Identification and characterization of VC1123, a novel gene required for colonization in Vibrio cholerae
Xiuping Fu, Xinyu Wen, Yuanyuan Yang, Weili Liang, Bo Pang, Xu Li, Baowei Diao, Jie Li, Jingyun Zhang, Biao Kan

TL;DR
A new gene, VC1123, was found to reduce Vibrio cholerae's ability to colonize the gut by regulating key bacterial proteins.
Contribution
Identification of VC1123 as a novel negative regulator of Vibrio cholerae intestinal colonization through its effect on outer membrane porin genes.
Findings
Deletion of VC1123 significantly enhances Vibrio cholerae colonization in neonatal mice.
VC1123 regulates ompU and ompT, which are linked to bile resistance and gut colonization.
VC1123 operates independently of the ToxR/ToxS regulatory system.
Abstract
A novel lineage of serogroup O1 El Tor Vibrio cholerae, genetically distinct from the seventh-pandemic strain, has recently emerged in China and has been associated with diarrheal outbreaks. However, the molecular basis underlying its virulence potential remains incompletely understood. This study aimed to identify virulence-associated determinants in the new lineage through comparative transcriptomic and functional analyses. We compared the transcriptomes of two new-lineage isolates—VC6050 (ctxAB+) and VC6055 (ctxAB−)—with the representative seventh-pandemic strain N16961 (ctxAB+) following growth in virulence-inducing (AKI) and non-inducing (LB) media. Differential expression patterns were analyzed, with particular focus on the toxigenic new-lineage strain VC6050 and N16961 under AKI conditions. A highly expressed gene shared across all three strains, VC1123, was selected for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
