Gigantochloa falcihumeris (Poaceae, Bambusoideae), a New Paleotropical Woody Bamboo Species From Southwest Yunnan, China
Jian‐Wei Li, Chao‐Mao Hui, Wei‐Yi Liu, Mao‐Sheng Sun, Wan‐ling Qin, Hao‐Feng Bao, Ru‐li Zhang

TL;DR
Scientists discovered a new bamboo species in Southwest Yunnan, China, with unique leaf features and confirmed its classification using genetic analysis.
Contribution
The discovery and phylogenetic confirmation of a new woody bamboo species, Gigantochloa falcihumeris, in Southwest Yunnan.
Findings
Gigantochloa falcihumeris is distinguished by its tall culms and unique falcate appendage on leaf sheath shoulders.
Phylogenetic analysis using plastome and ITS sequences confirms the new species belongs to the Gigantochloa genus.
The species is morphologically similar to G. verticillata and G. felix but differs in several key traits.
Abstract
A new species Gigantochloa is described and illustrated from Southwest Yunnan, China. The new species is similar to G. verticillata and G. felix in general appearance, but differs by its taller culms, internodes without stripes, culm leaf sheath shoulders prominent upward forming ca. 1 cm long falcate appendage, culm blade erect, culm leaf auricle absent, and culm leaf ligule ca. 5 mm tall. Additionally, we constructed a phylogenetic tree based on plastome and ITS sequences, which revealed that the new species clusters within the Gigantochloa clade. We describe Gigantochloa falcihumeris, a new species of paleotropical woody bamboo discovered in Southwest Yunnan, China. It is distinguished by its culm leaf sheath shoulders that prominently rise into a distinctive ca. 1 cm long falcate point. Phylogenetic analyses based on plastome and ITS sequences confirm its placement within the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
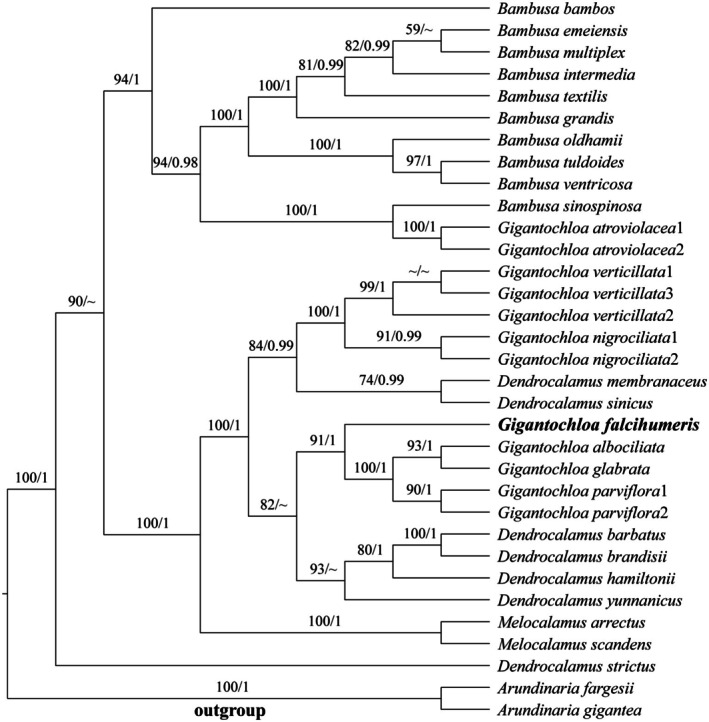 FIGURE 1
FIGURE 1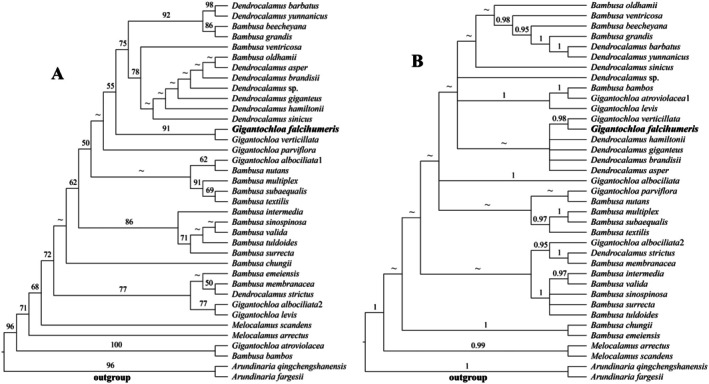 FIGURE 2
