Novel insights into how gestational diet affects maternal-infant microbiota: a cross-sectional causal mediation analysis at one month postpartum
Eduard Flores Ventura, Sergio Ruiz-Saavedra, Raúl Cabrera-Rubio, Cecilia Martínez-Costa, Sonia González, Maria Carmen Collado

TL;DR
A Mediterranean-style diet during pregnancy is linked to healthier infant gut microbiota through effects on maternal gut bacteria.
Contribution
Identifies maternal Coprococcus as a key mediator linking gestational diet to infant gut microbiota composition.
Findings
Higher Mediterranean diet scores correlated with increased infant gut diversity and lower Veillonella abundance.
Maternal Coprococcus mediated the link between diet and infant microbiota, reducing Veillonella in infants.
Pro-inflammatory diets were associated with higher infant Veillonella and lower gut diversity.
Abstract
Maternal diet shapes maternal and infant microbiota, influencing early-life health. This study assessed associations between maternal dietary indices and infant gut microbiota, and the mediating role of maternal core taxa. In the MAMI cohort, pregnancy diet was evaluated using the Modified Mediterranean Dietary Score (MMDS), Dietary Quality Index (DQI), Healthy Eating Index (HEI), and Dietary Inflammatory Index (DII). Perinatal data, clinical records, and infant gut microbiota (1 month postpartum, 