Microbial diversity and potential pathogens associated with the plastisphere on beaches of Rio de Janeiro, Brazil
Luana Gomes Morais, Henrique Fragoso dos Santos, Douglas Alfradique Monteiro, Marina Montenegro Mascarenhas, Helena Antunes Portela, Mônica Regina da Costa Marques, Fábio Vieira de Araújo

TL;DR
This study explores the microbial communities on plastic waste from Rio de Janeiro beaches, identifying potential pathogens and plastic-degrading bacteria.
Contribution
The study identifies potential pathogens and plastic-degrading bacteria in the plastisphere and evaluates the impact of plastic type on microbial community formation.
Findings
Microbial biofilms on plastic waste contain potentially pathogenic bacteria and plastic-degrading species.
No specific associations were found between polymer types (PP and PE) and microbial communities.
Plastic waste poses risks to human health and marine biodiversity due to microbial colonization.
Abstract
Marine pollution caused by plastic waste is a global issue. Improperly discarded plastics often end up in seas and oceans, where they become colonized by a microbial biofilm known as the Plastisphere. In this environment, especially in places where sewage is discharged untreated into water bodies, pathogenic microorganisms and those carrying antimicrobial resistance genes (ARGs) can settle and spread over long distances. To assess the presence of these microorganisms and examine the impact of substrate type on the formation of these communities, plastic waste samples were collected from three beaches in Rio de Janeiro, Brazil. The plastics were identified as polypropylene (PP) and polyethylene (PE) using FTIR analysis, and the microbial biofilm attached to them was studied through 16 S rRNA gene sequencing. The results revealed the presence of bacteria potentially pathogenic to humans…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
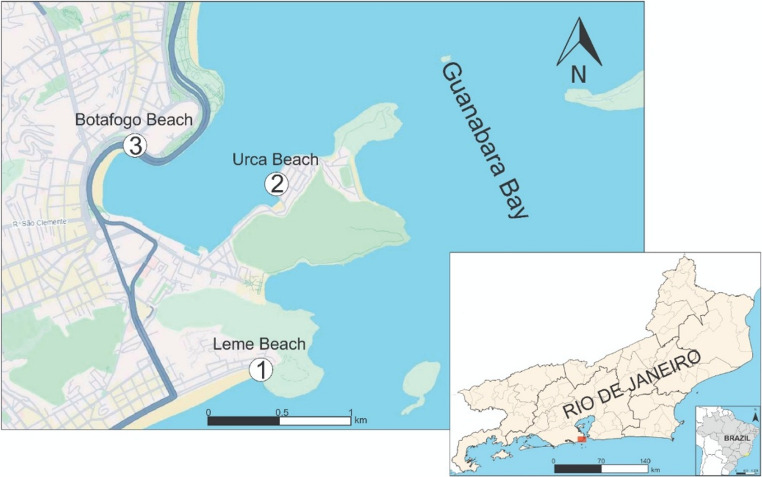 Figure 1
Figure 1 Figure 2
Figure 2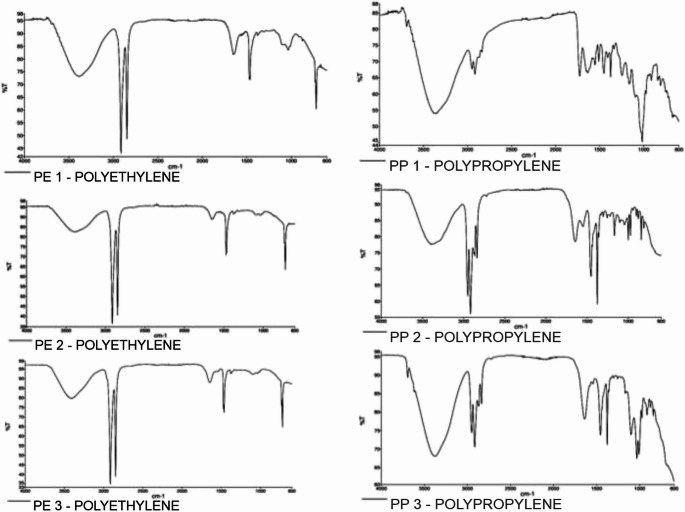 Figure 3
Figure 3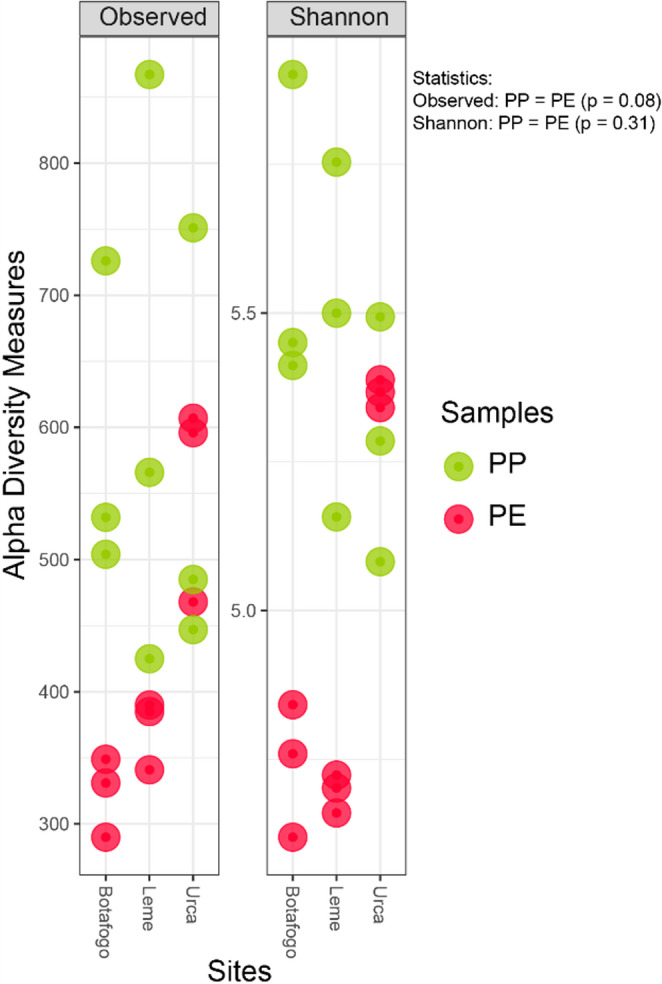 Figure 4
Figure 4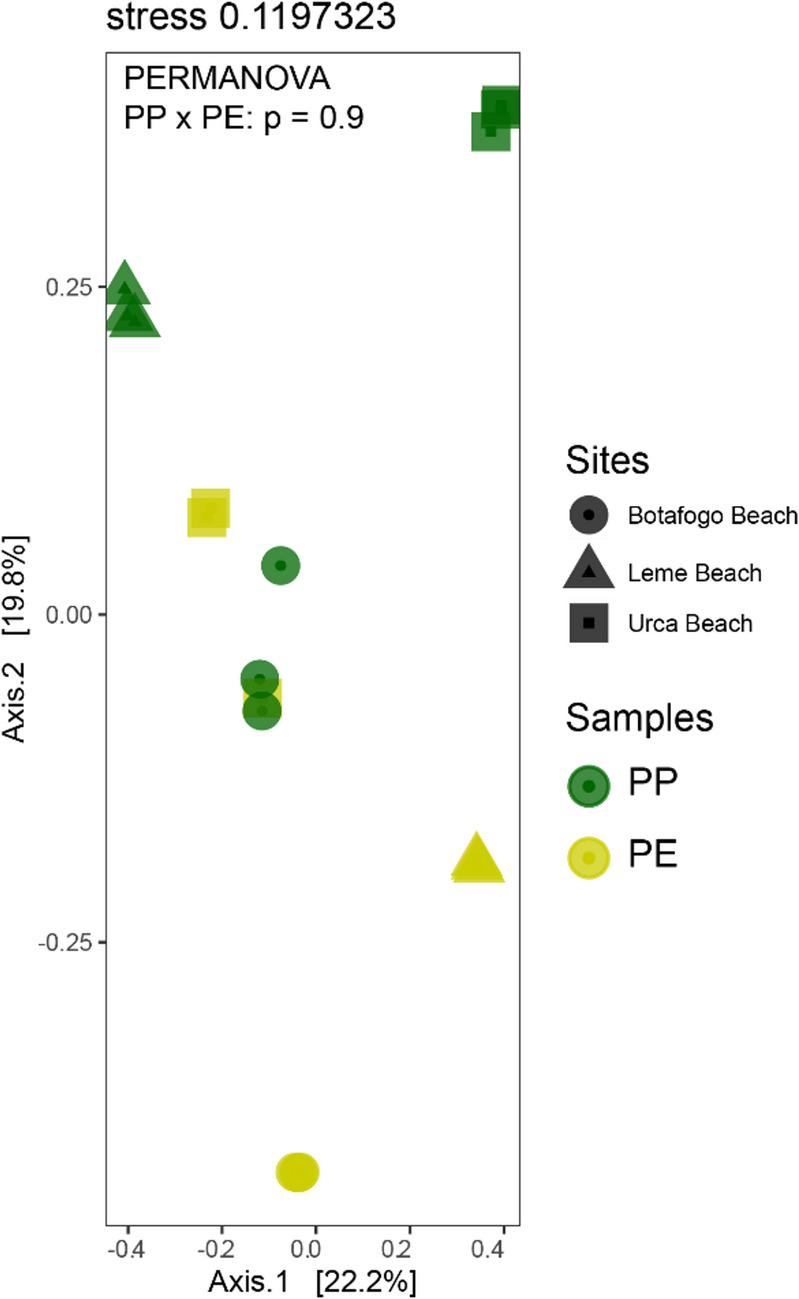 Figure 5
Figure 5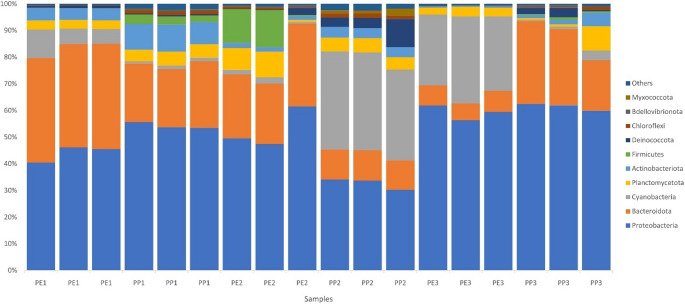 Figure 6
Figure 6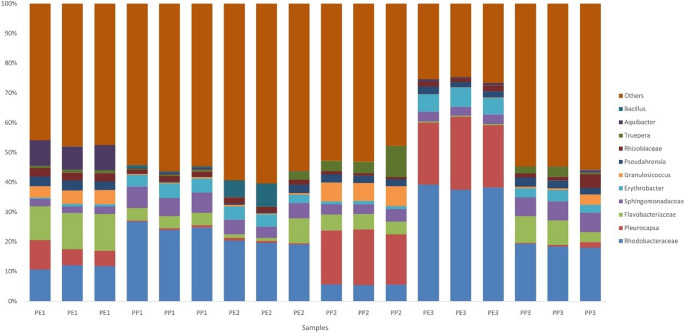 Figure 7
Figure 7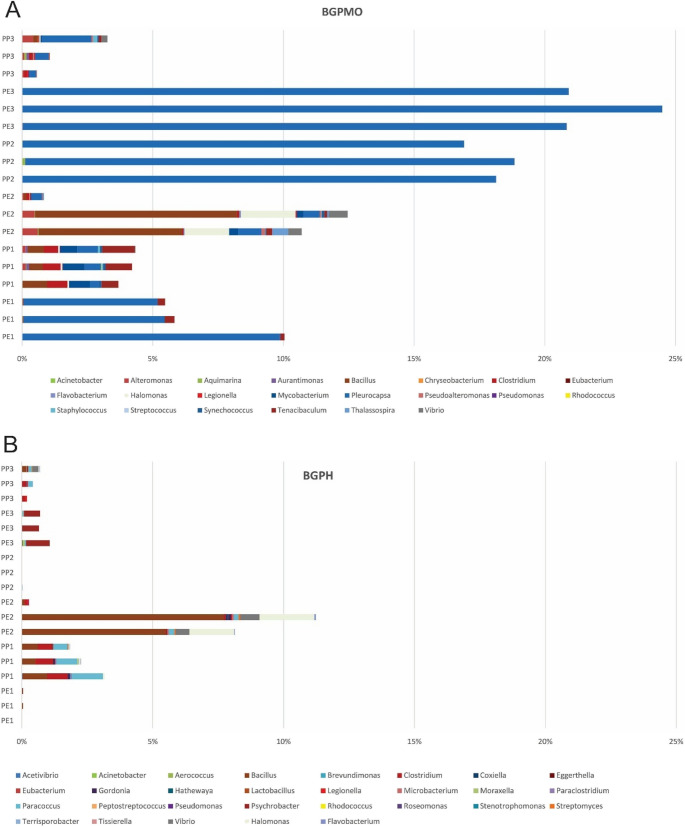 Figure 8
Figure 8- —Universidade Do Estado Do Rio De Janeiro
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicroplastics and Plastic Pollution · biodegradable polymer synthesis and properties · Antimicrobial agents and applications
Introduction
Global plastic production is projected to reach 460 million tons by 2030 [1], with approximately 22% of this total being improperly discarded [2], contributing to the increasing accumulation of plastic waste in marine environments. Jacquin et al. [3] report that between 4.8 and 12.7 million tons of plastic enter the ocean annually due to improper urban waste disposal, coastal tourism, and maritime activities. This pollution has become a global issue, significantly impacting coastal and marine environments [4].
Once in the ocean, these pollutants rapidly disperse over long distances due to ocean currents and winds, facilitating their deposition in coastal areas [5, 6]. Plastic in the marine environment poses numerous problems for organisms, including entanglement and accidental ingestion, which can cause illness and death in animals such as turtles, birds, fish, and marine mammals [7, 8].
Due to characteristics such as longevity, resilience, and hydrophobicity, drifting plastic debris serves as a persistent artificial surface for microbial colonization [9], and surface roughness and the size of the debris can accelerate this process [10]. This colonization leads to the formation of a biofilm, known as the Plastisphere [11].