FIGURE 2 FIGURE 3
FIGURE 3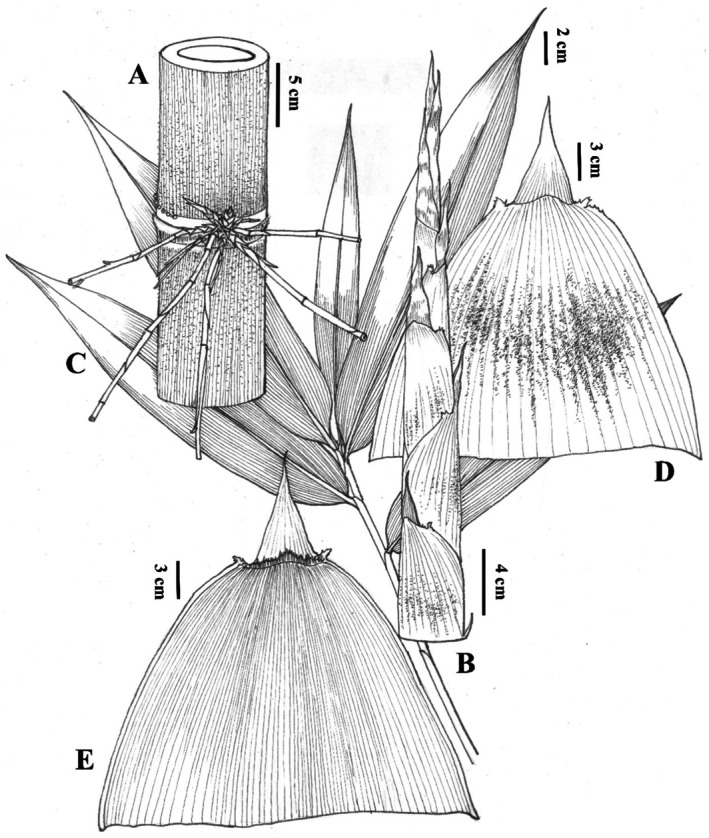 FIGURE 4
FIGURE 4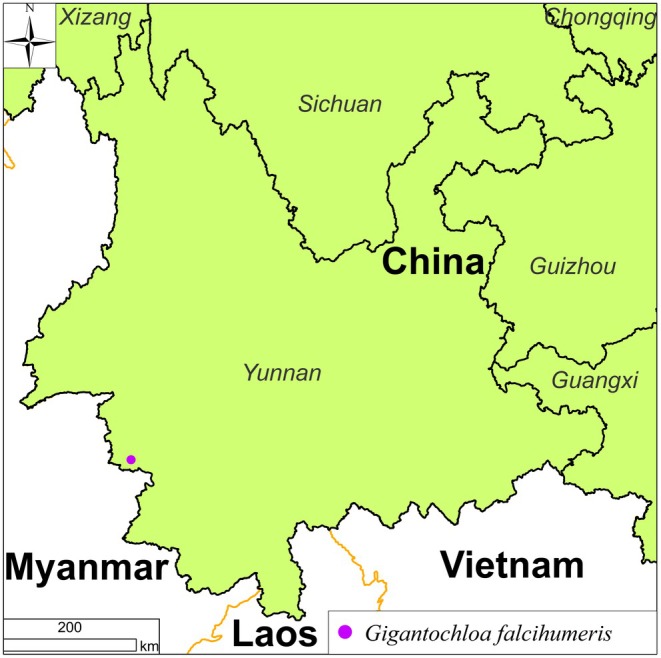 FIGURE 5
FIGURE 5| Number | Taxon | Plastome | ITS |
|---|---|---|---|
| 1 |
| ||
| 2 |
| — | |
| 3 |
| — | |
| 4 |
| ||
| 5 |
| NC068816 | |
| 6 |
| ||
| 7 |
| — | |
| 8 |
| ||
| 9 |
| — | |
| 10 |
| NC012927 | |
| 11 |
| NC050781 | |
| 12 |
| — | |
| 13 |
| — | |
| 14 |
| NC071780 | |
| 15 |
| ||
| 16 |
| — | |
| 17 |
| ||
| 18 |
| — | |
| 19 |
| ||
| 20 |
| ||
| 21 |
| — | |
| 22 |
| ||
| 23 |
| NC050766 | — |
| 24 |
| ||
| 25 |
| — | |
| 26 |
| ||
| 27 |
| ||
| 28 |
| NC050765 | |
| 29 |
| — | |
| 30 |
| ||
| 31 |
| NC050777 | — |
| 32 |
| ||
| 33 |
| — | |
| 34 |
| — | |
| 35 |
| NC050778 | — |
| 36 |
| — | |
| 37 |
| ||
| 38 |
| NC050749 | — |
| 39 |
| ||
| 40 |
| NC050779 | — |
| 41 |
| — | |
| 42 |
| ||
| 43 |
| ||
|
| |||
| 1 |
| ||
| 2 |
| — | |
| 3 |
| — | |
| Characters |
|
|
|
|---|---|---|---|
| Culms | Erect, 7.5–12 (18) cm diameter, 18–23 m tall; apex drooping | Erect, 7–10 cm diameter, 8–15 m tall; apex drooping and inclining | Nearly erect, 4–6 cm in diameter, 9–13 m tall; apex drooping |
| Internode | Without stripes | Light yellow stripes | Without stripes |
| Culms leaf sheath | Black hairs abaxially, shoulders prominent upward forming falcate appendage | White and brown setae abaxially, shoulders not prominent | Brown black setae abaxially, shoulders not prominent |
| Culm blade | Erect | Reflexed | Erect |
| Culm leaf auricle | Absent | Inconspicuous | Absent |
| Culm leaf ligule | ca. 5 mm tall, margins dentate | ca. 3 mm tall, margins fimbriate or dentate | ca. 1 mm tall, margin serrulate |
| Foliage leaf | 14–26 cm × 2.5–4 cm, both surface slightly scabrous | 24–47 cm × 3.5–7 cm, both surface glabrous | 30–45 cm × 4.5–6 cm, pubescent abaxially |
| Foliage leaf ligule | 1–2 mm tall, margin serrulate | 5–10 mm tall, margins entire | 3–4 mm tall, margins bifid or concave |
