Identification of SORD as A Key Gene Mediating Osimertinib Primary Resistance in EGFR-Mutant Lung Adenocarcinoma via Machine Learning and Methylomics
Guowei LIANG, Hongfeng WU, Chaoyi JIA, Penghu GAO, Zhanrui ZHANG, Chen CHEN, Yongwen LI, Hongyu LIU, Jun CHEN

TL;DR
This study identifies SORD as a key gene linked to primary resistance to the drug osimertinib in lung adenocarcinoma patients with EGFR mutations, using machine learning and methylation data.
Contribution
The study introduces SORD as a novel biomarker for predicting osimertinib resistance and proposes targeting the SORD pathway as a new strategy to overcome resistance.
Findings
SORD methylation levels are negatively correlated with its mRNA expression in EGFR-mutant lung adenocarcinoma.
Modulating SORD expression reverses osimertinib resistance in resistant and sensitive cell lines.
Targeting SORD methylation could serve as a potential therapeutic strategy to overcome drug resistance.
Abstract
肺腺癌(lung adenocarcinoma, LUAD)是非小细胞肺癌中最常见的亚型,而表皮生长因子受体(epidermal growth factor receptor, EGFR)突变是其主要的分子驱动事件。尽管第三代酪氨酸激酶抑制剂(trrosine kinase inhibitors, TKIs)药物奥希替尼已成为该类患者的标准一线疗法,但耐药性的产生严重限制了患者的长期生存获益。证据表明,表观遗传重塑是导致耐药的重要非遗传机制。其中,DNA启动子区的高甲基化可通过沉默关键抑癌基因或代谢调节基因,协助肿瘤细胞逃避药物杀伤。本研究旨在通过机器学习和高通量筛选,挖掘调控奥希替尼敏感性的关键基因,并解析甲基化修饰在其调控奥希替尼原发性耐药中的作用。 基于癌症基因组图谱(The Cancer Genome Atlas, TCGA)-LUAD EGFR突变队列,应用oncoPredict模型预测奥希替尼的半数抑制浓度(half inhibitory concentration, IC50)。通过多组学关联分析,分别筛选与IC50显著相关的转录组与甲基化组差异基因。根据“高甲基化-低表达”负相关模式对两组学数据取交集,并应用LASSO回归与Bootstrap验证筛选出最稳健的核心基因。最后,在EGFR突变LUAD细胞系中,通过药物敏感性测定、去甲基化处理及基因过表达/敲低实验,验证核心基因对奥希替尼耐药、细胞增殖及凋亡的影响。 多组学分析与机器学习算法鉴定出SORD为核心候选基因,其关键位点(cg06424894)甲基化水平与mRNA表达呈显著负相关。体外实验显示,奥希替尼耐药株H1650呈SORD高甲基化与低表达特征,去甲基化药物处理可显著恢复其表达;敏感株则呈相反趋势。功能实验证实,在H1650耐药细胞中过表达SORD可逆转耐药,抑制细胞增殖并促进凋亡。相反,在H1975和PC9敏感细胞中敲低SORD,则显著诱导耐药,促进增殖并抑制凋亡。…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