The formation of biofilms on the surface of plastics is important for the degradation process [12], as it can protect the material from UV radiation, thereby reducing the direct action of photocatalysis or indirectly by decreasing its buoyancy [13], which increases its persistence in marine debris. On the other hand, studies show that microorganisms can accelerate the degradation of these materials [9].
The Plastisphere is also responsible for the dispersion of microorganisms, some of which are potentially pathogenic [14], and others that carry antimicrobial resistance genes [15, 16] throughout the ocean. The presence of these microorganisms in biofilms formed on plastic debris in the ocean enhances the role of water as a vehicle for the transmission of various diseases. In this new environment, microorganisms may encounter conditions that increase their viability and promote gene transfer mechanisms, thereby raising the risk for a significant portion of the population that relies on coastal environments for recreation and subsistence [4].
Moreover, microorganisms present in the Plastisphere represent a key category of life to understanding and mitigating the potential adverse effects of plastics due to their role in driving the overall functioning of the marine biosphere and as possible mediators of the biodegradation of associated plastic additives, contaminants, or even the plastics themselves [17]. Therefore, examining the formation, structure, and functions of biofilms related to this type of waste is crucial to inform management strategies aimed at preserving the ecological integrity of marine ecosystems.
Globally, research has focused on the composition, quantification, and distribution of marine plastic waste; however, much less attention has been given to the microbial communities colonizing these materials, particularly in tropical environments. These regions often encompass developing countries with limited sanitation infrastructure, where pathogenic microorganisms and carriers of antimicrobial resistance genes—once present in waters receiving untreated effluents—can be transported by plastic waste to previously unpolluted areas [18, 19].
Thus, this study aimed to characterize the bacterial community in the Plastisphere of plastic waste with distinct polymer compositions found on urban beaches in Rio de Janeiro. The study tested the following hypotheses: (i) potentially pathogenic microorganisms and plastic-degrading species are present in the Plastisphere of plastics with distinct polymer compositions; (ii) the polymer composition of plastic waste influences the composition of the Plastisphere.
Our study holds significant value as it sheds light on microbial communities associated with plastic waste in a region previously unexplored in this context. While several studies have examined the Plastisphere in temperate environments, little is known about microbial communities colonizing plastics in tropical urban bays subjected to severe sewage contamination, such as Guanabara Bay, one of the most polluted coastal systems in South America.
Methodology
Study areas
Rio de Janeiro, considered the largest coastal city in Latin America, with a population of 6,750,000 inhabitants, is renowned worldwide for its natural landscapes and is a highly sought-after tourist destination. Despite this, some municipalities in their metropolitan region lack sanitation infrastructure, leading to pollution of the waters of Guanabara Bay, which receives effluent from several of these municipalities. The selected study sites include the beaches of Leme, Urca, and Botafogo, located in the urban area of Rio de Janeiro, RJ (Fig. 1). These beaches were chosen as representative sites due to their distinct pollution gradients.
Fig. 1. Map showing the collection sites: Leme (thermotolerant coliform < 1,000/100mL), Urca (thermotolerant coliform ≥ 1,000–10,000/100 mL), and Botafogo (thermotolerant coliform > 10,000/100 mL) beaches in Rio de Janeiro (RJ), Brazil. The sampling sites were designated as 1 (Leme Beach), 2 (Urca Beach), and 3 (Botafogo Beach)
Leme is an oceanic beach near the entrance to Guanabara Bay, offering favorable conditions for swimming nearly all the time (thermotolerant coliform levels below 1,000/100mL), according to INEA [20]. It has a high flow of bathers and is frequented by various vendors along its sands. Urca and Botafogo beaches, which exhibit thermotolerant coliform counts of 1,000–10,000/100 mL and > 10,000/100 mL, respectively, are in Guanabara Bay, a region known for its high levels of pollution. Although both beaches have calm waters, Botafogo exhibits poorer water quality than Urca, as it is in a more sheltered area with reduced water circulation. While the water at Urca is not always suitable for swimming, it is frequented by bathers, unlike Botafogo beach [20]. Despite this, both beaches serve as locations for recreational activities on their sands.
Sampling
The collections were carried out on March 28, 2022, under moderate winds and no precipitation, according to the National Institute of Meteorology (INMET). The collection began at Leme Beach at 14:00 (Latitude: −22.9635213, Longitude: −43.1649918), followed by Urca Beach at 15:30 (Latitude: −22.9471088, Longitude: −43.1634273), and concluded at Botafogo Beach around 16:30 (Latitude: −22.9434220, Longitude: −43.1772478). Plastic waste, such as disposable cups and bags, was collected from each beach using sterile gloves to prevent contamination. Some waste showed signs of degradation, while others were covered with algae (Fig. 2). Using tweezers and scissors previously sterilized with 70% ethanol, smaller fragments of the waste were placed into sterile 15 mL Falcon tubes and stored in refrigerated containers (0–4 °C) to preserve the biofilms until arrival at the laboratory, where they were transferred to a freezer (−20 °C) until analysis.