| Transverse veins | Inconspicuous | Conspicuous | Inconspicuous |
- —Lincang Forestry and Grassland Bureau
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Taxonomy and Phylogenetics · Plant Diversity and Evolution · Chromosomal and Genetic Variations
Introduction
1
In 1864, Gigantochloa was named by Kurz without morphological description (Kurz 1864). It was not until 1868 that Munro provided the first morphological description of the genus in his “Monograph of the Bambusacea” (Munro 1868), marking the formal publication of Gigantochloa; however, he did not designate a type species for the genus. In 1956, Holttum conducted a systematic study of Gigantochloa and formally designated G. atter as the type species of the genus (Holttum 1956), thereby fully establishing Gigantochloa as a taxonomic unit.
The typical morphological characteristics of Gigantochloa include: Culms erect, sometimes scrambling; internodes setose, basal internodes with light yellow‐white stripes; sheath scars prominent; culm nodes flat. Branches many, dominant branches conspicuous. Culm leaves deciduous, densely setose abaxially; auricles inconspicuous; ligules conspicuous; blades erect or reflexed. Foliage leaves many per branchlet; auricles absent; ligules conspicuous; blades medium to large (Keng and Wang 1996; Li et al. 2006; Yi et al. 2008, 2017). Currently, approximately 63 species of Gigantochloa have been recognized (Vorontsova et al. 2017), which are mainly distributed in the rainforests from Southeast Asia to South Asia (Keng and Wang 1996, Li et al. 2006, Yi et al. 2008, 2017). Among these, 9 species have been recorded in China, with seven species found in Yunnan Province (Yi et al. 2017). In recent years, new species of Gigantochloa continue to be discovered and published in the Yunnan region (Xu et al. 2021; Qin et al. 2023).