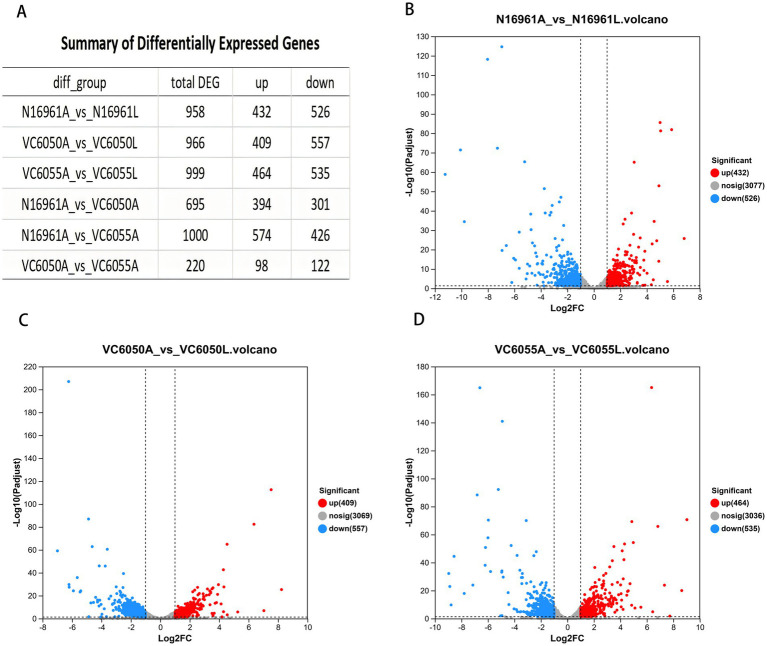 Figure 1
Figure 1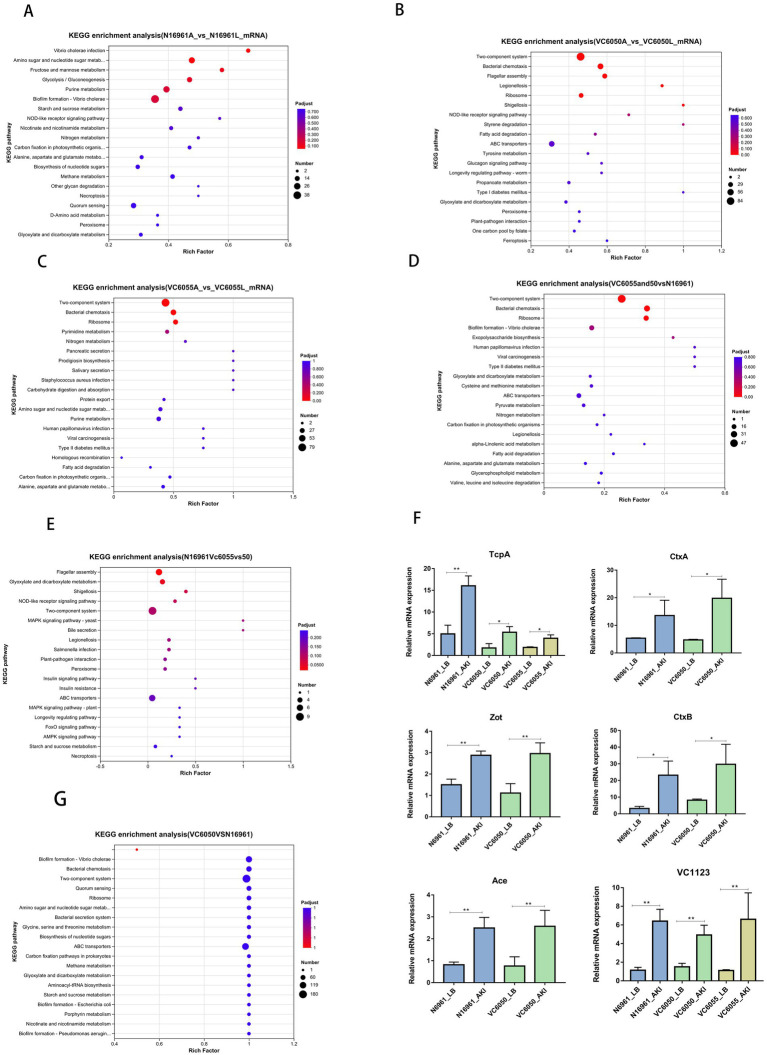 Figure 2
Figure 2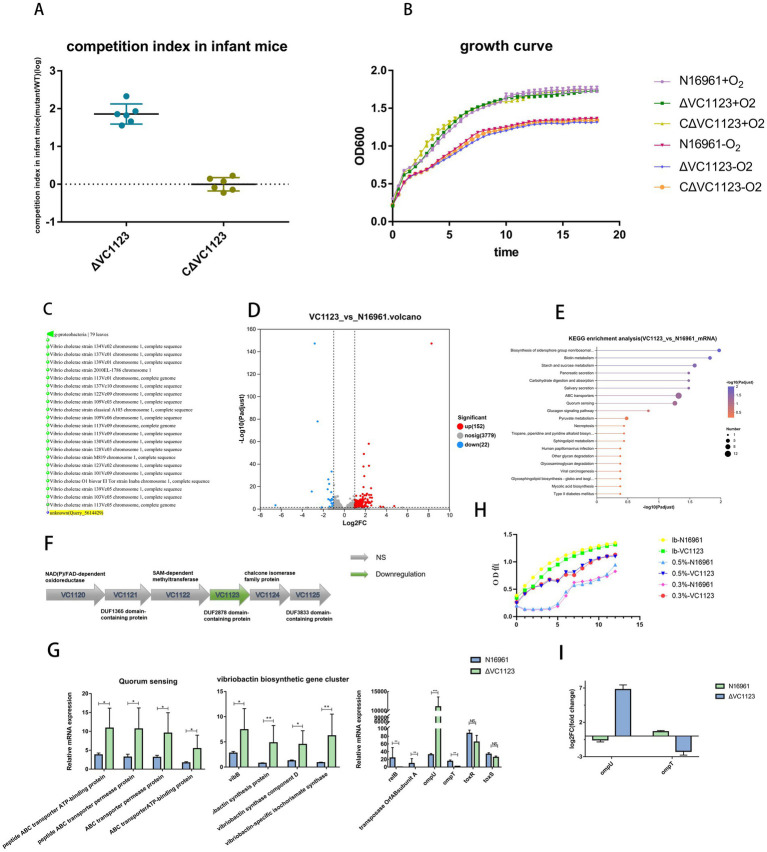 Figure 3
Figure 3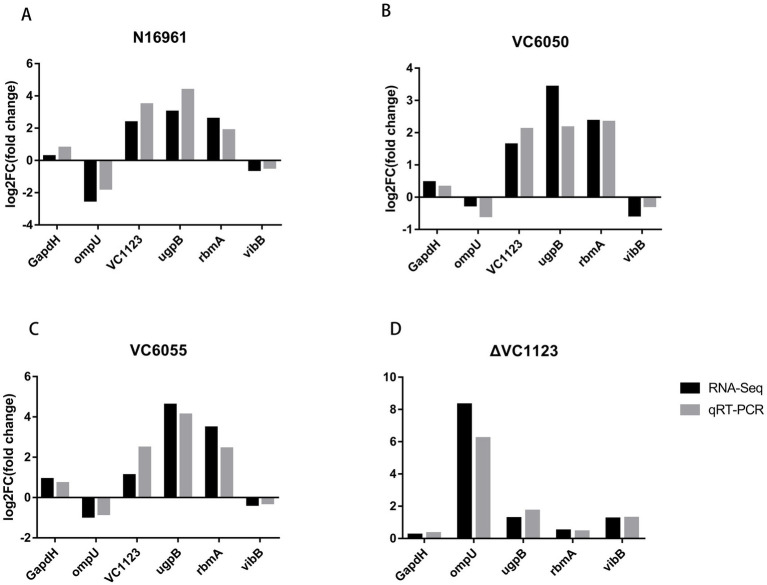 Figure 4
Figure 4- —National Key Research and Development Program of China10.13039/501100012166
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Listeria monocytogenes in Food Safety · Escherichia coli research studies
Introduction
Cholera is a severe intestinal infectious disease caused by Vibrio cholerae. According to the World Health Organization, over 560,000 cases and more than 6,000 deaths were reported globally across 60 countries in 2024 (World Health Organization, 2025). V. cholerae is a Gram-negative bacterium that naturally resides in aquatic environments and associates with plankton. Infection occurs via the oral route, followed by colonization of the small intestinal epithelium. Intestinal colonization depends on the toxin-co-regulated pilus (TCP), which also serves as the receptor for the CTXφ bacteriophage (Gutierrez-Rodarte et al., 2019).
V. cholerae produces cholera toxin (CT), an ADP-ribosylating toxin that elevates intracellular cyclic AMP (cAMP) levels in host enterocytes, leading to massive secretory diarrhea and dehydration (Lebens and Holmgren, 1994). The genes encoding CT (ctxA and ctxB) are located on the lysogenic CTXφ bacteriophage. The core region of CTXφ includes ctxA, ctxB, zot, cep, ace, and orfU, and can be horizontally transferred between toxigenic and non-toxigenic strains (Waldor and Mekalanos, 1996).
