJAG1F207A/E228K (Notch-binding defective). Signal shown relative to WT Notch-1 (mN1 wt) transfected with empty pcDNA3.1 and co-cultured with B16 cells expressing mJag1. Cis-inhibition (black) is monitored indirectly as a decrease in luminescence signal arising from cells transfected with cis-JAG1 and shown alongside the luminescent signal obtained from cells transfected with empty vector (hatched). Data presented as mean ± SD.(D) Percentage reduction in Notch trans-activation observed for cis-inhibited Ax variants (1-(% luminescence with cis-hJAG1)/(% luminescence empty vector)) ×100 based on data in (C). Data presented as mean ± SD. Statistical significance was determined by ordinary one-way ANOVA and Dunnett’s post hoc test (^∗∗∗^ = p < 0.001, ^∗∗∗∗^ = p < 0.0001) (also see Figures S7 and S8).
Cis-inhibition of Notch-1 requires native Ca2+-dependent structure of EGF24-25
D909V, N947I, and DVNI variants were also tested in a cis-inhibition assay where each Notch variant cell line was transiently transfected with a construct expressing full-length JAG1 on the same membrane and challenged with cell surface-expressed ligands on B16 cells (Figures 5A, 5C, and S7). Lower levels of Notch activation were seen when WT JAG1 was expressed in the same cell as WT Notch-1, indicative of cis-inhibition. This was a direct effect of ligand ectodomain on Notch ligand-binding region (LBR) since the lower receptor activity observed was reversed by introduction of an F207A/E228K DSL JAG1 variant which disrupts the ligand/receptor interface (Figures 5C and S7).17^,^25 The Notch variants, when similarly tested, also showed evidence of cis-inhibition, since the activity level of each cell line was reduced compared to that expressing empty plasmid alone. However, overall activity of variant and WT lines was very similar, once cis-inhibited (Figure 5C). Similar data were obtained irrespective of the ligand used to trans-activate the cis-inhibited cells (Figure S7). These data suggest there is also a negative effect of the Ca^2+^-binding substitutions on the ability of Notch to be cis-inhibited (Figure 5D); otherwise the overall activity levels seen would be lower (reflecting the defect in trans-activation associated with these variants) (Figure 5B).
Discussion
Although the Notch receptor was identified more than 30 years ago, the architecture of the large extracellular domain, which is crucial for understanding how Notch can form both activatory and inhibitory complexes with its ligands Delta and Serrate/Jagged, is still unknown. Up to now, high-resolution structural methods have failed to provide models for the full extracellular domain due to the difficulties of studying such a large extended and flexible molecule. Furthermore, electron microscopy (EM) studies have been largely unsuccessful at identifying receptor architecture due to problems associated with obtaining pure, natively folded protein (108 disulphide bonds in the EGF-domain-rich sections of Notch-1 alone) and the very narrow dimensions (in two directions) of this fibrous protein. Despite these difficulties, progress has been made by applying structural and biophysical methods to analyze multidomain fragments from different regions and combining these with functional studies. As a result, a structural model for hNotch-1 EGF4-13 has been produced which demonstrates that this region is not simply a rigid straight rod but has flexibility and non-linear sections.30 Using similar methods, combined with SAXS analysis, we focused on a region C-terminal to the LBR which in Drosophila gives rise to distinctive Abruptex phenotypes when mutated, suggesting an important functional role in regulating Notch activity.32
Using X-ray crystallography, NMR, and SAXS analyses we describe a Ca^2+^-dependent elongated structure for hNotch-1 EGF20-27 which is rigid in solution at high Ca^2+^ concentrations. Under conditions which mimic extracellular free [Ca^2+^], we observe one weak affinity Ca^2+^ binding site in EGF21 which could confer flexibility at the EGF20/EGF21 interface. A structural model based on these data, together with that previously shown for EGF4-13, is shown in Figure 6. Full-length Notch variants with Ca^2+^-binding substitutions in EGF24 and EGF25 show a reduced ability to be trans-activated and cis-inhibited in cell-based assays, demonstrating the importance of rigidity of EGF23/24 and EGF24/25 domain interfaces for these functions. Notch activity, when ligand is present in cis or trans, is dependent on the Notch LBR interacting with ligand DSL, as evidenced by the effects of ligand DSL substitutions F207A/E228K (Figures 5C, S7, and S8) which abrogate the known receptor/ligand interface. These data suggest that the DSL-EGF11 antiparallel interaction identified in high resolution structures of receptor/ligand complexes is common to both cis-inhibitory and trans-activatory complexes.Figure 6. Structural model of the ectodomain of hNotch-1 comprising EGF domainsThe previously published model of the EGF4-13 region is shown alongside that of the EGF20-27 region derived from the X-ray structure of EGF20-24 and SAXS data for EGF20-27. The flexible region determined by {^1^H}-^15^N heteronuclear NOE and RDC data is indicated with a blue arrow. The thick horizontal lines show the EGF4-13 region (containing the ligand-binding region (LBR) and the flexible interface between EGF9 and EGF10), the EGF20-24 region (determined by X-ray crystallography), and the EGF20-27 region (containing the Abruptex (Ax) region studied here and modeled by SAXS). Ca^2+^ and non-Ca^2+^-EGF domains are indicated in green and wheat respectively.