Gigantochloa is a paleotropical woody bamboo and is closely related to Bambusa Schreber and Dendrocalamus Nees (Liu et al. 2020). Bambusa can be readily distinguished from the other two genera by vegetative characteristics such as culm leaf blade erect, auricle conspicuous, and having branch thorn. In contrast, Dendrocalamus and Gigantochloa are highly similar in morphology, both exhibit culm leaf auricle inconspicuous, ligule well‐developed, and blade erect or reflexed (Keng and Wang 1996; Li et al. 2006; Yi et al. 2008, 2017). Except for certain Gigantochloa species that can be identified by yellow stripes on internodes, accurate identification generally relies on reproductive organs such as flowers and fruits. However, given the rarity of bamboo flowering, molecular systematics has become an essential supplementary approach in taxonomic studies of these groups (Triplett 2023; Wang et al. 2023; Zhang, Zhang, et al. 2024; Chen et al. 2025a; Li et al. 2025).
In 2022, during a bamboo resource survey in Lincang City, Yunnan Province, we discovered a bamboo with distinctive morphological characteristics; it exhibits 1 or without dominant branch, culm leaf auricle absent, ligule prominent, blade erect, and without branch thorn. These characteristics align with the morphological definitions of both Gigantochloa and Dendrocalamus. However, its culm leaf sheath shoulder is prominent upward, forming a ca. 1 cm long falcate appendage, which is highly distinctive. As no inflorescence specimens were collected, it was difficult to determine its taxonomic status based solely on vegetative organs. To address this, we collected fresh and healthy leaf samples for total genomic DNA extraction and constructed a phylogenetic tree to clarify its systematic position. The results showed that it clusters within the Gigantochloa clade with strong statistical support, leading us to identify it as a new species of Gigantochloa, which is hereby formally described.
Materials and Methods
2
Field Work, Specimen Collection, and Morphological Comparison
2.1
Observations and measurements of living plants were conducted in natural habitat. Specimens were examined under a stereomicroscope (AOSV HD206), while fresh and healthy leaves were collected and dried in silica gel for molecular analyses. The morphological characteristics of closely related species were primarily referenced from the authoritative literature monographs (Keng and Wang 1996; Li et al. 2006; Yi et al. 2008, 2017) and newly described species (Xu et al. 2021). Additionally, specimens deposited in the Herbarium of Southwest Forestry University (SWFC), Herbarium of Kunming Institute of Botany, Chinese Academy of Sciences (KUN), and Herbarium of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences (HITBC), among other herbaria, were examined; at the same time, we also conducted field observations on species of Gigantochloa in the Bamboo Garden of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences.
DNA Extraction, Sequencing, and Assembly
2.2
Total genomic DNA was extracted from silica gel‐dried leaves using the TIANGEN Magnetic Plant Genomic DNA Kit (TIANGEN, Beijing, China). DNA quality was assessed for degradation, contamination, and concentration by the Agilent 5400 system. Qualified DNA was sheared into approximately 350 bp fragments with a Covaris focused ultrasonicator, followed by end polishing, A‐tailing, ligation of full‐length Illumina sequencing adapters, and PCR amplification. PCR products were accurately quantified using real‐time PCR (3 nM) to determine the effective library concentration. Eligible libraries were sequenced on the Illumina platform with PE150 strategy, generating approximately 2 GB of raw data per sample. All the aforementioned sequencing experiments were conducted at Novogene Bioinformatics Technology Co. Ltd. in Beijing, China.
The raw data were quality‐controlled using Fastp 0.19.7 (Chen et al. 2018) to remove low‐quality sequences. Plastome and nuclear ribosomal DNA (nrDNA) sequence of the new species were assembled using GetOrganelle (Jin et al. 2020) with default parameters. Subsequently, the plastome sequence was visualized using Bandage (Wick et al. 2015), and the assembled sequence was analyzed for collinearity using Mauve (Darling et al. 2004). Finally, Plastome sequence was annotated using CPGAVAS2 (Shi et al. 2019), and annotations were manually adjusted in Geneious Prime 2022.0.1 (Kearse et al. 2012) based on the annotation information of Dendrocalamus strictus (Roxb.) Nees (NCBI: MK679802). The nrDNA sequences of the new species were assembled and annotated in Geneious Prime 2022.0.1 (Kearse et al. 2012) with reference to D. strictus (Roxb.) Nees (NCBI: JX139103). ITS sequences of the new species were accurately extracted using ITSx (Bengtsson‐Palme et al. 2013).