Expression of the major virulence factors TCP and CT is coordinately regulated by the ToxR/ToxS transcriptional activator system (Miller et al., 1989; Herrington et al., 1988). ToxR, the first identified virulence gene regulator in V. cholerae, forms a complex with TcpP to activate toxT, which encodes a cytoplasmic transcriptional activator that directly induces expression of ctxAB, tcp, and accessory colonization factor genes (Miller and Mekalanos, 1985; Kovacikova and Skorupski, 1999; Haas et al., 2015; Richard et al., 2010). Independently of TcpP and ToxT, ToxR also modulates the expression of two outer membrane porins: it activates ompU transcription and represses ompT (Chakrabarti et al., 1996; Champion et al., 1997; Li et al., 2000; Crawford et al., 1998). Both OmpU and OmpT have been implicated in intestinal colonization (Provenzano and Klose, 2000).
This ToxR-dependent virulence cascade is robustly induced in the host intestinal environment, whereas in vitro induction requires specific, artificial culture conditions (Ross, 1996). To mimic key physicochemical cues of the small intestine—such as low oxygen tension, bicarbonate, pH, and osmolarity—the AKI culture system was established in 1986 (Iwanaga et al., 1986). Under AKI conditions, the V. cholerae virulence regulatory network is effectively activated, making this system a widely used model for studying virulence gene expression and regulation.
Although more than 200 V. cholerae serogroups have been identified, only toxigenic strains of serogroups O1 and O139 have caused epidemic cholera (Albert et al., 1993; Devault et al., 2014). Recently, a novel lineage of O1 El Tor V. cholerae, genetically distinct from the seventh pandemic strain, was identified in China and associated with diarrheal outbreaks (Yan et al., 2022). Our prior work showed that this new lineage induces significantly lower levels of inflammatory cytokines and chemokines compared to the classical pandemic strain (Fu et al., 2025). The new lineage contains toxigenic strains but is genetically distinct from the seventh pandemic strain. It remains unclear whether its virulence expression resembles that of the pandemic strains.
In this study, the new lineage strains VC6050 (ctxAB^+^) and VC6055 (ctxAB^−^), as well as the representative seventh pandemic strain N16961 (ctxAB^+^), were selected and cultured in AKI medium or LB medium. Then, their transcriptomes were compared to identify similarities and differences in virulence—factor expression between the new lineage and the pandemic strains, and to determine potential virulence factors. In addition, we screened candidate genes for functional validation to pinpoint potential virulence factors involved in V. cholerae pathogenesis.
Materials and methods
Bacterial strains
The following strains were used in this study: the seventh-pandemic strain N16961 (serogroup O1, El Tor biotype, ctxAB^+^); its isogenic deletion mutants N16961-ΔVC1123, the complemented strain N16961-ΔVC1123/cVC1123; and two isolates from the newly identified lineage, VC6050 (serogroup O1, ctxAB^+^) and VC6055 (serogroup O1, ctxAB^−^).
Media and culture conditions
For in vitro growth experiments, strains N16961, VC6050, and VC6055 were cultured in either AKI medium (1.5% Bacto peptone, 0.4% yeast extract, 0.5% NaCl, 0.4% NaHCO₃, pH 7.4) or standard LB broth (1% tryptone, 0.5% yeast extract). Cultures were prepared in biological triplicate for each condition. In the AKI induction protocol, bacteria were first grown statically at 37 °C for 4 h to reach an OD_600_ of ~1.0, followed by continued incubation under shaking conditions (200 rpm, 37 °C) for an additional 6 h to induce virulence gene expression.
Growth curve analysis
To assess whether deletion of VC1123 affects the growth kinetics of V. cholerae, strains N16961 (wild type), N16961-ΔVC1123, and the complemented strain N16961-ΔVC1123/cVC1123 were pre-cultured in LB broth at 37 °C with shaking (200 rpm) for 5 h to reach an OD_600_ of ~1.0. These cultures were then diluted 1:100 into fresh LB broth and grown in triplicate under both aerobic and anaerobic conditions at 37 °C shaking at 200 rpm. OD_600_ was measured hourly using a spectrophotometer. Growth curves represent the mean ± standard deviation of three biological replicates.
Total RNA extraction and RNA sequencing
V. cholerae strains N16961, VC6050, and VC6055 were cultured in either AKI medium or standard LB broth until they reached an OD_600_ of approximately 1.0. V. cholerae strains N16961 and N16961-ΔVC1123 were cultured in LB medium until they reached an OD_600_ of approximately 1.0.
Then, total RNA was extracted from bacterial cultures grown in AKI and LB media using TRIzol^®^ Reagent (Thermo Fisher Scientific), and genomic DNA was removed according to our previous experimental procedures (Fu et al., 2014).
All experiments were conducted with three independent biological replicates per sample. An rRNA Depletion Kit for Mixed Bacterial Samples (lexogen, United States) was used. Then, all mRNAs were first broken into short (200 nt) fragments by adding fragmentation buffer. Secondly, double-stranded cDNA was synthesized with random hexamer primers (Illumina). Then, the synthesized cDNA was subjected to end—repair, phosphorylation, and ‘A’ base addition according to Illumina’s library construction protocol. An RNA—seq transcriptome library was prepared following Illumina^®^ Stranded mRNA Prep, Ligation (San Diego, CA) using total RNA. A paired—end RNA—seq library was sequenced with the Illumina Novaseq Xplus (Illumina Inc., San Diego, CA, United States).
The original images were processed into sequences, and base—calling and quality value calculations were performed. Clean reads were obtained by removing low-quality sequences, reads with more than 10% of N bases (unknown bases), and reads containing adaptor sequences.
Differential expression analysis and functional enrichment
Transcript abundance was quantified as transcripts per million (TPM), and gene counts were estimated using RSEM. Differentially expressed genes (DEGs) between conditions were identified using DESeq2 implemented in R (v4.3.0) (Love et al., 2014).