Significance of Ca2+ binding
A variety of methods has been used previously to identify the Ca^2+^-binding affinity of a distinctive subset of EGF domains ranging from mM to nM dependent upon the domain context.41 This measurement is of biological significance since the extracellular environment of EGF-rich proteins is high in Ca^2+^ and saturation of such sites, in EGF-cbEGF or cbEGF-cbEGF pairs usually confers rigidity to interfaces. This can facilitate Ca^2+^-dependent protein-protein interactions (particularly where the interaction surface extends over a number of different adjacent domains), protect against proteolysis or confer biomechanical properties/resistance to pulling of the molecule in tissues subjected to tension.
In hNotch-1 at least 20 of the 36 domains are of the Ca^2+^-binding type with a clearly defined set of residues known either to ligate Ca^2+^ directly or contribute to the stability of the Ca^2+^-binding site. So far most sites studied in Notch-1 have exhibited affinity such that they would be saturated under physiological conditions (free Ca^2+^ ∼1.5 mM I =0.15, pH 7.4). However, unusually a weak affinity site was observed in EGF21 which imparted dynamic behavior on the EGF20-21 interface. The other sites determined here at 23, 24, and 25 (see Figure 1) would all be expected to be saturated under physiological conditions. In fibrillin-1, another extensively studied cbEGF-rich protein, only one similar weak site in the mM range has been identified (cbEGF32). Its functional significance is underscored by the discovery of a Marfan syndrome mutation resulting in a cbEGF32 N to S substitution, which by altering the geometry of Ca^2+^ ligation weakens the affinity 9-fold from ∼1.6 mM to 14 mM.55 Thus, the Ca^2+^-binding affinity of EGF domains within a protein is fine-tuned for biological function. The flexibility at the hNotch-1 EGF20-21 domain interface conferred by a weak site may facilitate trafficking, protein-protein interactions in different contexts, contribute to catch bond behavior, or impart biomechanical properties. One other site of flexibility has previously been observed in hNotch-1 between EGF9 and 10.30 In this case, EGF10 is non-Ca^2+^ binding and therefore the properties of this interface are Ca^2+^ independent. These sites of flexibility may explain the relatively short measurement for the Notch ectodomain calculated by SAXS in a previous study.31
Cis-trans interactions
It has been known for many years that cis and trans complexes of Notch with its ligands can form resulting in inhibitory/activatory effects on signaling. The mechanism by which such complexes could form when receptor and ligand were both membrane-bound, had asymmetrical binding sites and were apparently composed of rigid domain interfaces was difficult to envisage. Identification in this study of a Ca^2+^-dependent pairwise linkage within contiguous EGF repeats, membrane proximal to the LBR, together with earlier identification of a flexible EGF9-10 interface, indicates that a variety of conformations are possible at the cell surface which might facilitate cis and trans interactions with ligand. For example, rotation about the EGF20-21 interface under physiological conditions could allow Notch to dock with ligand presented in cis (on the same cell surface) or in trans (on the opposing cell surface) while maintaining the same core ligand-binding interface. Flexible regions in receptors involved in cis and trans interactions with ligands which maintain the same binding site, such as Ly49 and MHC-I56 and LILRB and MHC-I,57 have previously been proposed. However other distinct modes of binding, such as seen in plexin receptors with semaphorins, can also occur.58 The ability of hNotch-1 to sample a range of conformations may also allow it to dimerize on the same cell surface, form intramolecular interactions within the same monomer, and interact with ligands in cis and in trans.