16S rRNA sequencing) were analyzed using clustering, regression, and causal mediation analysis (CMA). Among 104 mother–infant pairs, DII was inversely correlated with MMDS, HEI, and DQI. Higher MMDS (β = 0.785, p = 0.037) and lower DII (β = –0.783, p = 0.037) were associated with increased infant Veillonella relative abundance; MMDS was also positively linked to Shannon and Simpson…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
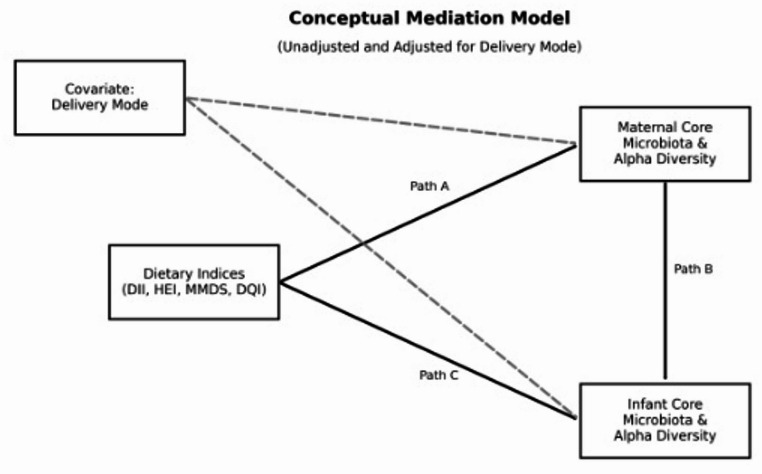 Figure 1
Figure 1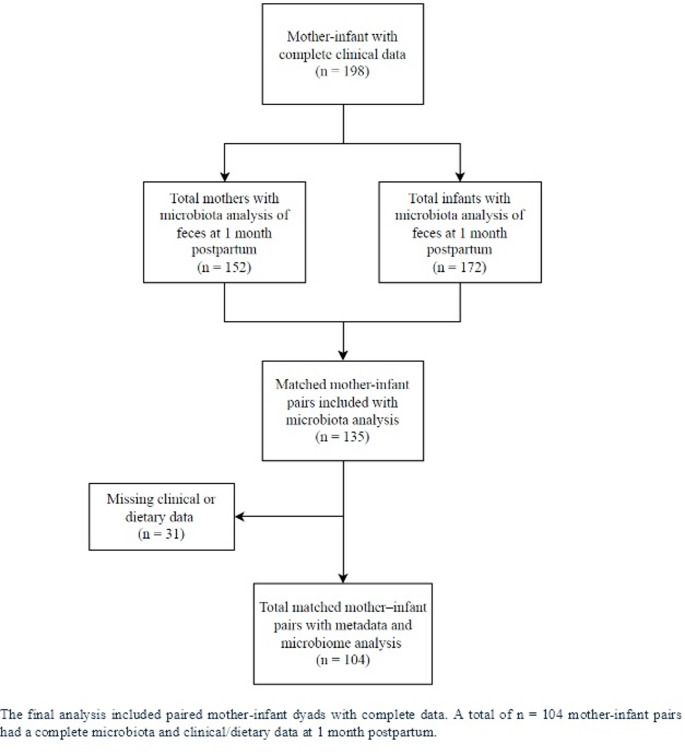 Figure 2
Figure 2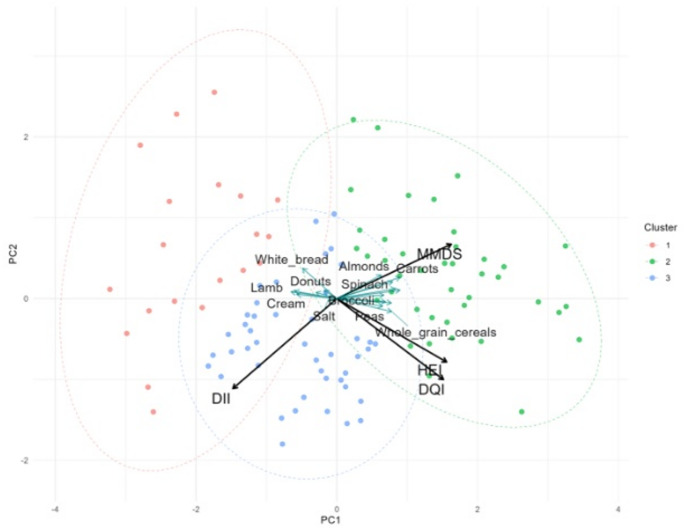 Figure 3
Figure 3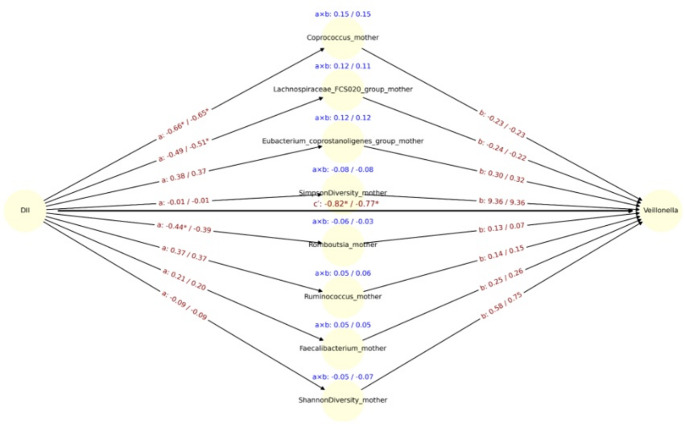 Figure 4
Figure 4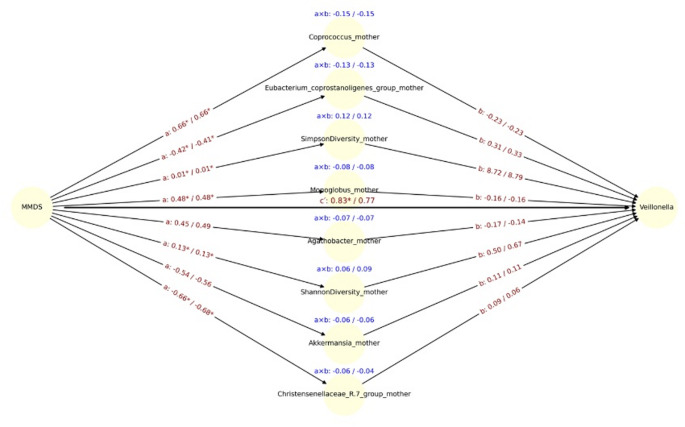 Figure 5
Figure 5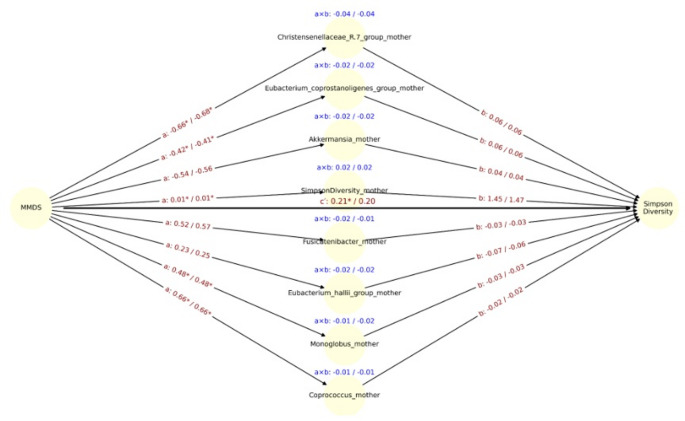 Figure 6
Figure 6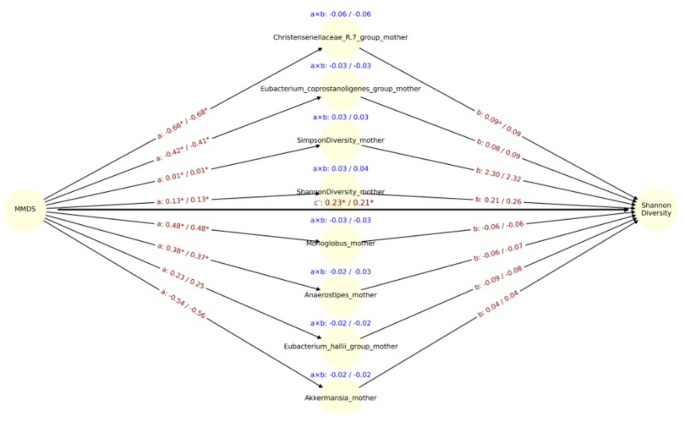 Figure 7
Figure 7- —Consejo Superior de Investigaciones Cientificas (CSIC)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Nutritional Studies and Diet · Health, Environment, Cognitive Aging
Background
Alteration on the infant gut microbiota have been linked to immune-related conditions, and higher risk of non-communicable diseases (NCDs) with implications for health programming [1–5]. Early life microbial colonisation and assembly is known to beshaped by several perinatal factors, including maternal nutritional status, mode of delivery, antibiotic use and breastfeeding practices [6, 7]. The maternal microbiota is the main microbial source for the infant gut, that is characterised by lower microbial diversity and higher prevalence of Bifidobacterium members [8–10], which have been also reported to be influenced by the maternal diet [11–15]. Where maternal gut Bifidobacterium and Bacteroides strains are vertically transferred and they are adapted to persist in the infant gut [16–18] and also, breastfeeding practices influence the microbiota [9, 19] and is linked to reduce risk of infectious diseases and allergies development later in life [20–24].
The maternal diet have been reported to modulate some bioactive compounds present in human milk as HMO [25], microRNA [26] and also, microbiota [27] with potential consequences on the infant microbiota evolution. Although the exact mechanisms is unclear, some studies have reported. For example, maternal intake of polyphenols can bind to and alter cell membrane functions, inhibiting bacterial growth [28], and the anaerobic fermentation of dietary fibers and HMO produces short-chain fatty acids and other metabolites that impact microbial composition and activity [29]. Consequently, maternal diet significantly shapes both maternal and infant microbiota, impacting infant health and supporting the 'thrifty phenotype hypothesis’ [30].