Functional enrichment analysis was performed to identify Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways among the DEGs. KOBAS 2.01 is used to identify statistically significantly enriched pathway using Fisher’s exact test. Enrichment significance was assessed against the whole-transcriptome background using FDR-corrected p-values, with a threshold of p ≤ 0.05.
Reverse transcription quantitative real-time PCR (RT-qPCR)
To validate RNA-seq results, six DEGs were selected for RT-qPCR analysis. Quantitative PCR was performed according to our previous experimental procedures (Fu et al., 2025). Gene expression levels were normalized to the housekeeping gene gap (encoding glyceraldehyde-3-phosphate dehydrogenase) and calculated using the 2^−ΔΔCq^ method. Each qPCR reaction was run in technical triplicate, and the entire experiment was independently repeated twice. Primer sequences are listed in Supplementary Table 1.
To further verify whether the expression of ompU and ompT also differed between the wild-type strain and the VC1123 mutant under AKI conditions, rather than only in LB medium, V. cholerae strains N16961 and N16961-ΔVC1123 were cultured in AKI medium for 4 h to reach an OD_600_ of ~1.0. Total RNA was then extracted, and RT-qPCR analysis was performed. Primers for ompU are listed in Supplementary Table S1, while primers for ompT were as follows: Forward primer: 5′-CGCCAGTGTTCGCTTCTTG-3′ and Reverse primer: 5′-AGGTCTTCGTTACGGCACACTG-3′.
PCR amplification was carried out with an initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 5 s and annealing at 60 °C (or the corresponding primer Tm) for 30 s.
Construction of VC1123 deletion and complementation strains
The nucleotide sequence of the V. cholerae VC1123 gene was retrieved from the NCBI RefSeq database. Homologous sequences were identified using BLASTN with an E-value threshold of 1 × 10^−5^.
A clean, in-frame deletion mutant of VC1123 was constructed in the parental strain N16961 via allelic exchange using the suicide vector pWM91. Briefly, DNA fragments corresponding to the upstream (amplified with primers VC1123F1/VC1123R1) and downstream (amplified with primers VC1123F2/VC1123R2) regions of VC1123 were PCR-amplified from N16961 genomic DNA. These fragments were assembled into XhoI/SacI-digested pWM91 using the ClonExpress^®^ Ultra One Step Cloning Kit V3 (Vazyme), yielding the allelic exchange plasmid pWM91-ΔVC1123. The plasmid pWM91-ΔVC1123 was then transformed into the Escherichia coli strain SM10 pir and subsequently transferred to the strain N16961 by conjugation. The transconjugants were selected on LB agar plates containing streptomycin (100 μg/mL) and ampicillin (100 μg/mL). The clones were streaked on LB agar plates with 8% sucrose and without NaCl at 22 °C. After 48 h, double—crossover recombination mutants were selected and verified by sequencing to generate the mutant strain, N16961-ΔVC1123.
For genetic complementation, the full-length VC1123 open reading frame (ORF) was amplified from N16961 DNA using primers H1123F and H1123R. The amplicon was cloned into the SpeI/NdeI-digested vector pSRKKtc using the same ClonExpress^®^ cloning kit, generating the plasmid pSRKKtc-VC1123. The construct was confirmed by restriction digestion and sequencing. The plasmid was then electroporated into N16961-ΔVC1123, producing the complemented strain N16961-ΔVC1123/pSRKKtc-VC1123 (abbreviated as N16961-ΔVC1123/cVC1123).
All primers used for mutant and complementation construct generation are listed in Supplementary Table S2.
Infant mouse colonization competition assays
Strains N16961, N16961-ΔVC1123, and the complemented strain N16961-ΔVC1123/cVC1123 were grown overnight on LB agar plates at 37 °C. Cells were harvested and resuspended in phosphate-buffered saline (PBS) to an OD_600_ of 1.0 (~1 × 10^9^ CFU/mL). For each competition assay, ~1 × 10^5^ CFU of either N16961-ΔVC1123 or N16961-ΔVC1123/cVC1123 was mixed with an equal number of wild-type N16961 cells in a total volume of 50 μL (1:1 input ratio). The actual the input ratios (mutant to wild type) of the inoculum was verified by serial dilution and plating on LB agar.
Five- to six-day-old C57BL/6 neonatal mice, separated from their dams for 2–3 h prior to infection, were intragastrically inoculated with 50 μL of the bacterial mixture. At 24 h post-inoculation, pups were euthanized, and the entire small intestine (SI) from each mouse was aseptically removed and placed individually into 5 mL of cold PBS. Each intestinal sample was homogenized separately. Serial dilutions were made and plated on LB agar plates supplemented with streptomycin for calculation of the output ratios (mutant to wild type). Genomic DNA was extracted directly from 10-fold diluted homogenates (see below) for digital PCR (dPCR)-based quantification. Each experimental group included five mice, and two independent biological replicates were performed. The competitive index (CI) was calculated for each mouse as:
A CI < 1 indicates impaired colonization fitness of the mutant relative to the wild type. All assays were repeated in at least two independent experiments.
Digital PCR (dPCR) quantification of bacterial loads
Intestinal colonization levels were quantified using dPCR on a QIAcuity One system (Qiagen, Hilden, Germany). Primers (Supplementary Table S2) were designed to target two genomic loci: (i) a unique sequence within the deleted region of VC1123 (absent in the ΔVC1123 mutant but present in wild-type and complemented strains), and (ii) the single-copy housekeeping gene recA, which is conserved in all strains and serves as a proxy for total V. cholerae genome equivalents. Because both targets are present in one copy per chromosome, dPCR-derived copy numbers directly reflect bacterial genome counts. The recA signal represents the total bacterial population (wild type + mutant), whereas amplification of the VC1123-specific locus specifically quantifies wild-type (or complemented) cells. The mutant population was inferred by subtracting the wild-type count from the total (recA) count.