Fig. 2. Plastic cups (A, C, and E) and plastic bags (B, D, and F) collected at Leme Beach (A, B), Urca Beach (C, D), and Botafogo Beach (E, F)
Sample processing
Before analysis, samples were thawed and gently rinsed with sterile distilled water to remove surface residues. From each sample, one fragment was aseptically collected to analyze its polymer chemical composition. Three other fragments were taken for molecular analysis in triplicate. Each fragment was placed in an Eppendorf tube, resulting in three subsamples per waste type (technical replicates) and a total of 18 samples for molecular analysis.
Polymer composition
The samples were analyzed for their polymer composition using the methodology described in Castro et al. [21], employing a PerkinElmer infrared spectrometer (model Spectrum 3) coupled to the Spotlight 200i microscope (also from PerkinElmer) at the Chemistry Institute of UERJ. The analysis was conducted in attenuated total reflectance (ATR) mode with a Germanium (Ge) crystal, with a wavenumber range of 4000–600 cm⁻¹ and scans equal to 16. Analyses were performed using a Zinc Selenide (ZnSe) slide as a support medium. PerkinElmer Polymers Library was used to validate the results.
Bacterial community analysis
The fragments intended for molecular analysis from the different samples were cut into tiny pieces and immersed in liquid nitrogen within a mortar for freezing. During the freezing process, these fragments were pulverized with a pestle. 0.5 g of the material from this procedure was transferred to tubes containing glass beads, provided in the Qiagen extraction kit.
DNA extraction was performed using the “PowerSoil Isolation Kit” (©Qiagen USA - www.qiagen.com.br) following the manufacturer’s instructions. DNA quality and concentration were evaluated by fluorimetry using a Qubit fluorometer (ThermoFisher - USA). For sequencing purposes, the variable V4-V5 region of the 16 S rRNA gene was targeted and amplified using the primers 515 F and 909R [22]. The amplicons were sequenced using the Illumina MiSeq platform (Illumina, San Diego, CA, USA) by StarSeq GmbH (Mainz, Germany).
After sequencing, the generated reads were processed using bioinformatics software. Subsequent analyses, including diversity, taxonomy, and classification, were conducted using RStudio software.
The raw data were demultiplexed and imported into the QIIME2 pipeline [23]. The DADA2 plugin [24] was utilized to assess read quality and cluster them into Amplicon Sequence Variants (ASVs). The taxonomic classification of each ASV was accomplished using the feature-classifier plugin with a Bayesian classifier trained with the SILVA 138 database [25]. Singletons, ASVs classified as Eukaryota and Archaea, off-targets, and organellar DNA (chloroplast and mitochondria) were discarded. Rarefaction curves were generated to evaluate the adequacy of sequencing depth.
Statistical analysis
Alpha diversity analysis, including observed ASVs and Shannon index [26], was calculated using the R programming language (version 3.6.3) [27].
For beta diversity analysis, which examined variations in sample composition between plastic polymer type, multidimensional scaling (MDS) was employed [28], using the Phyloseq package [29]. R software generated an ordering among all communities through the Bray-Curtis distance. The differences between the bacterial profiles identified in each sample type were analyzed using the t-test and PERMANOVA tests, assuming p < 0.05 [30], after averaging the technical replicates per site for each polymer type.
Estimation of potentially pathogenic bacteria
The identified bacterial genera were compared with a set of 292 genera potentially associated with human diseases (BGPH) and 72 genera potentially linked to diseases in marine organisms (BGPMO), as reported by Magalhães et al. [31] and Jurelevicius et al. [32]. For beta diversity analysis, which examined variations in sample composition between plastic types, multidimensional scaling (MDS) was employed by Wickelmaier [28], using the Phyloseq package [29].
Results
Polymer composition
FTIR analysis revealed that the spectra of plastic bags are similar, indicating that they originate from the same polymer. The presence of absorption bands only for the CH2 group (2917, 2847, 1471, and 720 cm⁻¹) suggests that the polymer was polyethylene (PE). Plastic cups also exhibited similar spectra, corresponding to polypropylene (PP). Figure 3 shows the spectra acquired for each sample after analysis.
Fig. 3. Image with the spectra resulting from the FTIR-ATR analyses. On the left from top to bottom, samples PE1, PE2, and PE3 (polyethylene), respectively, and on the right, PP1, PP2, and PP3 (polypropylene) in the same order
The bacterial community associated with the samples
The richness and diversity of the bacterial community found in the samples are illustrated in Fig. 4. The “Observed” section of Fig. 4 illustrates the microbial richness within the plastic waste samples from each beach. The community diversity in the Plastisphere is depicted in the “Shannon” section. These graphs reveal that PP samples exhibit the same richness (t-test = 2.35, p = 0.08) and diversity (t-test = 1.15, p = 0.31) as PE.