Construct Phylogenetic Tree
2.3
Phylogenetic trees of the new species were reconstructed using both Maximum Likelihood (ML) and Bayesian inference (BI) based on plastome and ITS sequences. Sequences of related species were retrieved from the NCBI database, comprising a total of 46 sequences from 40 species (Table 1). Sequence alignment was performed using MAFFT (Katoh et al. 2002). For ML analyses, optimal nucleotide substitution models were identified using ModelFinder (Kalyaanamoorthy et al. 2017) based on the Bayesian Information Criterion (BIC). Specifically, K3Pu + F + R4 and TPM2u + G4 were determined as the best‐fit models for plastome and ITS datasets, respectively. ML analysis was conducted with IQ‐TREE 2.2.5 (Nguyen et al. 2015), with branch support assessed through 1000 ultrafast bootstrap replicates and SH‐aLRT tests (Hong et al. 2022). BI was carried out in MrBayes 3.2.7a (Ronquist et al. 2012) under the GTR + I + G model, which was selected via BIC using jModelTest 2.1.7 (Darriba et al. 2012). Two independent Markov Chain Monte Carlo (MCMC) runs were performed for 10,000,000 generations, sampling every 1000 generations. The first 25% of samples were discarded as burn‐in, and a 50% majority‐rule consensus tree was generated when the average standard deviation of split frequencies dropped below 0.01. Nodal support was evaluated based on posterior probabilities (Ronquist et al. 2012).
Results
3
Morphological Comparisons
3.1
Gigantochloa falcihumeris is morphologically similar to G. verticillata and G. felix, but can be clearly distinguished by differences in key morphological characters such as the taller culms, internodes without stripes, culm leaf sheath shoulders prominent upward forming ca. 1 cm long falcate appendage, culm blade erect, culm leaf auricle absent, and culm leaf ligule ca. 5 mm tall (Table 2).
Phylogenetic Analysis
3.2
Upon alignment, the total length of the plastome sequence was 102,776 bp, including 2017 variable sites and 1384 parsimony information sites. In the plastome phylogenetic tree, the clustering of various species was relatively poor: two sequences of Gigantochloa atrovilacea were nested within the Bambusa clade, while Dendrocalamus membranaceus and D. sinicus were grouped with Gigantochloa species, and other D. strictus formed a distinct monophyletic clade (Figure 1). Despite the relatively disorganized phylogenetic relationships among species in the plastome tree, the new species within a clade containing species of Gigantochloa (MLBP/BI = 91/1) was most closely related to G. albociliata, G. glabrata and G. parviflora (Figure 1).
Phylogenetic tree reconstructed by Maximum Likelihood (ML) analysis based on plastome sequences. Numbers along branches indicate the Maximum Likelihood bootstrap values (MLBP) (left) and Bayesian posterior probabilities (BI) (right). “~”: nodes with MLBP < 50% (left), Bayesian posterior probabilities (BI) < 95% (right).
The total length of the ITS sequences was 285 bp, containing 182 variable sites and 104 parsimony‐informative sites. In the ITS phylogenetic tree, the interspecific evolutionary relationships were more disorganized than those in the plastome tree. Additionally, the topologies generated by ML and BI methods showed inconsistencies; however, both methods consistently grouped the new species with G. verticillata on the same clade with strong support (MLBP = 91, BI = 0.98) (Figure 2).
Phylogenetic tree reconstructed from ITS sequences. (A) Maximum Likelihood (ML) method. (B) Bayesian Inference (BI) method. Numbers along branches indicate the Maximum Likelihood bootstrap values (MLBP) (A) and Bayesian posterior probabilities (BI) (B). “~”: nodes with MLBP < 50% (A), Bayesian posterior probabilities (BI) < 95% (B).
Taxonomy
4
Gigantochloa falcihumeris C.M.Hui & J.W.Li sp. nov. (Figures 3I and 4).