Each 40 μL dPCR reaction contained 13.3 μL of 3 × QIAcuity EG Master Mix, 0.4 μM of each primer, and 4 μL of template DNA. Reactions were loaded onto QIAcuity Nanoplate 26 k chips, sealed, and thermocycled under the following conditions: 95 °C for 2 min; 40 cycles of 95 °C for 15 s, 58 °C for 15 s, and 72 °C for 15 s; followed by a final hold at 40 °C for 5 min. Fluorescence data were acquired and analyzed using QIAcuity Software Suite v2.0 to determine target copies per microliter.
Template DNA was prepared from intestinal homogenates as follows: small intestines were homogenized in 5 mL PBS, diluted 1:10 in PBS, boiled for 10 min, and centrifuged at 12,000 × g for 2 min; the supernatant was used directly as PCR template. Meanwhile, to evaluate the potential influence of DNA from nonviable bacteria on the digital PCR (dPCR) assay, single colonies recovered from plates following serial dilution were analyzed by dPCR to identify their genotypes (wild-type, deletion mutant, or complemented strain). The results demonstrated that the competitive index (CI) determined from single-colony analysis were not significantly different from intestinal homogenates obtained by dPCR. Accordingly, the subsequent presentation and discussion of results are based directly on the dPCR data. All dPCR reactions were performed in technical triplicate. Primer sequences are listed in Supplementary Table S3.
Bile salt tolerance assay
To assess whether deletion of VC1123 increased bile salt tolerance, V. cholerae N16961 and N16961-ΔVC1123 were pre-cultured in LB broth at 37 °C with shaking (200 rpm) for 5 h to reach an OD_600_ of ~1.0. The cultures were subsequently diluted 1:100 into fresh LB medium supplemented with 0.3% or 0.5% bile salts, with LB medium without bile salts used as the control. Cultures were incubated at 37 °C with shaking at 200 rpm for 12 h. OD_600_ was measured hourly using a spectrophotometer. Growth curves represent the mean ± standard deviation of three biological replicates. All experiments were performed with three independent biological replicates per sample.
Results
Transcriptomic profiling of Vibrio cholerae strains under virulence-inducing (AKI) and non-inducing (LB) conditions
To investigate the transcriptional responses of pandemic and emerging V. cholerae lineages to virulence-inducing conditions, we performed RNA sequencing on the seventh-pandemic strain N16961 and two isolates from the newly identified lineage —VC6050 (ctxAB^+^) and VC6055 (ctxAB^−^)—cultured in AKI medium (virulence-inducing) and standard LB broth (control). A total of 18 libraries (3 strains × 2 conditions × 3 biological replicates) yielded 67.97 Gb of high-quality sequencing data, with each sample generating >2.94 Gb of clean data and a Q30 base percentage exceeding 96.56%.
Global transcriptomic responses to AKI induction
Relative to LB-grown controls, AKI culture induced extensive changes in gene expression in all three strains: 958, 966, and 999 DEGs were identified in N16961, VC6050, and VC6055, respectively (Figure 1).
Results of transcriptome analysis. (A) Summary of differentially expressed genes. (B–D) Volcano plot of differentially expressed genes (DEGs) for groups N16961, VC6050, and VC6055 in AKI compared to the control group (LB), respectively.
In N16961, DEGs were significantly enriched in pathways including Vibrio cholerae infection, amino sugar and nucleotide sugar metabolism, fructose and mannose metabolism, glycolysis/gluconeogenesis, and biofilm formation (Figure 2A). In strain VC6050, DEGs were mainly enriched in the two-component system, bacterial chemotaxis, ribosome, flagellar assembly, and ABC transporters (Figure 2B). For strain VC6055, DEG enrichment was observed predominantly in the two-component system, bacterial chemotaxis, ribosome, pyrimidine metabolism, and nitrogen metabolism (Figure 2C).
KEGG enrichment analysis results. (A–C) The top 20 pathways ranked by KEGG enrichment analysis for groups N16961, VC6050, and VC6055 compared to the control group (LB), respectively. (D) The top 20 pathways ranked by KEGG enrichment analysis for N16961 versus VC6050/VC6055. (E) The top 20 pathways ranked by KEGG enrichment analysis for N16961/VC6050 versus VC6055. (F) The expression of genes, with particular attention given to (tcpA, ctxAB, zot, ace, and VC1123) [n = 3, mean ± standard deviation (SD)]. * p < 0.01, ** p < 0.001; unpaired two-tailed t-test. (G) The top 20 pathways ranked by KEGG enrichment analysis for VC6050 versus N16961.
Notably, bacterial chemotaxis and biofilm formation pathways were consistently downregulated across all three strains under AKI conditions. Key chemotaxis genes—including cheW, cheR, cheB, cheD, mcpH, and cher2—showed pronounced transcriptional repression (Figure 2F). Conversely, canonical virulence determinants were upregulated: tcpA was induced in all strains, while ctxAB (cholera toxin), zot (zonula occludens toxin), and ace (accessory cholera enterotoxin) were specifically upregulated in the toxigenic strains N16961 and VC6050 (Figure 2F).