Functional experiments
Functional data indicate that the Ca^2+^-dependent conformation of the EGF24/EGF25 region which would be extended and rigid under physiological conditions is important for Notch-1 activation. Impairment of Ca^2+^ binding to these two domains does not impact on trafficking however as variants show similar levels of ligand-independent activation and cell surface expression (Figure S7). The location of this region between the LBR ligand interaction site and the NRR containing the S2 site cleaved by ADAM10 protease may help facilitate ligand binding in cis and trans but may also aid transmission of the pulling force required for S2 cleavage of the NRR mechanosensor. The Ax region has been observed previously to interact with the LBR of Notch suggesting that it may compete with ligand for this region.59^,^60
Dominant Ax missense mutations which affect the EG24-29 region in Drosophila Notch result in a shared phenotype, but can be distinguished by their behavior when crossed with Notch null alleles. The “suppressor” class of missense mutations affect EGF24 and EGF25 (and cause defective Ca^2+^ binding, Figure S3) while the “enhancer” class affect residues in C-terminal portions of EGF27 and 29, both of which are non-Ca^2+^-binding domains. Our study indicates that loss of Ca^2+^-stabilized interfaces at EGF23-24 and EGF24-25 underlies the suppressor phenotype and reduces the ability of Notch to be trans-activated and cis-inhibited. These cellular data suggest that the phenotypic consequences of such mutations may ultimately depend on the balance of cis- and trans- interactions that occur in a tissue context. When trans signals dominate, Notch activation may be reduced in the suppressor variants compared to WT. Where cis-inhibition occurs, activation may be similar to WT receptor. This may explain the observed loss of repression of Notch activity of Ax variants at the dorsal/ventral border in the Drosophila wing disc, compared to WT, when ligands Delta or Serrate are expressed ectopically.61
Overall, this study has increased our understanding of the architecture of the Notch ectodomain by providing information on the conformation of the EGF20-27 region. These data, coupled with earlier high- and low-resolution studies, show that the extracellular portion does have extended sections, but has at least two points of flexibility within it which allow the receptor to sample a range of conformations. Cellular experiments have demonstrated the importance of the Ca^2+^-dependent rod-like structure of the EGF24-25 region for optimal trans-activation and cis-inhibition and thus establishes a mechanism for the effects of unusual phenotypes associated with Abruptex mutations.
Resource availability
Lead contact
Requests for further information and further resources should be directed to, and will be fulfilled by the lead contact, Penny A Handford ([email protected]).
Materials availability
Reagents generated in this study will be made available on request, but we may require a payment and/or a completed materials transfer agreement if there is potential for commercial application.
Data and code availability
- •The coordinates of EGF21-23 and EGF20-24 have been deposited in the Protein Data Bank under accession numbers PDB: 9B3G and PDB: 9B3N.
- •Resonance assignments for EGF20-23 have been deposited in the BioMagResBank (BMRB) under accession number BMRB: 51699. They are publicly available as of the date of publication. Accession numbers are also listed in the key resources table.
- •All data reported in this paper will be shared by the lead contact on request.
- •This paper does not report original code.
- •Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Acknowledgments
This work was supported by 10.13039/501100000265MRC grant MR/R009317/1 and MR/V008935/1 awarded to P.A.H. S.M.L. was supported by a 10.13039/100010269Wellcome Trust Investigator Award. J.H., T.R., and P.W. were supported by 10.13039/100010269Wellcome Trust, 10.13039/501100000268BBSRC and 10.13039/501100000289Cancer Research UK studentships, respectively. We thank Abi Boyce and Lucy Barber for technical help. We thank the Diamond Light Source for access to SAXS and X-ray crystallography beamlines. HEK-RBP cells were a kind gift from R.Kopan, University of Cincinnati College of Medicine.
Author contributions
Protein constructs were expressed and purified by H.N., Z.B., P.C.W., S.L., R.J.S., J.H., and T.R.; T.R., J.H., P.C.W., S.L., and C.R. collected and analyzed Ca^2+^-binding data. J.H. and C.R. collected and analyzed heteronuclear NOE and RDC data. T.R., H.N., Z.B., and E.D.L. collected and analyzed SAXS data. R.J.S., D.S., S.J. and S.M.L. collected and analyzed crystallographic data. Z.B., T.R., B.K., Y.M., and P.A.H. collected and analyzed Notch activation data. P.A.H., C.R., and S.M.L. conceived and supervised the research and wrote the manuscript. All authors discussed the results and implications of the data.
Declaration of interests
The authors declare no competing interests.