(I) Gigantochloa falcihumeris: (A) Habitat, (B) culm fragment showing nodal line, (C) culm bud, (D) new shoot showing culm leaves, (E) falcate appendage, (F) culm leaf sheath shoulders prominent, (G) culm leaf ligule, (H) bamboo shoot and falcate appendage, (I) branch complement. (II) Gigantochloa felix: (A) clump, (B) branch complement, (C) culm leaves, (D) foliage leaves. (III) Gigantochloa verticillata : (A) clump, (B) internodes yellow striate, (C) culm leaves, (D) culm leaf ligule, (E) foliage leaves.
Gigantochloa falcihumeris: (A) Branch complement, (B) bamboo shoot, (C) foliage leaf, (D) culm leaf, adaxial view, (E) culm leaf, abaxial view.
Type
4.1
CHINA. Yunnan: Lincang City, Cangyuan County, Menglai Township, Papeng Village, in sparse forests, N23°11′ 57.40″, E99°14′ 52.33″, elev. ca. 1380 m, 24 September 2024, C. M. Hui, J. W. Li, H. F. Bao & C. H. Zhang 0072417(holotype: SWFC!).
Diagnosis
4.2
Gigantochloa falcihumeris is morphologically similar to G. verticillata and G. felix, but can be distinguished by the following characteristics: Culms 7.5–12 (18) cm in diameter, 18–23 m tall (vs. 7–10 cm in diameter, 8–15 m tall in G. verticillata ; 4–6 cm in diameter, 9–13 m tall in G. felix); internodes without stripes (vs. light yellow stripes in G. verticillata ); culm leaf sheath shoulders prominent upward forming ca. 1 cm long falcate point (vs. shoulders not prominent in G. verticillata and G. felix); culm blade erect (vs. reflexed in G. verticillata ); culm leaf auricle absent (vs. inconspicuous in G. verticillata ); culm leaf ligule ca. 5 mm tall (vs. ca. 3 mm tall in G. verticillata ; ca. 1 mm tall in G. felix) (Table 2).
Description
4.3
Rhizomes simpodial. Culms erect, the apex drooping, 7.5–12 (18) cm in diameter, 18–23 m tall; internodes 30–38 (40) cm long, without stripes, initially covered with white hairs; intranode ca. 1 cm tall, with a ring of white tomentum and aerial roots; culm nodes flat, glabrous; sheath scar slightly prominent with dense brown setae. Branch developing from higher nodes; branches several with 1 or without dominant branch. Culm leaves deciduous, leathery; sheaths 28–35 cm × 33–42 cm, equal to or slightly shorter than internodes, abaxially white powdery and covered with deciduous black hairs initially, mid‐upper with a ca. 5 mm wide thin marginal membrane, shoulders prominent upward forming ca. 1 cm long falcate appendage with margins serrate; blades erect, 8–12 cm long, rugose, sparsely white powdery, margins involute; auricle absent; ligule ca. 5 mm tall, margins dentate. Foliage leaves 6–10 per ultimate branch; blade elliptic‐lanceolate, thin‐textured, 14–26 cm × 2.5–4 cm, both surface slightly scabrous, secondary veins 8–12 pairs, transverse veinlets inconspicuous; pseudopetiole 3–5 mm long, glabrous; sheath 4–9 cm long, covered with procumbent black hairs; auricle absent; ligule 1–2 mm tall, glabrous, margin finely dentate. Inflorescence and caryopsis unknown.
Phenology
4.4
New shoots June to September.
Distribution and Habitat
4.5
Gigantochloa falcihumeris was recently found near Papeng Village at an elevation of 1200–1400 m in Menglai Township, Cangyuan County, Southwestern Yunnan, China (Figure 5). It coexists with other species, such as Lithocarpus fenestratus (Roxb.) Rehder, Castanopsis indica (Roxb. ex Lindl.) A. DC., Dendrocalamus sinicus L.C. Chia & J.L. Sun, D. brandisii (Munro) Kurz, and Castanopsis hystrix Hook. f. & Thomson ex A. DC.
Geographical distribution of Gigantochloa falcihumeris C.M.Hui & J.Wei Li.
Etymology
4.6
The specific epithet “falcihumeris” refers to the culms leaf sheath shoulders prominently upward forming falcate appendage.
Chinese Name
4.7
zhí gǎn jù zhú (Chinese pronunciation); 直杆巨竹(Chinese name); yě duō wǎ (local Lahu language).