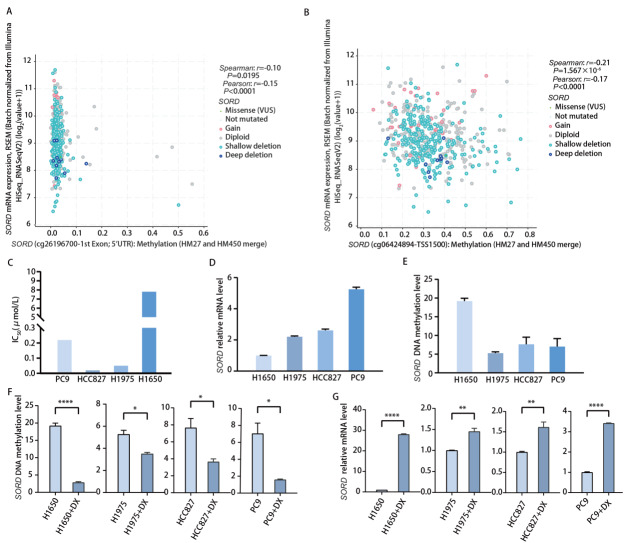 图 1
图 1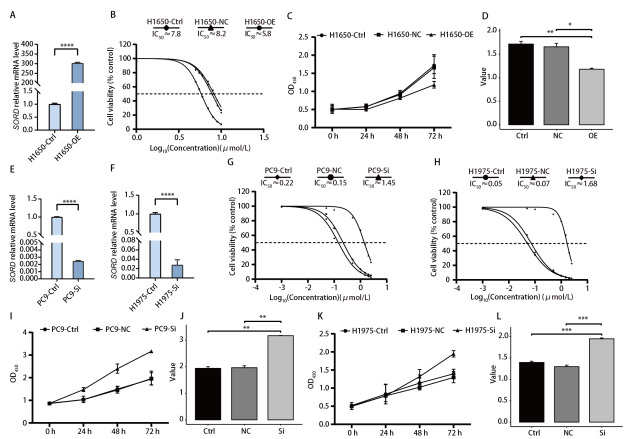 图 2
图 2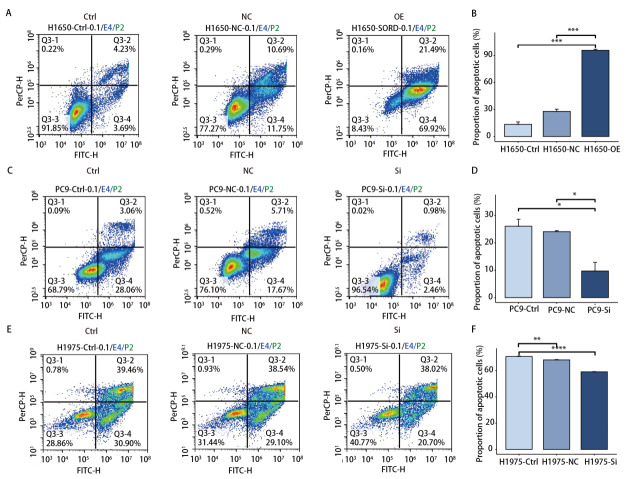 图 3
图 3- —国家自然科学基金项目
- —National Natural Science Foundation of China
- —国家自然科学基金项目
- —National Natural Science Foundation of China
- —天津市医学重点学科建设项目
- —Tianjin Key Medical Discipline Construction Project
- —天津市医学重点学科建设项目
- —Tianjin Key Medical Discipline Construction Project
- —天津市自然科学基金重点项目
- —Natural Science Foundation of Tianjin
- —天津市自然科学基金重点项目
- —Natural Science Foundation of Tianjin
- —北京市科技创新医学发展基金项目
- —Beijing Science and Technology Innovation Medical Development Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
肺腺癌(lung adenocarcinoma, LUAD)是非小细胞肺癌(non-small cell lung cancer, NSCLC)最主要的亚型,占所有肺癌病例的40%-50%,是全球癌症相关死亡的主要原因之一^[1]^。在LUAD患者中,表皮生长因子受体(epidermal growth factor receptor, EGFR)突变是一个关键的驱动基因,尤其在非吸烟的亚洲女性患者中,EGFR突变率高达50%-60%^[2]^。对于EGFR敏感突变(如外显子19缺失或L858R点突变)的NSCLC患者,第三代EGFR-酪氨酸激酶抑制剂(EGFR-tyrosine kinase inhibitors, EGFR-TKIs)奥希替尼已成为标准的一线治疗方案^[3]^。尽管奥希替尼显著改善了患者的无进展生存期和总生存期,但其疗效仍面临严峻挑战。这些挑战不仅包括几乎所有初始治疗敏感的患者在治疗1-2年内出现的获得性耐药,也包括一部分具有EGFR敏感突变的患者从治疗开始就表现出的原发性耐药^[4]^。目前,关于奥希替尼的获得性耐药机制研究已大量开展,C797S二级突变、间质上皮细胞转化因子(mesenchymal-epithelial transition factor, MET)扩增或突变以及旁路激活、组织学转化等是导致奥希替尼获得性耐药的重要机制^[5]^。然而,对于原发性耐药的机制,特别是基于表观遗传学调控在奥希替尼原发性耐药中的作用机制,目前的理解仍然不足。
DNA甲基化是一种重要的表观遗传修饰方式,这种修饰不改变DNA序列,却能稳定遗传并调控基因表达。在肿瘤中,启动子区异常高甲基化常导致抑癌基因沉默,进而影响DNA修复和凋亡通路^[6]^。近年研究^[7]^发现,特定基因的甲基化状态可直接介导EGFR-TKIs获得性耐药,正成为预测疗效和克服耐药的重要表观遗传标志物与潜在治疗靶点。此外,异常的DNA甲基化,如抑癌基因启动子区的高甲基化,可导致其表达沉默,进而影响药物敏感性,被认为是EGFR-TKIs原发性耐药的可能潜在机制之一^[8]^。