STAR★Methods
Key resources table
REAGENT or RESOURCESOURCEIDENTIFIERAntibodiesRat monoclonal anti-mouse Notch-1 (22E5), APCInvitrogenCat#17-5765-82; RRID:AB_10670632Bacterial and virus strainsBL21 pREP4This paperN/AChemicals, peptides, and recombinant proteinsHuman Notch-1 EGF21-23This paperN/AHuman Notch-1 EGF20-23This paperN/AHuman Notch-1 EGF20-24This paperN/AHuman Notch-1 EGF23-24This paperN/AHuman Notch-1 EGF23-24 D909VThis paperN/AHuman Notch-1 EGF23-27This paperN/AHuman Notch-1 EGF23-25This paperN/AHuman Notch-1 EGF20-27This paperN/ADrosophila Notch EGF23-24This paperN/ADrosophila Notch EGF23-25This paperN/ADrosophila Notch EGF23-24 D948VThis paperN/ADrosophila Notch EGF23-25 D948VThis paperN/ADrosophila Notch EGF23-25 N986IThis paperN/A^15^N ammonium chlorideGoss ScientificNLM-467; Labeled CAS: 2483734-97-15; Unlabeled CAS: 6521-29-12^13^C GlucoseGoss ScientificCLM-1396-5; CAS: 110187-42-3D_2_OSigma Aldrich151882-125G; CAS: 7789-20-0Deposited dataBMRB NMR assignmentsThis paperBMRB: 51699Human Notch-1 EGF20-24 structureThis paperPDB: 9B3NHuman Notch-1 EGF21-23 structureThis paperPDB: 9B3GExperimental models: Cell linesS2Expres2ion Biotechnologies, DenmarkCat#94-005FHEK293/RBPJkIlagan et al.51N/AHEK293/RBPJk/mN1 WTIlagan et al.51N/AHEK293/RBPJk/mN1 D909VThis paperN/AHEK293/RBPJk/mN1 N947IThis paperN/AHEK293/RBPJk/mN1 D909V/N947IThis paperN/AHEK293/RBPJk/mN1 C993SThis paperN/AHEK293/RBPJk/mN1 C960YThis paperN/AHEK293/RBPJk/mN1 S990NThis paperN/AHEK293/RBPJk/mN1 L468AThis paperN/AB16 WTMasiero et al.80N/AB16 mJag1Masiero et al.80N/AB16 mDll4Laboratory of Adrian L. HarrisN/AOligonucleotidesPrimers used in this paper, see Table S1.This paperN/ARecombinant DNAHuman Notch-1 EGF21-23 in pQE30This paperN/AHuman Notch-1 EGF20-23 in pQE30This paperN/AHuman Notch-1 EGF23-24 in pQE30This paperN/AHuman Notch-1 EGF23-24 D909V in pQE30This paperN/AHuman Notch-1 EGF23-27 in pQE30This paperN/AHuman Notch-1 EGF20-24 in pEXS2.2This paperN/AHuman Notch-1 EGF20-24 in pEXS2.2This paperN/AHuman Notch-1 EGF20-27 in pEXS2.2This paperN/ApcDNA5/FRT mN1 L468AThis paperN/ApcDNA5/FRT mN1 D909VThis paperN/ApcDNA5/FRT mN1 N947IThis paperN/ApcDNA5/FRT mN1 D909V/N947IThis paperN/ApcDNA5/FRT mN1 C933SThis paperN/ApcDNA5/FRT mN1 C960YThis paperN/ApcDNA5/FRT mN1 S990NThis paperN/ADrosophila Notch EGF23-24 in pQE30This paperN/ADrosophila Notch EGF23-25 in pQE30This paperN/ADrosophila Notch EGF23-24 D948V in pQE30This paperN/ADrosophila Notch EGF23-25 D948V in pQE30This paperN/ADrosophila Notch EGF23-25 N986I in pQE30This paperN/ASoftware and algorithmsPhaserMcCoy et al.69https://phenix-online.org/Xia2Winter et al.68https://phenix-online.org/PHENIXLiebschner et al.66https://phenix-online.org/COOTEmsley et al.67https://www2.mrc-lmb.cam.ac.uk/personal/pemsley/coot/AlphaFold2Jumper et al.70https://github.com/google-deepmind/alphafoldCCPN softwareVranken et al.72https://ccpn.ac.uk/NMRPipeDelaglio et al.71https://www.ibbr.umd.edu/nmrpipe/ATSAS package (DAMMIF, DAMAVER, CRYSOL)Manalastas-Cantos et al.50https://biosaxs.com/software.htmlGraphPad Prism 9GraphPad, San Diego, CA, USAwww.graphpad.com
Experimental model and study participant details
Strains used in protein production
The hNotch-1 EGF20-24 and hNotch-1 EGF20-27 protein fragments used in this study were expressed in S2 cells.18 The hNotch-1 EGF20-23, EGF21-23, EGF23-24, EGF23-25, EGF23-27 protein fragments used in this study were expressed in Escherichia coli BL21 cells transformed with a pQE-30 (Qiagen) expression vector and pREP4 plasmid for control of expression via the lac repressor.30 hNotch-1 EGF23-27 contained serine substitutions at two non-canonical cysteine residues in EGF25 (Cys963Ser) and EGF27 (Cys1040Ser) to facilitate in vitro refolding. Drosophila Notch fragments EGF23-25, EGF23-25 D948V, EGF23-25 N986I and EGF23-24 were produced using the same system. When cloned into the expression vector, an N-terminal His_6_ tag was included for purification, followed by an Ser-Ala spacer and either a factor Xa protease recognition site (Ile-Glu-Gly-Arg) (constructs d/hEGF23–24, d/hEGF23-25, hEGF 23–27) or an enterokinase cleavage site (constructs EGF20-23, EGF21-23) for later removal of the His_6_ tag. Ligand constructs were produced as C-terminal Fc and His-tag fusion proteins by transiently transfecting HEK293T cells with cDNA cloned into pHLSec plasmid (hJAG1 NE3)17 or HEK293F cells with cDNA cloned into pcDNA3.1 (hDLL4 NE3).