Fig. 4. Alpha diversity plot showing the richness and diversity of the microbial community present in the plastic bag (PE) and plastic cup (PP) samples from each beach (left side). In the graph, individual samples of PE (red) and PP (green) refer to the richness and diversity of the microbial community on each beach
The nMDS analysis (Fig. 5) illustrates the distribution of bacterial profiles from plastic samples, categorized by polymer composition. The two primary axes explain 22.2% (Axis 1) and 19.8% (Axis 2) of the total variance, with a stress value of 0.119, indicating a good fit of the data in the two-dimensional space. Even though the figure indicates spatial separation among PP and PE, PERMANOVA indicates no statistical difference (F = 0.58, p = 0.9).
Fig. 5. Non-metric multidimensional scaling (nMDS), where the profile of the plastic samples is evaluated through the classification by location and polymer composition. The colors green and yellow represent, respectively, polypropylene (PP) and polyethylene (PE). The symbols circle, triangle, and square are the collection sites
Figure 6 displays the microbial phyla found in plastic waste, showing their abundance at each location and polymer type. Proteobacteria were predominant at all collection sites, particularly in Leme (PE1 and PP1) and Botafogo Beaches (PE3 and PP3). Bacteroidota was the second most abundant phylum, with a higher prevalence observed in the samples from Urca (PE2 and PP2). Cyanobacteria were prominent in PE3 and PP2. Planctomycetota appeared at all collection sites and polymer types with moderate but consistent abundance. Actinobacteriota were more evident in PP1, while Firmicutes and Deinococcota were more prominent in PE2 (Fig. 6). Although not very visible or abundant, it is noteworthy that Bdellovibrionota were present in both polymers, PE and PP, varying in abundance across different collection sites, being more noticeable in PP3, PP1, and PE2.
Fig. 6. Relative abundance of microbial phyla found in plastic waste. The sample identification represents the polymer composition under analysis (PE for polyethylene and PP for polypropylene), followed by the beach from which the samples were collected (1- Leme, 2- Urca, 3- Botafogo)
Figure 7 depicts the relative abundance at the genus and family levels for each plastic waste sample and collection site. The category labeled "Other" in the graph includes various other genera identified in the analysis, albeit in smaller proportions (< 2%).
Fig. 7. Relative abundance of the microbial taxonomic groups (genus and family level) at the different collection sites and waste types. The sample identification represents the polymer composition under analysis (PE for polyethylene and PP for polypropylene), followed by the beach from which the samples were collected (1- Leme, 2- Urca, 3- Botafogo)
The family Rhodobacteraceae is the most abundant in nearly all samples, with a particularly notable prevalence in PE3 and PP1. The genus Pleurocapsa is prominent in PE3 and PP2. Flavobacteriaceae and Sphingomonadaceae are present, but their abundance depends on the collection site and type of plastic. Bacillus,* Aquibacter*,* Truepera*,* Rhizobiaceae*,* Pseudahrensia*,* Granulosicoccus*,* Erythrobacter*, and other families are also present, but their abundances vary widely depending on the location and type of plastic polymer.
Figure 8 shows the genera of bacteria potentially pathogenic to humans (BGPH) and marine organisms (BGPMO) found in the different plastic waste samples collected from the three beaches. BGPH was detected in all samples, except in PP2, but always in smaller proportions than BGPMO, which was present in all studied communities. While Bacillus was the prevalent genus found in PE2 and PP1, Clostridium and Paracoccus (also in PP1), Psychrobacter (in PE3), and Halomonas and Vibrio (in PE2) were other predominant genera in the BGPH group (Fig. 8A).
Fig. 8. Potentially pathogenic bacteria for marine organisms (BGPMO), humans (BGPH), and abundances identified in the different plastic waste samples collected from the three studied beaches. The sample identification represents the polymer composition under analysis (PE for polyethylene and PP for polypropylene), followed by the beach from which the samples were collected (1- Leme, 2- Urca, 3- Botafogo)
Among the BGPMO, Pleurocapsa was observed in all samples, with high abundance (> 15%) in PP2 and PE3. Bacillus (PE2 and PP1), Tenacibaculum (PP1 and PE1), Halomonas and Vibrio (PE2), and Mycobacterium and Clostridium (PP1) were other notable genera in some samples (Fig. 8B).
The greatest diversity of pathogens for humans and marine organisms was found in the samples PP1 and PE2.
Discussion
Among the various threats of plastic pollution is the dispersal of pathogens and antimicrobial resistance genes through the plastisphere [33, 34]. Since the initial studies on this community, various marine environments have been investigated [3], although few have focused on tropical regions, where many areas lack adequate sanitation infrastructure and waters are contaminated by domestic effluents [4]. Two of the beaches in this study (Botafogo and Urca) are located along the waters of Guanabara Bay, where Silva et al. [4] isolated strains of Vibrio spp. and enteropathogenic Escherichia coli, while Leme, the other beach studied, is an oceanic beach near the entrance of this bay. These beaches, like others in the municipality of Rio de Janeiro, have been studied for the presence of solid waste [35, 36], but never in characterizing the microbial community associated with this waste.