近年来,深度学习与机器学习算法为整合高通量多组学数据提供了有力工具,这尤其依赖于对大型公共数据库,如癌症基因组图谱(The Cancer Genome Atlas, TCGA)和肿瘤药物敏感性基因组学数据库2(Genomics of Drug Sensitivity in Cancer 2, GDSC2)等的挖掘和分析。oncoPredict等工具^[9]^则是利用这类数据,通过机器学习方法,建立样品对药物敏感性的预测工具。尽管oncoPredict等工具能有效预测药物敏感性,但对于耐药机制的解析仍然缺乏。此外,对于表观遗传调控(如甲基化)与药物敏感性之间的相互作用探讨仍不充分。
本研究通过整合TCGA-LUAD队列的转录组和甲基化数据,利用oncoPredict等模型量化伴有EGFR突变患者的奥希替尼的半数抑制浓度(half inhibitory concentration, IC_50_),筛选受DNA甲基化调控并可能导致EGFR突变LUAD患者原发性耐药的关键基因,并对筛选出的关键基因SORD在具有EGFR突变的LUAD细胞中的功能进行初步探讨,旨在揭示其调控LUAD细胞对奥希替尼敏感性的潜在机制,为识别新型耐药生物标志物和开发个性化治疗策略提供新的思路。
1 资料与方法
1.1 数据获取及基因突变分析
本研究中有关LUAD患者的数据来源于cBioPortal数据库(https://www.cbioportal.org)中处理好的LUAD数据[name: Lung Adenocarcinoma (TCGA, PanCancer Atlas), Reference: TCGA, Cell 2018],总共纳入了566个LUAD样本,包括转录组数据以及相应的临床信息。
使用cBioPortal数据库获取TCGA-LUAD患者EGFR基因突变信息,突变包含点突变、缺失突变、插入突变等。
1.2 EGFR突变LUAD患者对奥希替尼药物敏感性的计算及药敏相关基因筛选
提取TCGA中具有EGFR突变的LUAD患者对应的RNA-seq数据,使用oncoPredict(v1.2)包并结合GDSC2数据库,得到每例患者对应的IC_50_。
构建多阶段筛选策略以精准筛选核心基因。首先,结合Spearman相关性分析与Limma模型,对转录组及甲基化数据进行初步过滤,筛选IC_50_相关基因。随后,应用带有10折交叉验证的LASSO回归进一步去除冗余,实现特征降维。为确保结果的稳健性,我们引入Bootstrap重抽样技术进行了10,000次迭代验证,最终保留在多次模拟中均表现稳定的关键基因。
1.3 细胞培养
细胞系H1650、H1975、HCC827、PC9购自中科院典藏生物资源保藏中心。这些细胞系在含有10%胎牛血清的DMEM(诺普赛,PM150210)培养基、37 °C和5% CO_2_的湿润培养箱中培养。
1.4 细胞凋亡检测
为评估细胞在奥希替尼压力下的凋亡情况,药物刺激48 h后,收集上清中的悬浮细胞及贴壁细胞,使用0.25%胰蛋白酶/EDTA,37 °C消化5 min,以冷PBS洗涤2次。随后将细胞在缓冲液中与FITC Annexin V和PI共同孵育25 min。最后,使用NovoCyte 2000R流式细胞仪进行分析并处理数据。
1.5 细胞增殖实验(CCK-8法)
首先,将待测细胞按3000个/100 µL的密度接种于96孔板中,每组设置至少3个复孔。接种后将培养板置于细胞培养箱中过夜以确保细胞完全贴壁,期间尽量避免晃动以保证细胞分布均匀。次日,弃除孔内原培养基,按预设浓度加入新鲜配制的药液处理48 h。处理结束后,吸除含药培养基,每孔加入100 µL含10% CCK-8的培养基。随后将培养板用锡纸包裹避光孵育1 h,最后利用酶标仪在450 nm波长下检测各孔的吸光度(OD值)。