Until now, literature has often studied maternal dietary exposure to specific foods, such as fish oil [12], salmon [31], and nutrients including omega-3 fatty acids [32], vitamin D [13, 33], and fat intake [34], as well as food groups or clusters [11, 35, 36] in relation to infant gut microbiota. However, there is limited evidence [14] analysing the impact that dietary patterns may have on the microbiota in children.
In this scenario, we aim to assess the influence of maternal diet on infant core microbiota measured by several dietary indices and to investigate the potential mediation by maternal core gut microbiota of the effect of maternal diet on the core gut microbiota of exclusively breastfed infants (Fig. 1).Fig. 1. Conceptual Mediation Model representing the hypothesized paths between maternal dietary indices and infant gut outcomes. Path A illustrates the association between maternal Dietary Indices (DII, HEI, MMDS, DQI) and maternal core microbiota and alpha diversity (mediator). Path B represents the link from maternal core microbiota and alpha diversity to infant core microbiota and alpha diversity (outcome). Path C denotes the direct effect of maternal dietary indices on infant gut microbial outcomes. Dashed arrows indicate covariate adjustment for delivery mode. Maternal and infant core microbiota are defined as taxa present in at least 50% of samples above 0.001% relative abundance
Materials and methods
Study design and volunteers
This study is a cross-sectional analysis involving mother–infant dyads from the Maternal Microbiome (MAMI) cohort, as outlined in the flowchart (Fig. 2). Maternal and infant samples were collected at 1 month postpartum (n = 104 pairs). The following maternal-infant clinical parameters were collected: gestational age, pregestational BMI, gestational weight gain, antibiotics during pregnancy, antibiotics during labor, delivery mode, sex, weight at birth, and antibiotics 7 to 15 days post-partum, amongst others. Maternal-infant biosamples, feces and human milk, were collected at 1 month post-partum, respectively. Details about the recruitment, sampling methods and storage were described previously [37]. In brief, the recruitment was conducted in primary health centers and hospitals. Eligible mothers were over 18, had a healthy pregnancy, and spoke Spanish, excluding those with certain medical conditions or on medication. Participants received oral and written information about the study, and written consent was obtained from each participant. The study was approved by the Ethical Committee of the Hospital Clínico Universitario Valencia, Spain and by CSIC Ethics Committee.Fig. 2. Participants flowchart. The final analysis included paired mother-infant dyads with complete data. A total of n = 104 mother-infant pairs had a complete microbiota and clinical/dietary data at 1 month postpartum
This study followed the STROBE-nut (Strengthening the Reporting of Observational Studies in Epidemiology—Nutrition) [38] reporting guidelines for observational studies (Electronic Supplementary Material Table S1).
Maternal dietary assessment and diet-related indexes
Dietary records were collected during the first weeks after birth by a nutritionist using a 140-item food frequency questionnaire (FFQ) as described by [39]. FFQ information was analyzed for the energy and daily intake of macro- and micronutrients by using the nutrient Food Composition Tables developed by the Centro de Enseñanza Superior de Nutrición Humana y Dietética (CESNID) [40]. Based on a previously validated FFQ [14], specific dietary indices were calculated, including the Dietary Inflammatory Index (DII), associated with adverse health outcomes, and others linked to positive health effects, such as the Diet Quality Index (DQI), the Modified Mediterranean Diet Score (MMDS), and the Healthy Eating Index (HEI):
The modified dietary inflammatory index (DII)