Method details
Protein expression, purification, refolding and characterization
Protein expression, isotopic labeling, refolding and purification protocols for preparation of NMR samples have been described previously.64^,^65 Briefly for proteins expressed in E.coli, cell lysates were spun down at 40,000 rpm for 45 min (Beckman L7–55) and supernatant loaded onto a Ni^2+^ chelating Sepharose column (GE Healthcare). Following elution of His-tagged protein with buffer containing 50 mM EDTA and 100 mM Tris pH 8.3, protein was reduced for 1 h at RT by addition of dithiothreitol (DTT) to a final concentration of 0.1 M. The pH was adjusted to pH ∼2, by addition of concentrated HCl, and the solution was dialyzed against 0.1% (v/v) trifluoroacetic acid (TFA) overnight. Following filtration (0.2 μM filter (Millex-GP)), the soluble fraction was purified by reverse-phase HPLC (C8 column) using a Beckman Gold system. Purified, reduced protein was subsequently refolded in an oxido-shuffling buffer containing 100 mM Tris–HCl pH 8.3, 3 mM L-cysteine, 0.3 mM L-cystine, 0.2 mg/mL protein at 37°C for ∼48 h. Following acidification to pH ∼2, dialysis was performed against 0.1% (v/v) TFA for >5 h. Protein was subsequently concentrated by ultrafiltration and purified by HPLC. After lyophilization, the His_6_ tag was removed by incubation overnight with either one unit of bovine factor Xa (DENZYME) per mg of protein in 50 mM Tris–HCl pH 7.5, 0.1 M NaCl, 1 mM CaCl_2_ at 37°C (or enterokinase (NEB) in same buffer) were incubated with an enzyme:protein ratio by weight of 1:1,000,000 overnight at RT. Protein was further purified by cation-exchange fast liquid protein chromatography and HPLC. hNotch-1 receptor constructs EGF20-24 and EGF20-27 were recombinantly expressed in S2 insect cells (Expres2ion Biotechnologies, Denmark) as C-terminal His-tagged fusion proteins.18 Medium containing recombinantly expressed protein was filtered and loaded onto a cOmplete His-tag Purification Column (Roche Diagnostics, UK), for purification. Following washing with 50 mM Tris pH 9.0, 2.5 mM imidazole, 200 mM NaCl and 5 mM CaCl_2_, proteins were eluted with wash buffer containing 250 mM imidazole. Proteins were further purified by size-exclusion chromatography (SEC) using a Superdex S200 (GE Healthcare, Sweden) preparative column in 10 mM Tris pH 7.5, 200 mM NaCl and 5 mM CaCl_2_ buffer. Each nanomole of protein was incubated with 0.5 units of N-terminal His-tagged HRV-3C protease (Sigma) overnight at 4°C for His tag removal. Non-cleaved protein and HRV-3C protease were removed through incubation with Ni-NTA agarose (Qiagen) for 1 h at 4°C. The supernatant was collected and purified by SEC. Fc and His-tagged ligand proteins for activation assays were purified as previously described.17 Cell medium containing hJAG1 NE3 protein was mixed with 3 times volumes of 50 mM Tris and 200 mM NaCl (pH 9.0). Cell medium containing hDLL4 NE3 was mixed with 2 times volumes of 20 mM MES and 200 mM NaCl (pH 6.5). The diluted medium was filtered and loaded onto a cOmplete His-tag Purification Column (Roche Diagnostics, UK) for purification. Following washing with 50 mM Tris, 10 mM imidazole and 200 mM NaCl (pH 9.0) for hJAG1 NE3 or 20 mM MES, 50 mM imidazole and 200 mM NaCl (pH 6.5) for hDLL4 NE3, proteins were eluted with wash buffer containing 300 mM imidazole. Proteins were further purified by SEC using a SuperdexTM S200 (GE Healthcare, Sweden) preparative column in 50 mM Tris and 200 mM NaCl (pH 7.5).