1.6 细胞转染与RNA干扰
对于SORD基因的过表达,过表达质粒购自质粒与蛋白共享库,使用线性聚乙烯亚胺转染试剂(MW 40,000)按照说明书进行转染。对于SORD敲低,使用了2条针对人SORD基因的siRNA混合物。序列分别为:5’-CAGAAUCCCUGAUGUUAAU-3’; 5’-GAUCAUCGGUAAAGCACCU-3’。使用Lipofectamine 2000(Invitrogen)按照标准步骤将siRNA池转染至细胞。转染48 h后收集细胞进行后续分析。
1.7 荧光定量聚合酶链式反应(polymerase chain reaction, PCR)
使用胰蛋白酶消化并收集细胞,使用SPARKeasy Cell RNA快速提取试剂盒(Sparkjade,中国山东)提取总RNA。测量RNA浓度后,取2 µg总RNA,使用逆转录试剂盒(Takara,中国北京)将其逆转录为cDNA。荧光定量PCR在SLAN-96P全自动PCR分析仪上进行。所有基因的表达水平均以GAPDH作为内参进行归一化。本研究所使用的SORD引物序列如下:F:5’-GCCGATACAATCTGTCACCTTCC-3’,R:5’-CGCCTTCCTCAAAGGTGACATTG-3’。
1.8 DNA亚硫酸氢盐转化及甲基化特异性PCR引物设计
提取各组样本的基因组DNA,使用DNA亚硫酸氢盐转化试剂盒(碧云天,编号:D0068S)对DNA进行转化处理。转化后的DNA存储于-80 ^o^C。针对SORD基因启动子区域的CpG岛,利用MethPrimer在线工具(http://www.urogene.org/methprimer/)设计甲基化特异性引物(M)和非甲基化特异性引物(U)。引物序列如下:*SORD*-M-L:5’-TATTTAAGTTTTAGTAATGGCGGAC-3’,*SORD*-M-R:5’-TATACCTAACTCAATAAATCCCACG-3’;*SORD*-U-L:5’-TTTAAGTTTTAGTAATGGTGGATGT-3’,*SORD*-U-R:5’-TACCTAACTCAATAAATCCCACACC-3’。采用荧光定量PCR法检测各组样本的甲基化水平。根据甲基化引物与非甲基化引物扩增所得Ct值(分别为Ct_M和Ct_U),按如下公式计算各样品中*SORD*基因的相对甲基化率:[2^-Ct_M^/(2^-Ct_M^+2^-Ct_U^)]×100%。
1.9 统计分析
所有统计分析均在R语言环境(v4.3.3)中进行。数据表示为均数±标准差(Mean±SD),两组间差异比较采用双尾非配对Student's t检验。P<0.05被认为差异具有统计学意义。
2 结果
2.1 基于机器学习预测TCGA-LUAD队列中EGFR突变患者的奥希替尼IC50
为预测LUAD患者对奥希替尼的敏感性,本研究利用cBioPortal平台对TCGA-LUAD数据集中的566例原始病例进行分析。以EGFR突变为筛选标准,共鉴定出79例携带EGFR突变的患者,其EGFR突变以经典激活突变为主,其中L858R点突变20例(23.3%)、外显子19 E746_A750del缺失14例(16.3%);伴随或独立EGFR扩增21例(24.4%)。其余为少见激活突变(L861Q 3例、G719A/C 3例、S768I 2例)及罕见插入/缺失(E709_T710delinsD 3例、L747-T751del 2例、L747_A750delinsP 2例、A767_V769dup 1例、H773dup 1例、K754_I759del 1例、T751_I759delinsN 1例),并检测到耐药相关T790M 2例。余为散发突变或意义未明变异。