Comparative analysis among strains under AKI conditions
Pairwise comparisons of the three strains in AKI medium revealed extensive transcriptional divergence: N16961 vs. VC6050, 695 DEGs; N16961 vs. VC6055, 1,000 DEGs; VC6050 vs. VC6055, 220 DEGs (Figure 1). Relative to the N16961 AKI group, the AKI-cultured new-lineage strains VC6050 and VC6055 shared 342 DEGs, which were significantly enriched in the two-component system, bacterial chemotaxis, and biofilm formation (Figure 2D). When comparing only the toxigenic strains (N16961 and VC6050) against the non-toxigenic VC6055, 84 DEGs were shared and enriched in flagellar assembly and bile secretion pathways (Figure 2E).
Focusing specifically on the comparison between VC6050 and N16961 under AKI conditions, we identified 301 upregulated and 394 downregulated genes in VC6050. These DEGs were predominantly associated with bacterial chemotaxis, biofilm formation, and two-component signaling. Importantly, the magnitude of downregulation in chemotaxis- and biofilm-related genes was greater in VC6050 than in N16961 (Figure 2G).
Deletion of VC1123 enhances intestinal colonization by Vibrio cholerae in infant mice
The gene VC1123, which was highly expressed across all three strains—N16961, VC6050, and VC6055 (Figure 2F)—was selected for functional characterization based on several considerations beyond transcriptional abundance alone. VC1123 is a 540-bp open reading frame in V. cholerae that encodes a protein containing a domain of unknown function (DUF), making it an attractive candidate for uncovering previously unrecognized regulatory mechanisms. In addition, VC1123 is highly conserved among V. cholerae isolates, as evidenced by multiple-sequence alignment showing highly conserved amino acid sequences in O1 and O139 serogroups as well as diverse non-O1/non-O139 environmental strains (Figure 3C), suggesting an important biological role. Moreover, its consistently high expression in both the new-lineage isolates and the seventh-pandemic reference strain under virulence-inducing conditions indicates that VC1123 may participate in core processes related to host adaptation rather than strain-specific traits. Collectively, these features distinguished VC1123 from other AKI-induced genes and provided the rationale for selecting it for further functional investigation.
Deletion of the VC1123 gene enhances the colonization ability of V. cholerae in the infant mouse intestine. (A) Colonization competition assays in infant mice. (B) Growth curve analysis of the V. cholerae strain N16961, N16961-ΔVC1123, and N16961-ΔVC1123/cVC1123 in aerobic or anaerobic conditions. (C) Homologous sequences were identified using BLASTN of the V. cholerae VC1123 gene with an E-value threshold of 1e-5. (D) Volcano plot of differentially expressed genes (DEGs) for N16961-ΔVC1123 versus N16961. (E) The top 20 pathways ranked by KEGG enrichment analysis for N16961-ΔVC1123 versus N16961. (F) The transcription of VC1123 neighboring genes (VC1120–VC1125). (G) The expression of ompU, ompT, toxR, toxS and several genes related to iron uptake and the quorum sensing system [n = 3, mean ± standard deviation (SD)] * p < 0.01, ** p < 0.001, *** p < 0.0001, **** p < 0.00001; unpaired two-tailed t-test. (H) Bile salt tolerance assay. V. cholerae N16961 and N16961-ΔVC1123 were incubated in LB medium supplemented with 0.3% or 0.5% bile salts, with LB medium without bile salts used as the control. (I) The relative transcript levels of ompU and ompT in groups of N16961 and the VC1123 mutant under AKI conditions as determined by RT-qPCR.
Efforts to construct VC1123 deletion mutants in the new-lineage strains VC6050 and VC6055 were unsuccessful, as no transconjugants could be recovered on dual-antibiotic selection plates following conjugation. During the deletion of other genes in the new-lineage strains, the suicide plasmid similarly failed to be transferred into the recipient cells; the same phenomenon was also observed in our previous gene deletion experiments using environmental isolates. Consequently, only the VC1123 deletion mutant in the N16961 background (N16961-ΔVC1123) was obtained. A complemented strain, N16961-ΔVC1123/cVC1123, was also generated.
To assess the role of VC1123 in intestinal colonization, competitive infection assays were performed in 5–6-day-old C57BL/6 neonatal mice. Equal mixtures (1:1 ratio) of wild-type N16961 with either N16961-ΔVC1123 or N16961-ΔVC1123/cVC1123 were administered intragastrically. Consistently, 24 h post-inoculation, the ΔVC1123 mutant exhibited an 86.34-fold increase in intestinal colonization relative to the wild-type strain (Figure 3A). In contrast, the complemented strain showed colonization levels comparable to wild-type, with a mean CI of 1.06 (Figure 3A), indicating that VC1123 negatively regulates intestinal colonization.
To rule out growth defects as a confounding factor, we compared the in vitro growth kinetics of the mutant and wild-type strains under both aerobic and anaerobic conditions. No significant differences in growth rate were observed (Figure 3B), indicating that the enhanced colonization was not due to altered bacterial fitness in broth culture.
To explore the molecular basis of this phenotype, we performed RNA sequencing on N16961 and N16961-ΔVC1123 cultured in LB medium to mid-log phase (5 h). Deletion of VC1123 did not affect transcription of neighboring genes (VC1120–VC1125; Figure 3F), suggesting minimal polar effects on the surrounding genes. Differential expression analysis identified 174 DEGs in the mutant relative to wild-type, of which 152 were upregulated and 22 were downregulated (Figure 3D). KEGG pathway enrichment revealed that these DEGs were primarily associated with quorum sensing, biosynthesis of siderophore group nonribosomal peptides, and biotin metabolism (Figure 3E).