X-Ray crystallography
Human Notch-1 EGF21-23 was crystallized by vapor diffusion from sitting drops at 9 mg/mL, with 100 nL mother liquor (0.2 mM ammonium acetate, 0.1 M BIS-Tris pH 5.5, 25% (w/v) PEG3350) and 100 nL protein solution (25 mM Tris pH 7.5, 150 mM NaCl, 10 mM BaCl_2_). Crystals were cryo-protected by addition of 30% (v/v) ethylene glycol, and data collected at Diamond Light Source on beamline I04-1. The dataset was indexed, integrated and scaled using Xia2 (Winter, 2010). The structure was phased and built iteratively using AutoSol in PHENIX (dev-4694),66 using the anomalous signal from Ba^2+^. The structure was refined using PHENIX.refine (dev-4694),66 with COOT used for manual rebuilding and inspection.67
Human Notch-1 EGF20-24 was crystallized by vapor diffusion from sitting drops at 20.7 mg/mL, with 200 nL mother liquor (0.2 mM imidazole malate pH 6, 8% (w/v) PEG4K) and 200 nL protein solution (5 mM Tris pH 7.5, 50 mM NaCl, 10 mM CaCl_2_). Crystals were cryo-protected by addition of 30% (v/v) ethylene glycol and 40 mM CaCl_2_, and data collected at Diamond Light Source on beamline I03. The dataset was indexed, integrated and scaled using Xia2.68 The structure was phased using molecular replacement in Phaser,69 using the hN-1 EGF21-23 and AlphaFold270 models for EGFs 20 and 24. The structure was refined using PHENIX.refine (dev-4694),66 with COOT used for manual rebuilding and inspection.67
NMR spectroscopy
All NMR experiments were carried out using spectrometers operating at ^1^H frequencies of 500, 600 and 750 MHz equipped with Bruker Avance II (500 MHz) or Avance IIIHD (600 and 750 MHz) consoles and 5mm TCI cryoprobes. Data were processed using NMRPipe71 and spectra were analyzed using the CCPN software.72
Resonance assignments for EGF20-23 were carried out using ^13^C/^15^N labeled protein and standard methods as described previously.64^,^73 Unless otherwise stated, all NMR experiments were carried out at 25°C in 5 mM Tris-HCl, 150 mM NaCl at pH 7.5 in 95% H_2_O/5% D_2_O.
Ca^2+^ dissociation constants for the hNotch-1 EGF20-23 and EGF23-24 constructs were obtained from Ca^2+^ titrations monitored by 2D ^1^H-^15^N HSQC experiments, collected at 500 MHz. Values for EGF23-25, Drosophila Notch dN EGF23-24 and dN EGF23–25 were obtained from 1D and 2D ^1^H NMR experiments.41^,^42^,^74 Samples were initially Ca^2+^ free and the Ca^2+^ concentration was increased by addition of CaCl_2_ aliquots up to saturating concentrations (usually ∼25 mM).
{^1^H}-^15^N heteronuclear NOE experiments were carried out on ^15^N-labeled EGF20-23 in order to examine the sub-nanosecond dynamics of specific amides.75 Samples for measurement of the heteronuclear NOE contained 800μM EGF20-23 and 1.4 mM, 2.8 mM or 40mM CaCl_2_. The samples with 1.4 mM and 2.8 mM CaCl_2_ contained 150 mM NaCl while the 40 mM CaCl_2_ did not. With 1.4 mM CaCl_2_, the EGF23 site will be fully occupied with Ca^2+^ while EGF21 and EGF20 will be ∼23% and ∼4% occupied, respectively. With 2.8 mM CaCl_2_, the EGF23 site will be fully occupied with Ca^2+^ while EGF21 and EGF20 will be ∼55% and ∼14% occupied, respectively. Spectra with and without ^1^H saturation were collected as interleaved experiments; the {^1^H}-^15^N NOE was calculated as the ratio of the peak intensities in the spectra recorded with and without ^1^H saturation. Data were collected at a ^1^H frequency of 750 MHz. ^1^H saturation was applied for 4 s.
Residual dipolar couplings (RDCs) were collected for the EGF20-23 using liquid crystalline media containing n-alkyl-poly(ethylene glycols) (PEG) and n-alkyl alcohols as described previously.49 The final concentration of C12E6/hexanol used was 2%. EGF20-23 samples for measurement of the RDCs contained 540 μM EGF20-23 and 0.84 mM or 40 mM CaCl_2_. The sample with 0.84 mM CaCl_2_ contained 150 mM NaCl while the 40 mM CaCl_2_ sample did not. With 0.84 mM CaCl_2_, the EGF23 site will be fully occupied with Ca^2+^ while EGF21 and EGF20 will be ∼15% and ∼2% occupied, respectively. With 40 mM CaCl_2_, the EGF 21 and EGF23 sites will be fully occupied with Ca^2+^ while EGF20 will be ∼75% occupied. Isotropic spectra and aligned spectra were collected for protein solutions in 90% H_2_O/10% D_2_O using the interleaved IPAP experiment76 performed at a ^1^H frequency of 500 MHz. Residual dipolar couplings were measured as the difference between the splitting observed in the isotropic and aligned datasets. RDC values for EGF20, EGF21, EGF22 and EGF23 were fitted to the X-ray coordinates of EGF20-24, using an in-house program. The overall fit between experimental and calculated RDC values was assessed using the quality factor (Q) defined as: Q = [∑i=1, …,N (RDC^expt^ – RDC^calc^)^2^/N]^½^/RDC_rms_.77 With 0.84 mM CaCl_2_, Q values of 0.24, 0.23, 0.23 and 0.21 were obtained for EGF20, EGF21, EGF22 and EGF23. With 40 mM CaCl_2_, Q values of 0.23, 0.22, 0.24 and 0.21 were obtained for EGF20, EGF21, EGF22 and EGF23. These Q values indicate a good fit of the experimental RDCs to the X-ray structure.