采用这79例样本的RNA-seq数据作为机器学习模型的分子层面输入信息。进一步选用oncoPredict作为核心计算算法,并以GDSC2作为模型预测的参考标准,通过oncoPredict计算,获取全部79例EGFR突变患者对奥希替尼的IC_50_预测值。结果显示,39例样本的IC_50_值高度密集于2.00-5.00 μmol/L区间,该群体涵盖了大部分L858R(10例)及E746_A750del(5例)突变样本;极敏感区间(<0.68 μmol/L)则富集了K754I、759del等突变。尽管L858R和E746_A750del通常被视为奥希替尼的经典敏感突变,但我们的预测结果显示,除L833F/L861Q等非经典突变外,仍有4例L858R和2例E746_A750del样本IC_50_落入高IC_50_(>10.00 μmol/L)抗性区间。随后,我们对IC_50_值进行了log_2_转换以利于后续统计分析。
2.2 结合RNA-seq与TCGA甲基化数据筛选药敏相关基因
为了进一步挖掘与奥希替尼IC_50_相关的关键基因,首先,我们提取患者的RNA-seq数据与奥希替尼IC_50_值进行相关性分析,共筛选出1173个与IC_50_显著相关的基因,其中呈正相关的基因1100个,呈负相关的基因73个。
同样的,我们对79例患者中70例拥有DNA甲基化数据的患者样本,采用类似的相关性筛选标准,分析基因甲基化水平与IC_50_的相关性。此分析共获得2652个基因甲基化水平与IC_50_显著相关的基因,其中正相关基因118个(甲基化水平越高,患者对应的奥希替尼IC_50_值越高,提示耐药相关),负相关基因2534个(甲基化水平越高,IC_50_值越低)。
为了进一步锁定受甲基化调控且与IC_50_显著相关的核心基因,基于表观遗传学中“DNA甲基化水平与基因表达呈负相关”的机制,进行筛选。具体筛选策略如下:首先,取DNA甲基化分析中与IC_50_显著正相关的“高甲基化基因”与转录组分析中与IC_50_显著负相关的“低表达基因”的交集,最终获得36个高甲基化且低表达且与IC_50_显著相关的候选基因(附图1, http://www.lungca.org/files/2025s192s1.pdf)。
2.3 基于LASSO回归筛选奥希替尼原发性耐药相关高甲基化基因
我们进一步应用LASSO回归模型,对上述36个高甲基化且低表达的基因进行特征筛选。结果显示,所构建的预测模型对药物敏感性(log_2_(IC_50_))具有良好的预测性能,预测值与实际值的Pearson相关系数为0.738,决定系数为0.544。为进一步验证模型的稳健性并挖掘关键基因,我们采用了10,000次Bootstrap重抽样策略。结果显示,12个基因的入选频率超过50%,表明它们是与IC_50_强相关的稳定特征。这些基因包括SORD、NEURL3、FIGN、ZNF280B、LANCL2、LYPD6、WDFY2、ZNF418、BEND3、TIAM1、POM121和RNF220。值得注意的是,排名靠前的基因在功能上均高度富集于癌症进展的关键通路:SORD是代谢重编程的关键酶,FIGN是调控微管动态和有丝分裂的重要蛋白,ZNF280B作为转录因子参与p53信号通路的调控,而TIAM1则介导Rac1信号通路以影响细胞迁移。在所有稳定入选的基因中,代谢酶SORD的选择频率最高(0.7859),提示SORD是与药物敏感性(IC_50_)关联最密切的核心候选基因(附图2,http://www.lungca.org/files/2025s192s2.pdf)。
2.4 SORD基因表达受甲基化调控