Local Use
4.8
The local Lahu people refer to the new species as “yě duō wǎ” (meaning “rafter bamboo”). Due to its straight culms and moderate size, it has traditionally been used as rafters for building houses. Additionally, the local Lahu people also use its bamboo shoots to make pickled sour bamboo shoots.
Conservation Status
4.9
Gigantochloa falcihumeris is so far only known from its type locality, with a population of approximately five clumps, and the number of mature individuals in other similar habitats remains unknown. It is threatened by potential habitat disturbance and a lack of comprehensive surveys to define its full distribution. It can be categorized as “Data Deficient” (DD) following the IUCN categories and criteria (IUCN 2024).
Additional Specimens Examined
4.10
CHINA. Yunnan: Lincang City, Cangyuan County, Menglai Township, Papeng Village, in sparse forests, N23°11′ 57.40″, E99°14′ 52.33″, elev. ca. 1380 m, 24 September 2024, C. M. Hui, J. W. Li, H. F. Bao & C. H. Zhang, 0072418, 0072419, 0072420(SWFC!).
Discussion
5
Due to the exceptionally long flowering cycles of bamboo plants (Zheng et al. 2020; Chen et al. 2025b), their classification and identification primarily rely on morphological characteristics of vegetative organs (such as culms, culm sheaths, and foliage leaves). This approach has been commonly used for establishing new bamboo species in traditional taxonomy (Yi 1992, 2001; Wen 2001; Du et al. 2013; Zeng et al. 2014; Chen et al. 2013; Zhang et al. 2019; Ye et al. 2020, 2021; Shi et al. 2024; Zhang, Cao, and Ding 2024). In recent years, the integration of molecular phylogenetic data has provided further critical evidence for delineating new bamboo species (Triplett 2023; Wang et al. 2023; Zhang, Zhang, et al. 2024; Chen et al. 2025a; Li et al. 2025).
Through morphological comparison and phylogenetic reconstruction, this study identifies an unknown bamboo species as a new member of Gigantochloa. However, we constructed phylogenetic trees based on plastome and ITS sequences respectively, and found that the evolutionary relationships revealed by different datasets were significantly different. Specifically, the ML tree and BI tree constructed from plastome data exhibited consistent topological structures; in contrast, the phylogenetic trees based on ITS data not only showed inconsistent topologies but also had more complex branching relationships than the plastome trees. Nevertheless, both types of phylogenetic trees placed the new species within the Gigantochloa clade. Previous studies have generally adopted the approach of constructing trees using multiple datasets separately to comprehensively determine the systematic position when establishing new bamboo species (Zhang, Zhang, et al. 2024; Chen et al. 2025b; Li et al. 2025). This indicates that judging the taxonomic affiliation of a new species solely based on a phylogenetic tree constructed from a single dataset is insufficient and may lead to one‐sided conclusions.
Furthermore, this study also found that neither the phylogenetic trees constructed based on plastome nor ITS sequences exhibited a clear and stable evolutionary relationship structure at the species level. This phylogenetic ambiguity is likely related to the “Bambusa–Dendrocalamus–Gigantochloa (BDG) complex”—a group primarily composed of genera such as Bambusa, Dendrocalamus, Gigantochloa, and Melocalamus (Zhou et al. 2017). This complex represents the most diverse and phylogenetically challenging group within the paleotropical woody bamboos (PWB) of the grass subfamily Bambusoideae and has long presented a major challenge in bamboo systematics research (Sun et al. 2005; Yang et al. 2010; Goh et al. 2013; Zhou et al. 2017; Liu et al. 2020, 2024). Although significant progress has been made in recent studies (Liu et al. 2020, 2024), the intergeneric boundaries and evolutionary relationships within this complex remain not yet fully resolved.
According to records from Flora Reipublicae Popularis Sinicae (1996), Flora of China (2006), Iconographia Bambusoidearum Sinicarum (2008), and Illustrated Flora of Bambusoideae in China (2017), species of Gigantochloa in China are primarily distributed in the Xishuangbanna Prefecture of southern Yunnan, with very sporadic records in Dehong Prefecture. The new species discovered in Cangyuan County in southwestern Yunnan, which not only adds a new species to Gigantochloa but also expands the known geographic distribution range of Gigantochloa in China. Situated on the southwestern border of China and adjacent to Myanmar, Cangyuan County exhibits strong Southeast Asian floristic characteristics in its plant vegetation. However, due to its remote location and the constraints of local economic development, biodiversity baseline surveys remain relatively limited, resulting in many species yet to be documented. Enhanced field investigations in this region should be conducted in the future to further enrich the understanding of both Chinese and global biodiversity.