SAXS
Data collection on hNotch-1 EGF23-27 and EGF20-27 samples was carried out at the Diamond Light Source at Beamline B21 using a SEC-SAXS method. 100 μL samples were prepared (100 μL, 2.5 mg/mL, 5 mM Tris pH7.5, 15 mM CaCl_2_), including dialysis in 2 L of buffer overnight at 4°C (Table 3). Samples were subsequently frozen at −80°C for transport to the beamline and thawed immediately prior to use. SAXS samples (50 μL) were autoloaded onto a Shodex 402.5 analytical SEC column. The eluent from this column then passed through the X-ray beam and scatter pattern recorded. Data were collected up to a scattering vector (s) of 0.37 Å^−1^. Scatter resulting from buffer was subtracted from the peak corresponding to protein scatter using the program SCATTER 3.078 and the ATSAS software package50 was used for further analysis. Guinier analysis and ab initio modeling and structure fitting was carried out using the program SREFLEX in the ATSAS software package.50 Modeling was performed at default settings using 10 repetitions followed by DAMAVER. CRYSOL was subsequently used to confirm the data fits the structure model. A model for hNotch-1 EGF20-27 was also generated using AlphaFold2.70
HEK-RBP cells growth and transfection
mN1 Ax variants were created by overlap extension PCRs41 using a cDNA fragment from 521 bp to 4131 bp of the full-length mN1. The mutated cDNA fragment was initially inserted between the SbfI and BamHI sites of plasmid pUC19 and following sequencing and amplification, was digested and ligated into pcDNA5/FRT plasmid containing the remainder of the mN1 cDNA.
HEK-RBP cells were maintained in DMEM medium, supplemented with 10% FCS (Gibco), 2 mM L-glutamine (Gibco), 50 U/mL penicillin, 0.05 mg/mL streptomycin (Sigma) and 0.2 μg/mL puromycin. Cells were transfected using Mirus TransIT-293 Transfection Reagent and carried out using 80% confluent wells in a 6-well plate, using a total of 1000 ng of DNA per transfection (pOG44: pcDNA5/FRT mN1 at a ratio of 9:1). 24 h following transfection, cells were transferred to 15 cm dishes and additional selection (100 μg/mL hygromycin) applied. Clones were subsequently transferred to 6-well plates and expanded in the presence of both 0.2 μg/mL puromycin +100 μg/mL hygromycin.
B16 cells growth conditions
B16 cells expressing either mDll4 or mJag1 were maintained in RPMI medium, supplemented with 10% FCS (Gibco), 50 U/mL penicillin and 0.05 mg/mL streptomycin (Sigma) and split 1:15 once confluent.79^,^80
Notch activation assays
Ligand-independent assay
Established HEK-RBP cell lines expressing Notch-1 variants were split 1:3 into DMEM +10% FCS. After two days, 40,000 cells were added to each well of a 96-well Costar black clear bottom plate (DMEM +10% FCS). 48 h later, wells were washed with HBS (20 mM HEPES pH 7.4, 150 mM NaCl, 0.9 mM Ca^2+^), and cells resuspended in 100 μL HBS +150 μg/mL D-luciferin +10 mM EGTA. Control wells were treated with substrate solution without EGTA. Luciferase activity was monitored using a BMG LABTECH PHERAstar FS plate reader at 40 min after addition of EGTA.
Ligand-dependent assay
As for ligand independent but 96-well plates were pre-coated with 50 μL of 10 μg/mL purified hJAG1 NE3 or hDLL4 NE3 in HBS overnight at 4°C, control wells were incubated without hJAG1 or hDLL4. Subsequently wells were washed with HBS, and reporter cells added. 24 h later wells were washed with HBS, and cells resuspended in 100 μL substrate solution added (150 μg/mL D-luciferin in HBS +1 mg/mL Mg^2+^, 0.1% glucose). Luciferase activity was measured immediately. For co-culture assays: B16 ligand expressing cells (either mJag1 or mDll4) were split 1:15 into RPMI +10% FCS. 48 h later, 40,000 Notch-1 expressing cells were added to each well of a white 96-well Nunc MicroWell plate (DMEM +10% FCS) and grown overnight. An equivalent number of B16 cells were added to each well and 24 h later, wells were washed with HBS, 100 μL substrate solution added and luciferase activity measured immediately.
Cis-inhibition assay
Established HEK-RBP cell lines expressing Notch-1 variants were split into 6-well plates or T25 cell culture flasks and grown to 70–80% confluency. Cells were transiently transfected with pcDNA3.1 full-length hJAG1 or pcDNA3.1 empty vector using Mirus TransIT-293 Transfection Reagent. The presence of hJAG1 in the same cell (cis) as the Notch WT/variant provides cis-inhibitory potential in these cells. 24 h after transient transfection, 40,000 cells in DMEM +10% FCS were transferred to each well and incubated overnight. An equivalent number of B16 cells were then added to each well and, after 24 h, wells were subsequently washed with HBS, and luciferase activity measured as described above.