为确认SORD为受甲基化调控基因,我们通过cBioPortal平台工具分析TCGA-LUAD甲基化数据集。结果显示,SORD关键甲基化位点甲基化水平与其mRNA表达呈显著负相关,特别是cg06424894位点(TSS1500)相关性尤为显著(Spearman: r=-0.21, P=1.567×10^-6^; Pearson: r=-0.17, P=7.462×10^-5^)(图1A、1B)。同时,TCGA样本分析显示SORD基因突变频率极低,排除了基因突变作为影响其表达水平的因素。
候选基因SORD的表达验证及其与甲基化的相关性分析。A:SORD基因甲基化位点(cg26196700-1st Exon; 5'UTR)与其mRNA表达水平的相关性散点图;B:SORD基因甲基化位点(cg06424894-TSS1500)与其mRNA表达水平的相关性散点图;C:H1650、HCC827、H1975和PC9细胞系对奥希替尼的药物敏感性检测(IC50);D:不同细胞系中SORD基因的基础mRNA表达水平;E:不同细胞系中SORD基因的基础甲基化水平;F:地西他滨处理显著降低了不同细胞系中SORD基因的甲基化水平;G:地西他滨处理引起的甲基化水平降低显著影响了不同细胞系中SORD mRNA的表达变化。
为了在体外水平进行验证,我们采用4种LUAD细胞株(H1650、H1975、HCC827、PC9),这4种LUAD细胞株均具有EGFR突变,其中H1650、PC9、HCC827均携带EGFR外显子19缺失突变,H1975携带EGFR L858R与T790M双突变。我们首先评估了4种细胞株的奥希替尼敏感性。结果显示,H1650细胞对奥希替尼表现为原发性耐药(IC_50_约7.8 μmol/L),而其他3种细胞(H1975、HCC827、PC9)则均为敏感细胞系(IC_50 _≤0.22 μmol/L)(图1C)。
SORD基因的甲基化特异性PCR检测显示,耐药的H1650细胞呈现出SORD基因高甲基化状态,mRNA表达水平低;与此相反,3种敏感细胞系则均表现为低甲基化状态,SORD mRNA表达水平高(图1D、1E)。
4种细胞株进一步采用30 μmol/L去甲基化药物地西他滨处理48 h。结果显示,地西他滨可显著提升其mRNA表达,H1650细胞SORD mRNA水平相较于原始表达提升20倍以上(图1F、1G),支持SORD是一个受甲基化调控的基因。
2.5 SORD基因表达可影响EGFR突变LUAD细胞对奥希替尼的敏感性
为验证SORD基因对奥希替尼疗效的直接影响,本研究进行了细胞表型实验。首先,在耐药细胞系H1650中过表达SORD基因(图2A),IC_50_测定结果表明,SORD过表达显著逆转了H1650细胞的耐药性,IC_50_值从7.8 μmol/L降低至5.8 μmol/L(图2B)。在5 μmol/L奥希替尼处理下,CCK8实验显示SORD过表达显著抑制了细胞的增殖能力(与对照组相比,P=5.60×10^-3^)(图2C、2D)。
调控SORD基因表达对非小细胞肺癌细胞奥希替尼耐药性的影响。A:通过质粒转染在H1650细胞中过表达SORD基因;B:过表达SORD基因后H1650细胞对奥希替尼敏感性的变化曲线;C、D:评估过表达SORD基因后细胞在药物作用下的增殖能力变化;E、F:通过siRNA转染在PC9和H1975细胞中敲低SORD基因的表达;G、H:敲低SORD基因后PC9和H1975细胞对奥希替尼敏感性的变化曲线;I-L:敲低SORD基因后细胞在药物作用下的增殖能力变化。
同时,在敏感细胞系H1975和PC9中敲低SORD基因(图2E、2F),结果显示,SORD敲低显著增加细胞耐药性:PC9细胞系IC_50_从0.22 μmol/L升至1.45 μmol/L,H1975细胞系IC_50_从0.05 μmol/L升至1.68 μmol/L(图2G、2H)。在0.05 μmol/L低浓度奥希替尼作用下,CCK8实验亦表明SORD敲低显著提升了细胞的增殖能力(PC9组P=0.0022,H1975组P=0.00018)(图2I-2L)。