Notably, the outer membrane protein gene ompU was dramatically upregulated (315.6-fold) in the ΔVC1123 strain, while ompT was downregulated (8.47-fold). In contrast, the virulence-associated regulators toxR and toxS showed no significant changes in expression. Additionally, relB and the transposase orfAB subunit A were strongly downregulated (90.9-fold and 66.67-fold, respectively).
Genes involved in the quorum sensing, including ABC transporter permease protein, ABC transporter ATP-binding protein, peptide ABC transporter permease protein, and peptide ABC transporter ATP-binding protein, were significantly upregulated, with an average increase of 2.74-fold. Furthermore, multiple genes critical for iron acquisition via vibriobactin biosynthesis were significantly induced, including vibriobactin-specific 2,3-dihydroxybenzoate-AMP ligase, vibriobactin synthesis protein, vibriobactin synthase component D, and vibriobactin-specific isochorismate synthase (Figure 3G).
To evaluate whether deletion of VC1123 increased bile salt tolerance, V. cholerae N16961 and N16961-ΔVC1123 were cultured in LB medium supplemented with different concentrations of bile salts, and bacterial growth was compared under the indicated conditions. The results showed that, compared with the N16961 strain, N16961-ΔVC1123 exhibited a pronounced growth advantage at bile salt concentrations of 0.3 and 0.5% (Figure 3H).
To further verify whether the expression of ompU and ompT also differed between the wild-type strain and the VC1123 mutant under AKI conditions, rather than only in LB medium, RT-qPCR analysis was performed. The results showed that, under AKI culture conditions, ompU was significantly upregulated, whereas ompT was significantly downregulated in the VC1123 deletion mutant compared with the wild-type strain (Figure 3I), consistent with the observations in LB medium.
To validate the RNA-seq findings, six DEGs were selected for RT-qPCR analysis. The expression trends observed by RT-qPCR were fully consistent with those from transcriptomic data (Figure 4), confirming the reliability of the sequencing results.
RT-qPCR validation of transcriptome data. (A–C) The relative transcript levels of six DEGs were consistent between RNA-seq and RT-qPCR in groups of N16961, VC6050, and VC6055 in AKI medium. (D) The relative transcript levels of five DEGs were consistent between RNA-seq and RT-qPCR in ΔVC1123. Black bars represent RNA-seq results, while gray bars represent RT-qPCR results.
Discussion
In this study, we employed RNA-seq to characterize the transcriptional responses of pandemic and emerging V. cholerae lineages under virulence-inducing (AKI) versus non-inducing (LB) conditions. Consistent with the established role of AKI medium in mimicking the host intestinal environment, all three strains—N16961 (ctxAB^+^), VC6050 (ctxAB^+^), and VC6055 (ctxAB^−^)—exhibited strong upregulation of the major colonization factor tcpA. Moreover, the canonical virulence genes ctxAB, zot, and ace were specifically induced in the two toxigenic strains (N16961 and VC6050), confirming that AKI conditions effectively activate the core virulence regulon.
Notably, genes associated with bacterial chemotaxis and biofilm formation were consistently downregulated across all three strains in AKI medium. This transcriptional shift suggests a global reallocation of cellular resources away from environmental persistence mechanisms toward host colonization programs—a strategy previously linked to reduced intracellular cyclic diguanylate (c-di-GMP) levels (Matson et al., 2007). In V. cholerae, the phosphodiesterase VieA lowers c-di-GMP, thereby repressing vps-mediated biofilm formation and promoting expression of colonization factors such as TCP (Tischler and Camilli, 2004). Our data align with this model, reinforcing the notion that low c-di-GMP states favor virulence over environmental adaptation.
Intriguingly, the toxigenic new-lineage strain VC6050 displayed more pronounced repression of chemotaxis- and biofilm-related genes than the classical pandemic strain N16961. We hypothesize that this enhanced downregulation may reflect an evolved strategy to minimize host immune detection or phage predation. For instance, reduced flagellar synthesis and biofilm production could lower immunogenicity and decrease susceptibility to phage recognition. Supporting this idea, our prior work demonstrated that these new-lineage isolates exhibit reduced sensitivity to the VP3 bacteriophage (Yuanxi et al., 2025), suggesting potential co-evolutionary adaptations favoring stealth during infection.
Building on the transcriptomic profiling, we identified VC1123—a gene highly expressed in all three strains—as a candidate modulator of virulence. VC1123 encodes a conserved protein containing a DUF2878 (PF11086) domain and is annotated within the PhrR regulon (RegPrecise), implying regulation by the MerR-family transcription factor PhrR. Using N16961 as the parental strain, we generated an in-frame deletion mutant (N16961-ΔVC1123) and a complemented derivative. Competitive colonization assays in neonatal mice revealed that loss of VC1123 conferred an enhancement in intestinal colonization, without altering in vitro growth under aerobic or anaerobic conditions. Furthermore, transcription of flanking genes (VC1120–VC1125) remained unchanged, ruling out polar effects. These findings collectively indicate that VC1123 functions as a negative regulator of colonization.