The plastic cup and plastic bag samples collected from these beaches were identified as PP and PE, respectively. These two polymers are among the most collected plastics in marine environments [37, 38], likely due to their lower density compared to seawater, which allows them to float [39]. Additionally, these two types of plastic are among the most widely produced and consumed globally [40, 41].
The analysis of the plastisphere in plastic waste samples (PE and PP) revealed a diverse microbial community, with no evidence that polymer type influenced community composition. This finding corroborates the work of Zettler et al. [9], who also observed diverse bacterial communities in PE and PP samples, but differs from Amaral-Zettler et al. [42], who noted that these polymers influence the composition of the microbial communities found.
The slight differences in richness and diversity observed among the samples may be attributed to their exposure time to the ocean, as longer residence in the marine environment can directly influence community richness and diversity. Although present in all samples, the phylum Cyanobacteria was more abundant in PP2 and PE3. According to Zhai et al. [43], this phylum is primarily associated with the early stages of colonization in the plastisphere. On the other hand, the phylum Bacteroidota (especially Flavobacteriaceae) is considered a secondary colonizer [39]. Figure 6 clearly shows this succession process. In samples where Cyanobacteria predominated (PP2 and PE3), Bacteroidota abundance was lower. On the other hand, the opposite was observed in PE1 and PP3, indicating that the different samples are at distinct successional stages due to varying exposure times at their collection sites. The absence of differences in beta diversity between PP and PE suggests that their microbial communities were not shaped by plastic composition [20]. Rather, it indicates that colonization occurs during the dispersion of plastics from their source to the collection points.
The taxonomic groups found in plastic waste collected from the three beaches showed a predominance of microorganisms from the phylum Proteobacteria, commonly associated with plastic waste, surpassing their presence in seawater, glass, or organic surfaces [44], followed by Bacteroidota, with additional representation of Cyanobacteria, Planctomycetota, and Actinobacteriota. These findings align with previous studies [31, 45] showing that Proteobacteria and Bacteroidota are consistently dominant on microplastics in Brazilian and global marine environments. In Niterói beaches, also bathed by the waters of Guanabara Bay, Rhodobacteriaceae and Flavobacteriaceae were more abundant on microplastics than in the surrounding sand [31]. Similarly, research from Lagoa dos Patos, southern Brazil, reported a predominance of Proteobacteria on PP and PE samples, though with differences in other phyla, highlighting the strong environmental influence on plastisphere composition [45]. This influence is further reinforced by reviews of plastisphere studies conducted in the Northern Hemisphere [43].
Several of the dominant taxa detected are associated with metabolic capabilities relevant to plastic degradation or hydrocarbon processing. Rhodobacteraceae, a common marine family [46], occurred across all plastic types and sites and is frequently reported as a potential plastic degrader, along with other Alphaproteobacteria and Gammaproteobacteria [7, 47]. In our samples, Gammaproteobacteria—mainly Granulosicoccus—were particularly abundant in PP2 and PP3 and were also present in PE, consistent with observations from other plastisphere studies [47].
The family Sphingomonadaceae, commonly found across various polymers, including PE, PP, and polystyrene (PS) [44] and known for its ability to process hydrocarbons and produce carotenoids [48], was more abundant in PP but also present in PE. Additionally, Erythrobacter, a genus with phototrophic and aerobic capabilities and the ability to degrade PAHs [49, 50], was found across all polymers and sites. Another PAH-degrading genus, Truepera, was detected in all samples except PE3 and belongs to the phylum Deinococcota, also commonly found in plastisphere research [45, 51].
Within the family Flavobacteriaceae, the genus Aquibacter deserves attention due to its sensitivity to metal concentrations and its association with polluted environments [52]. Previously detected in metal-contaminated sediments from Sepetiba Bay, which faces issues similar to Guanabara Bay, Aquibacter’s elevated abundance in PE1 suggests that waste dispersal across areas with differing pollution levels may contribute to the microbial variation observed among sampling sites [53].
Although generally present in low abundance, several genera capable of causing disease in humans were detected in the plastisphere. Bacillus, which includes important pathogens such as B. anthracis and *B. cereus *[54], was the most notable, surpassing 5% in some samples. This low contribution differs from the results by Yang et al. [54], who reported Bacillus as one of the most abundant genera on marine microplastics and reflects the influence of factors such as location, plastic type, pollution level, and other environmental conditions on community structure. Vibrio and Clostridium, both containing clinically significant pathogenic species, contributed with ≤ 1% in abundance. Despite their low representation, their persistence on plastic surfaces raises public health concerns, especially considering previous reports of Vibrio spp. on microplastics in Guanabara Bay [4].