The DII quantifies the inflammatory potential of diets using a comprehensive literature review [41]. This method involved calculating 35 out of 45 parameter-specific inflammatory effect score. Using IBM SPSS Statistics 25.0 [42], individual dietary intake data were converted to Z-scores, percentile scores, and finally centered percentile scores, which were multiplied by food parameter effect scores. Summing these scores provided the overall DII, ranging from − 8.87 (anti-inflammatory) to 7.98 (pro-inflammatory).Table 1. Participant maternal and infant clinical characteristicsCharacteristics1 month postpartum (n = 104)Maternal characteristicsGestational age (weeks)40 [39, 40]Pregestational BMI, kg/m222.46 [20.5–24.05]Gestational weight gain, kg*12 [10–15]*Antibiotics during pregnancy (n)*Yes29 (29/104, 27.88%)No75 (75/104, 72.12%)*Antibiotics during labor (n)*Yes29 (29/104, 27.88%)No75 (75/104, 72.12%)Delivery mode (n)C-section27 (27/104, 25.96%)Vaginal77 (77/104, 74.04%)Infant characteristics †Sex (n)Female60 (60/104, 57.69%)Male44 (44/104, 42.31%)Weight at birth (kg)3.29 [3–3.58]Antibiotics 7 to 15 days post-partumYes6 (6/104, 5.77%)No98 (98/104, 94.23%)Maternal dietary characteristicsDII0.6 [− 2.25–2.52]DQI64 [55, 58–68]MMDS4 [3–5]HEI71.88 [65.39–76.27]*Median [Q1-Q3]^†^All infants were exclusively breastfed from birth
The diet quality index (DQI)
The DQI score, ranging from 0 to 100, was used to evaluate diet quality by matching individual dietary intake data with recommended servings and nutrient intakes, then categorizing them into food groups. Each food group's intake was scored on a scale, with higher scores indicating better dietary quality, using the scoring criteria from Table 2 of Mariscal-Arcas et al. [43]. Components such as variety, adequacy, moderation, and overall balance were assessed. Scores for each category were calculated based on specific criteria and summed to provide an overall DQI score. To integrate the DQI calculation in IBM SPSS Statistics 25.0, each component in the sample was evaluated by assigning a value of 0 to 3, 0 to 5, or 0 to 6, as appropriate, then scores were added to obtain the total DQI score for each individual.Table 2. Linear regression results showing statistically significant associations (p < 0.05) between dietary indices and bacterial generaBacteria and diversityDietβ [95% CI]P valueMembers of infant gut microbiota core (genus level)**VeillonellaDII-0.783 [-1.520, -0.047]0.037VeillonellaMMDS0.785 [0.049, 1.522]0.036Infant diversity indicesShannonMMDS0.211 [0.018, 0.403]0.032SimpsonMMDS0.194 [0.001, 0.387]0.048Members of maternal gut microbiota core (genus level)**RomboutsiaDII-0.439 [-0.869, -0.009]0.045CoprococcusDII-0.655 [-1.155, -0.155]0.011ButyricicoccusDQI0.715 [0.096, 1.334]0.023FaecalibacteriumDQI0.432 [0.037, 0.827]0.032FusicatenibacterHEI0.647 [0.072, 1.223]0.027Incertae_SedisHEI-0.395 [-0.715, -0.076]0.015FaecalibacteriumHEI0.442 [0.049, 0.835]0.027BifidobacteriumHEI-0.577 [-1.082, -0.073]0.025Eubacterium coprostanoligenes groupMMDS-0.416 [-0.8, -0.031]0.034AnaerostipesMMDS0.381 [0.069, 0.692]0.017MonoglobusMMDS0.482 [0.108, 0.856]0.011CoprococcusMMDS0.658 [0.158, 1.158]0.01BifidobacteriumMMDS-0.549 [-1.053, -0.045]0.033Christensenellaceae R.7 groupMMDS-0.662 [-1.072, -0.251]0.001Maternal diversity indicesChao1HEI0.204 [0.011, 0.397]0.038ShannonHEI0.249 [0.058, 0.441]0.011InvSimpsonHEI0.269 [0.0788, 0.459]0.006ShannonMMDS0.219 [0.028, 0.412]0.025InvSimpsonMMDS0.242 [0.051, 0.432]0.013SimpsonMMDS0.205 [0.012, 0.398]0.037Each row displays the estimated regression coefficient and rawp-value for the relationship between one of four dietary scores.DII, MMDS, DQI, and HEI and the relative abundance of a microbialtaxon.Positive coefficients indicate a direct association, while negative valuesreflect inverse relationships. FDR-adjusted p-values were no longer significant after correction areindicated accordingly
The modified mediterranean diet score (MMDS)
The MMDS ranges from 0 to 10 and evaluates adherence to a Mediterranean diet [44]. For each component typically consumed in high quantities (vegetables, legumes, fruit/nuts, fish/seafood, and cereals). To integrate the MMDS calculation in IBM SPSS Statistics 25.0, the population medians were obtained for each variable to be analyzed (except alcohol). Most variables can be taken directly from the database, while others had to be created (e.g., MMDS_fruit_nuts, MMDS_fish_seafood, and MMDS_fat_ratio, calculated as the the fraction of monounsaturated and polyunsaturated fatty acids relative to saturated fatty acids). The values below the median were recoded as 0 and values above the median as 1. For meat, meat products and dairy products, the coding was reversed. For alcohol, the scores were based on whether individuals fall within the recommended range (1 if within the range, 0 if outside). Finally, we added the scores of all the variables to obtain the total MMDS score for each mother.