RNA-seq analysis of the ΔVC1123 mutant uncovered significant dysregulation of outer membrane porin genes: ompU was dramatically upregulated (315.6-fold), while ompT was downregulated (8.47-fold), despite no detectable changes in toxR or toxS expression. This observation is particularly significant given the well-established roles of OmpU and OmpT in V. cholerae pathogenesis (Valeru et al., 2012). OmpU enhances resistance to bile salts and cationic antimicrobial peptides, promotes adhesion to intestinal epithelial cells, and facilitates colonization in murine models (Provenzano and Klose, 2000; Duret and Delcour, 2006; Wibbenmeyer et al., 2002; Mathur and Waldor, 2004; Sperandio et al., 1995). In contrast, OmpT compromises bile tolerance and reduces fitness in the gut (Wibbenmeyer et al., 2002). Notably, anti-OmpU antibodies inhibit bacterial attachment in vitro and diminish colonization in vivo (Sperandio et al., 1995), and OmpU can also trigger IL-8 secretion from intestinal epithelial cells, potentially shaping host inflammatory responses (Yang et al., 2018). Moreover, OmpU has been implicated as a receptor for epithelial cell endocytosis and as a scaffold for cholera toxin packaging into outer membrane vesicles (OMVs), thereby stabilizing toxin activity in the intestine (Zingl et al., 2021).
Given that toxR/toxS expression was unaffected, our data suggest that VC1123 modulates ompU/ompT through a ToxR-independent pathway. Potential mechanisms include direct or indirect regulation via alternative transcription factors, post-transcriptional control, or modulation by host- or microbiota-derived signals. For example, gut microbial metabolites have been shown to influence ompU expression and colonization efficiency (Qin et al., 2020). Additionally, the observed downregulation of relB—a component of the RelE/RelB toxin-antitoxin system—may contribute to the phenotype, as TA systems can impact biofilm formation and stress adaptation in other bacteria (Lemos et al., 2005), though its role in V. cholerae remains unclear. We hypothesize that VC1123 may function as part of a PhrR-associated regulatory module that links environmental or redox sensing to outer membrane remodeling. Loss of VC1123 disrupts this regulatory balance, leading to reciprocal regulation of OmpU and OmpT and thereby enhancing intestinal fitness independently of the canonical ToxR/ToxS pathway.
In summary, our findings reveal that VC1123, a highly conserved gene of unknown function, acts as a suppressor of intestinal colonization, likely by suppressing ompU and/or promoting ompT through a ToxR-independent mechanism. The enhanced virulence of the ΔVC1123 mutant underscores the importance of fine-tuning outer membrane composition during host infection. Future studies should aim to (i) define the molecular function of the VC1123 protein, (ii) identify its regulatory targets and interacting partners, and (iii) determine whether VC1123 expression is modulated by host or microbial cues in the gut. Such insights may uncover novel layers of virulence control in V. cholerae and inform strategies to disrupt colonization.
This study has several limitations. Functional analyses of VC1123 were conducted exclusively in the N16961 genetic background, and thus it remains to be determined whether the observed effects are conserved across other seventh-pandemic or new-lineage V. cholerae strains. In addition, although deletion of VC1123 resulted in reciprocal regulation of ompU and ompT and enhanced intestinal colonization, we have not directly demonstrated that porin imbalance is the causal driver of the colonization phenotype. Future studies using multiple strain backgrounds and targeted genetic epistasis analyses will be required to establish the generality of VC1123 function and to definitively link OmpU/OmpT dysregulation to intestinal fitness.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Albert M. J. Siddique A. K. Islam M. S. Faruque A. S. Ansaruzzaman M. Faruque S. M. . (1993). Large outbreak of clinical cholera due to Vibrio cholerae non-O 1 in Bangladesh. Lancet 341:704. doi: 10.1016/0140-6736(93)90481-U, 8095621 · doi ↗ · pubmed ↗
- 2Chakrabarti S. R. Chaudhuri K. Sen K. Das J. (1996). Porins of Vibrio cholerae: purification and characterization of Omp U. J. Bacteriol. 178, 524–530. doi: 10.1128/jb.178.2.524-530.1996, 8550475 PMC 177687 · doi ↗ · pubmed ↗
- 3Champion G. A. Neely M. N. Brennan M. A. Di Rita V. J. (1997). A branch in the Tox R regulatory cascade of Vibrio cholerae revealed by characterization of tox T mutant strains. Mol. Microbiol. 23, 323–331. doi: 10.1046/j.1365-2958.1997.2191585.x 9044266 · doi ↗ · pubmed ↗
- 4Crawford J. A. Kaper J. B. Di Rita V. J. (1998). Analysis of Tox R-dependent transcription activation of omp U, the gene encoding a major envelope protein in Vibrio cholerae. Mol. Microbiol. 29, 235–246. doi: 10.1046/j.1365-2958.1998.00925.x, 9701817 · doi ↗ · pubmed ↗
- 5Devault A. M. Golding G. B. Waglechner N. Enk J. M. Kuch M. Tien J. H. . (2014). Second-pandemic strain of Vibrio cholerae from the Philadelphia cholera outbreak of 1849. N. Engl. J. Med. 370, 334–340. doi: 10.1056/nejmoa 1308663, 24401020 · doi ↗ · pubmed ↗
- 6Duret G. Delcour A. H. (2006). Deoxycholic acid blocks vibrio cholerae Omp T but not Omp U porin. J. Biol. Chem. 281, 19899–19905. doi: 10.1074/jbc.m 602426200, 16670088 · doi ↗ · pubmed ↗
- 7Fu X. Wang D. Yin X. Du P. Kan B. (2014). Time course transcriptome changes in Shewanella algae in response to salt stress. P Lo S One 9:e 96001. doi: 10.1371/journal.pone.009600124789066 PMC 4006864 · doi ↗ · pubmed ↗
- 8Fu X. Zhang J. Liang W. Diao B. Pang B. Kan B. (2025). Inflammation induced by the new lineage of Vibrio cholerae serogroup O 1 in the neonatal mouse model. Front. Immunol. 16:1617803. doi: 10.3389/fimmu.2025.1617803, 41112267 PMC 12531251 · doi ↗ · pubmed ↗