Flow cytometry-based cell surface Notch-1 quantification
5 × 10^5^ cells expressing each Notch variant were harvested in a microcentrifuge tube and washed 2 times in 400 μL of FACS buffer (PBS, 0.1% W/V NaN_3_, 0.2% V/V FBS, sterile-filtered). Washed cells were incubated for 1 h in the dark in 100 μL FACS buffer with 2 μg/mL allophycocyanin (APC)-conjugated anti-mNotch-1 (22E5) antibody (Invitrogen #17-5765-82, initially created by Fiorini et al.81). Cells were washed 2 times with FACS buffer, resuspended in 400 μL PBS with 1 μg/mL DAPI and incubated for 10–15 min before flow cytometry analysis. All staining steps were performed in the dark with prechilled buffer solutions on ice or at 4°C. Samples were analyzed with a CytoFLEX LX flow cytometer (Beckman Coulter) with a flow rate between 200 and 1000 events/second. Data for 30,000 events were collected for each sample. DAPI fluorescence was measured with excitation at 405 nm and emission at 450 nm. APC fluorescence was measured with excitation at 638 nm and emission at 660 nm. Data were analyzed by FlowJo v10.8 Software (BD Life Sciences). For each sample, the median fluorescence intensity of APC for viable single cells was quantified and used to show the median cell surface Notch-1 level. Four biological repeats were performed for each cell line.
Quantification and Statistical analysis
X-ray crustallography data collection and refinement statistics are shown in Table 1. SAXS data collection statistics are shown in Table 3. Parametric and nonparametric tests used are indicated in each figure legend, together with (where appropriate) the post-hoc test used for multiple comparisons. Values are presented together with the mean ± SD (standard deviation). ns = not significant; ∗p < 0.05, ^∗∗^p < 0.01, ^∗∗∗^p < 0.001, ^∗∗∗∗^p < 0.0001; Statistical data were analyzed with Prism 9 (GraphPad, San Diego, CA, USA). SD is calculated using the following equation:
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Artavanis-Tsakonas S.Rand M.D.Lake R.J.Notch Signaling: Cell Fate Control and Signal Integration in Development Science 284199977077610.1126/science.284.5415.77010221902 · doi ↗ · pubmed ↗
- 2Bray S.J.Notch signalling: a simple pathway becomes complex Nat. Rev. Mol. Cell Biol.7200667868910.1038/nrm 200916921404 · doi ↗ · pubmed ↗
- 3Abby E.Dentro S.C.Hall M.W.J.Fowler J.C.Ong S.H.Sood R.Herms A.Piedrafita G.Abnizova I.Siebel C.W.Notch 1 mutations drive clonal expansion in normal esophageal epithelium but impair tumor growth Nat. Genet.55202323224510.1038/s 41588-022-01280-z 36658434 PMC 9925379 · doi ↗ · pubmed ↗
- 4Martincorena I.Fowler J.C.Wabik A.Lawson A.R.J.Abascal F.Hall M.W.J.Cagan A.Murai K.Mahbubani K.Stratton M.R.Somatic mutant clones colonize the human esophagus with age Science (New York, N.Y.)362201891191710.1126/science.aau 387930337457 PMC 6298579 · doi ↗ · pubmed ↗
- 5Blaumueller C.M.Qi H.Zagouras P.Artavanis-Tsakonas S.Intracellular cleavage of Notch leads to a heterodimeric receptor on the plasma membrane Cell 90199728129110.1016/S 0092-8674(00)80336-09244302 · doi ↗ · pubmed ↗
- 6Logeat F.Bessia C.Brou C.Le Bail O.Jarriault S.Seidah N.G.Israël A.The Notch 1 receptor is cleaved constitutively by a furin-like convertase Proc. Natl. Acad. Sci. USA 9519988108811210.1073/PNAS.95.14.81089653148 PMC 20937 · doi ↗ · pubmed ↗
- 7Pandey A.Niknejad N.Jafar-Nejad H.Multifaceted regulation of Notch signaling by glycosylation Glycobiology 31202182810.1093/glycob/cwaa 04932472127 PMC 7799146 · doi ↗ · pubmed ↗
- 8Sanchez-Irizarry C.Carpenter A.C.Weng A.P.Pear W.S.Aster J.C.Blacklow S.C.Notch Subunit Heterodimerization and Prevention of Ligand-Independent Proteolytic Activation Depend, Respectively, on a Novel Domain and the LNR Repeats Mol. Cell Biol.2420049265927310.1128/mcb.24.21.9265-9273.200415485896 PMC 522238 · doi ↗ · pubmed ↗