为进一步探究SORD对细胞凋亡的影响,我们在H1650过表达细胞中检测了5 μmol/L奥希替尼处理48 h后的凋亡情况。结果表明,SORD过表达显著提升了细胞凋亡率(P=1.70×10^-4^)(图3A、3B)。同样,在H1975-Si SORD和PC9-Si SORD细胞中,0.05 μmol/L奥希替尼处理48 h后,SORD敲低显著抑制了细胞凋亡(PC9组P=1.70×10^-2^,H1975组P=8.1×10^-6^)(图3C-3F)。
干扰SORD基因表达对奥希替尼诱导的细胞凋亡的影响。A、B:H1650细胞过表达SORD基因后,流式细胞术检测在奥希替尼处理下的细胞凋亡情况;C、D:PC9细胞敲低SORD基因后,流式细胞术检测在奥希替尼处理下的细胞凋亡情况;E、F:H1975细胞敲低SORD基因后,流式细胞术检测在奥希替尼处理下的细胞凋亡情况。
3 讨论
奥希替尼虽显著改善了EGFR突变LUAD患者的预后^[10]^,但原发性耐药仍是主要的临床瓶颈。DNA甲基化作为关键的表观遗传修饰,已被证实可通过沉默抑癌基因或激活耐药通路影响EGFR-TKIs疗效^[11]^,并促进耐药相关通路的激活^[12]^。因此,挖掘受甲基化调控的关键耐药节点,对于开发新型生物标志物及增敏策略具有重要意义。
与既往研究主要依赖细胞系模型或单一组学数据不同,本研究创新性地构建了“药物反应预测-多组学交互-机器学习筛选”的整合分析框架。首先,利用oncoPredict模型在TCGA临床队列中克服了药物反应数据缺失的难题;其次,通过转录组与甲基化组的交集分析,精准锁定了“表观遗传沉默”特征基因;最后,应用LASSO回归与Bootstrap重抽样验证,确保了筛选结果的稳健性与可靠性。这一严谨的筛选流程使我们最终锁定了SORD作为与奥希替尼耐药最相关的核心基因,避免了单一分析方法可能产生的假阳性偏差。
SORD编码的山梨醇脱氢酶是多元醇通路的关键限速酶^[13]^,该通路通过醛糖还原酶AKR1B1与SORD催化的两步酶促反应,实现葡萄糖向果糖的转化^[14]^。因此,现有研究多聚焦于其在糖尿病并发症中的病理作用^[15]^以及周围神经病变^[16]^,而该基因在肺癌耐药进程中的表观遗传调控机制则鲜有报道。本研究结果显示,SORD启动子区域的过度甲基化与其mRNA表达沉默呈显著负相关,这一发现在耐药细胞株H1650中得到了体外验证。 这一结果直接提示,DNA甲基化是导致SORD在耐药株中失活的主要机制。功能实验进一步证实,SORD并非单纯的“旁观者”,而是耐药表型的“驱动者”:在耐药株中恢复SORD表达可显著逆转耐药并诱导凋亡,而在敏感株中敲低SORD则重现了耐药表型。这表明SORD的缺失可能通过破坏细胞内的山梨醇/果糖代谢平衡,改变细胞氧化还原状态或能量代谢流,从而赋予癌细胞在药物压力下的生存优势^[17]^。与METTL3介导的m6A修饰等^[18]^其他机制相比,本研究突出了DNA甲基化驱动的特定代谢基因沉默在奥希替尼耐药中的独特作用。
总之,本研究通过多组学整合分析与机器学习算法,不仅筛选了SORD甲基化在奥希替尼耐药中的调控价值,也为开发广谱性TKIs耐药逆转策略提供了新思路。在NSCLC治疗已进入分子分型指导的精准时代,靶向表观遗传的疗法正成为克服靶向治疗耐药的重要方向,而本研究为这一领域的药物研发提供了新的潜在靶点。
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sung H, Ferlay J, Siegel RL, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin, 2021, 71(3): 209-249. doi: 10.3322/caac.21660 33538338 · doi ↗ · pubmed ↗