Author Contributions
Jian‐Wei Li: conceptualization (lead), formal analysis (lead), investigation (lead), writing – original draft (lead). Chao‐Mao Hui: data curation (equal), formal analysis (lead), funding acquisition (lead), methodology (equal), project administration (equal), resources (lead), supervision (equal), writing – review and editing (lead). Wei‐Yi Liu: conceptualization (equal), methodology (equal), supervision (equal), writing – review and editing (equal). Mao‐Sheng Sun: formal analysis (equal), methodology (equal). Wan‐ling Qin: investigation (equal). Hao‐Feng Bao: investigation (equal). Ru‐li Zhang: methodology (equal).
Funding
This work was supported by the National Key Research and Development Project of China (2021YFD2200501), the Agriculture Joint Special Project of Yunnan Province (202301BD070001‐123), the National‐level high‐quality variety “Yuntian No. 1” high‐standard cultivation technology promotion demonstration (YUN [2024]TG 11 No), the Monitoring Project of Bamboo Forest Ecosystem Positioning Observation and Research Station in South Yunnan, China (2025–YN–15), and the Sci‐Tech Service Station of Farmer Academician in Lancang and Changning City.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bengtsson‐Palme, J. , M. Ryberg , M. Hartmann , et al. 2013. “Improved Software Detection and Extraction of ITS 1 and ITS 2 From Ribosomal ITS Sequences of Fungi and Other Eukaryotes for Analysis of Environmental Sequencing Data.” Methods in Ecology and Evolution 4, no. 10: 914–919. 10.1111/2041-210X.12073. · doi ↗
- 2Chen, M. , Y. J. Chen , M. Y. Zhou , et al. 2025 b. “Syntenic Nuclear Genes and Morphology Reveal Unusual New Climbing Bamboo Species of Melocalamus in the Gaoligong Mountains.” Taxon 74, no. 6: 1–17. 10.1002/tax.13385. · doi ↗
- 3Chen, S. , Y. Zhou , Y. Chen , and J. Gu . 2018. “fastp: An Ultra‐Fast All‐In‐One FASTQ Preprocessor.” Bioinformatics (Oxford, England) 34, no. 17: 884–890. 10.1093/bioinformatics/bty 560.30423086 PMC 6129281 · doi ↗ · pubmed ↗
- 4Chen, S. H. , R. S. Chen , K. F. Huang , and H. Z. Guo . 2013. “ Dendrocalamus longiauritus S.H.Chen, K.F.Huang et R.S.Chen, a New Dendrocalamus Species of Bambusoideae From China.” Plant Science Journal 31, no. 6: 536–539.
- 5Chen, Y. J. , M. Chen , M. Y. Zhou , et al. 2025 a. “Further Investigation of Melocalamus (Poaceae, Bambusoideae) in China Based on Skmer Analysis and Morphology.” Phyto Keys 259: 177–198. 10.3897/phytokeys.259.151683.40642314 PMC 12242230 · doi ↗ · pubmed ↗
- 6Darling, A. C. , B. Mau , F. R. Blattner , and N. T. Perna . 2004. “Mauve: Multiple Alignment of Conserved Genomic Sequence With Rearrangements.” Genome Research 14, no. 7: 1394–1403. 10.1101/gr.2289704.15231754 PMC 442156 · doi ↗ · pubmed ↗
- 7Darriba, D. , G. L. Taboada , R. Doallo , and D. Posada . 2012. “j Model Test 2: More Models, New Heuristics and Parallel Computing.” Nature Methods 9: 772. 10.1038/nmeth.2109.PMC 459475622847109 · doi ↗ · pubmed ↗
- 8Du, F. , Y. M. Yang , and J. Wang . 2013. “ Cephalostachyum virulentum, a New Species of Cephalostachyum From West Yunnan.” Journal of West China Forestry Science 42: 10–12.