Multiple taxa with pathogenic potential to marine organisms were also identified. Pleurocapsa, known to cause disease in algae [55], was the most prominent, exceeding 20% in PE3 and PP2. Flavobacteriaceae, a family associated with coral disease and with disease states in sponges, starfish, macroalgae, and lobsters [56], occurred in nearly all samples and may additionally contribute to the spread of antibiotic-resistance genes [57, 58]. Rhodobacteraceae and Rhizobiaceae—both linked to coral diseases [48]—were also present in all samples, indicating that plastic waste may act as a vector for taxa harmful to coral ecosystems. While no corals occur in the study area, the spread of plastic waste could affect nearby coral-rich regions such as Arraial do Cabo and Angra dos Reis. Other genera of concern include Bacillus, which contains species that cause infections in fish and invertebrates [54, 59], and Halomonas and Tenacibaculum, each exceeding 1% in some samples (PE2 and PP1, respectively).
Planctomycetota, Actinobacteriota, Chloroflexi, and predatory taxa such as Bdellovibrionota and Myxococcota, were other phyla documented in this study, and all have been reported in plastisphere research [39].
Biofilm formation on plastics depends on factors such as polymer type, shape, exposure time, and environmental conditions, with polymer type being particularly well studied [54]. Carson et al. [60] reported higher bacterial abundance on PS compared to PE and PP, likely due to surface differences. Similarly, Zettler et al. [9] found distinct microbial communities on PP and PE, sharing less than 50% similarity. These authors reported that some microorganisms were more abundant on PP and high-density polyethylene (HDPE) surfaces than in the surrounding water and that the microbial communities on these plastics differed markedly from those in the ambient environment.
Our findings indicate that variations in microbial abundance across samples are more closely linked to the length of time the plastics remained in the marine environment. Although communities on different polymers formed separate clusters, no clear polymer-specific associations were observed, suggesting that exposure duration plays a stronger role in shaping microbial colonization. The presence of degraded plastics further supports prolonged environmental exposure and underscores the influence of durability on plastic pollution.
Conclusion
The results of this study align with other research on the plastisphere [9, 15, 39, 44, 49], indicating a consistent presence of various microorganisms adhering to plastic surfaces. The bacteria identified in the samples are cited in the literature of other studies involving plastic and microplastic waste. Some observed genera include species involved in degradation and potential pathogens for animals and humans.
Although the samples analyzed showed no influence of the polymer composition on bacterial community formation, the exposure time of the waste in the sea appears to have a greater effect.
The gaps in our understanding create opportunities for future research to explore various aspects, such as the effects of microorganisms associated with plastics on marine life, the potential transfer of antimicrobial resistance, and the impacts on human health. Comparative analyses with the marine water microbial community can provide valuable context, helping to discern the unique attributes of the plastisphere.
Future studies investigating the genetic diversity and functional capabilities of microorganisms associated with plastic waste could provide insights into their roles in degradation and potential pathogenicity. Additionally, isolating and testing pathogenic bacteria and antimicrobial resistance genes would contribute important data on the health risks posed by the plastisphere.
By revealing the ecological consequences of plastic pollution and its potential implications for human health, our research contributes to the growing body of evidence highlighting the urgent need for effective mitigation strategies. This understanding benefits the local population and has global significance in addressing the widespread issue of marine pollution.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Our World in Data (2024) Plastic Pollution. Retrieved from https://ourworldindata.org/plastic-pollution. Accessed 07 July 2024
- 2INEA, Instituto Estadual do Ambiente do Rio de Janeiro (2024) https://www.inea.rj.gov.br/rio-de-janeiro/. Accessed 31 March 2022
- 3Kuczynski J, Stombaugh J, Walters WA, González A, Caporaso JG, Knight R (2012) Using QIIME to analyze 16S r RNA gene sequences from microbial communities. Curr Protoc Microbiol. Chapter 1:Unit 1E.5. 10.1002/9780471729259.mc 01e 05s 27. PMID: 23184592; PMCID: PMC 447784310.1002/9780471729259.mc 01e 05s 27PMC 447784323184592 · doi ↗ · pubmed ↗
- 4Oberbeckmann S, Labrenz M (2019) Marine microbial assemblages on microplastics: Diversity, Adaptation, and role in degradation. Ann Rev Mar Sci 12(1). 10.1146/annurev-marine-010419-01063310.1146/annurev-marine-010419-01063331226027 · doi ↗ · pubmed ↗
- 5Egan S, Fernandes ND, Kumar V, Gardiner M, Thomas T (2014) Bacterial pathogens, virulence mechanism and host defence in marine macroalgae. Environ Microbiol 16(4), 925–938. 10.1111/1462-2920.1228856 Hudson 10.1111/1462-2920.1228824112830 · doi ↗ · pubmed ↗