- 2Borgeaud M, Olivier T, Bar J, et al. Personalized care for patients with EGFR-mutant nonsmall cell lung cancer: Navigating early to advanced disease management. CA Cancer J Clin, 2025, 75(5): 387-409. doi: 10.3322/caac.70024 40673977 PMC 12432819 · doi ↗ · pubmed ↗
- 3Ramalingam SS, Vansteenkiste J, Planchard D, et al. Overall survival with Osimertinib in untreated, EGFR-mutated advanced NSCLC. N Engl J Med, 2020, 382(1): 41-50. doi: 10.1056/NEJ Moa 1913662 31751012 · doi ↗ · pubmed ↗
- 4Mok TS, Wu YL, Ahn MJ, et al. Osimertinib or Platinum-Pemetrexed in EGFR T 790M-positive lung cancer. N Engl J Med, 2017, 376(7): 629-640. doi: 10.1056/NEJ Moa 1612674 27959700 PMC 6762027 · doi ↗ · pubmed ↗
- 5Leonetti A, Sharma S, Minari R, et al. Resistance mechanisms to osimertinib in EGFR-mutated non-small cell lung cancer. Br J Cancer, 2019, 121(9): 725-737. doi: 10.1038/s 41416-019-0573-8 31564718 PMC 6889286 · doi ↗ · pubmed ↗
- 6Lin L, Liu Y. Advances in epigenomic sequencing and their applications in cancer diagnostics. Precis Chem, 2025, 3(10): 581-603. doi: 10.1021/prechem.5c 00014. 41170156 PMC 12569958 · doi ↗ · pubmed ↗
- 7Koch A, Joosten SC, Feng Z, et al. Analysis of DNA methylation in cancer: location revisited. Nat Rev Clin Oncol, 2018, 15(7): 459-466. doi: 10.1038/s 41571-018-0004-4 29666440 · doi ↗ · pubmed ↗
- 8Cao-Lei L, Veru F, Elgbeili G, et al. DNA methylation mediates the effect of exposure to prenatal maternal stress on cytokine production in children at age 13½ years: Project Ice Storm. Clin Epigenetics, 2016, 8: 54. doi: 10.1186/s 13148-016-0219-0 27182285 PMC 4866030 · doi ↗ · pubmed ↗