The healthy eating index (HEI)
The HEI-2015 evaluates diet quality based on 13 components, scored from 0 to 100, where a higher score indicates a healthier diet, and a lower score suggests a less healthy diet [45]. The calculation relied on densities, the amount of a dietary component per 1,000 kcal. These densities were compared to established standards. Regarding the calculation, to integrate the HEI-2015 calculation in IBM SPSS statistics 25.0, the servings for scoring each individual from the provided Table 1 from Krebs-Smith et al. [45] were obtained. Next, the food group division as stablished in Electronic Supplementary Material Table S2. was used to categorize foods groups. For each food group, the amount of cups, ounces, grams, or percentage consumed by the individual was determined. These scores are detailed in Krebs-Smith et al. [45]. Finally, the total scores were added to determine each individual's overall HEI score.
Biomaterials and microbiota profiling by 16S amplicon sequencing
Fecal samples from mothers were collected at 1 month postpartum. Infant fecal samples were collected at the 1-month follow-up visit. These samples were gathered one month after childbirth using a standardized procedure well-documented in previous studies [46]. In summary, feces from both mothers and infants were collected in sterile containers, infant fecal samples were collected at home by their parents who were previously trained by clinical personnel in the health care centres where they were enrolled, morning collection was recommended. Briefly, fecal samples were deposited in provided sterile containers and immediately kept at − 20 °C, before the final storage at − 80 °C until further analysis.
Fecal DNA was extracted as previously described [15] using the Master-Pure DNA Extraction Kit (Epicentre, WI, USA) with physical and enzymatic modifications. Microbial profiling targeted the V3–V4 region of the 16S rRNA gene and was sequenced on an Illumina MiSeq platform (2 × 300 bp, MiSeq Reagent Kit v3) at FISABIO (Valencia, Spain). Sequence processing was done with DADA2 [47], and taxonomic assignment used the Silva v132 database [48]. Genus-level relative abundances and alpha-diversity metrics (Observed, Chao1, Shannon, Simpson, InvSimpson, Fisher) were calculated using the Phyloseq package (v4.3.1) [49]. Tables at genus taxonomical level and alpha-diversity indexes were used in this study to combine with the dietary indexes. Prior to regression analysis, maternal and infant bacterial counts were transformed into relative abundances. Subsequently, the core microbiota for both maternal and infant groups was defined as taxa present in at least 50% of the samples above 0.001% relative abundance. Then centered log-ratio transformed (CLR) was applied to bacterial relative abundances.
Data processing and statistical methods
Complete data, including clinical, dietary, and gut microbiota information, was available for 104, mother-infant pairs at 1 month postpartum, respectively, as described in Fig. 2. Data preprocessing involved z-score normalization of dietary indices, the individual food components of those indices, and diversity indices, as well as centered log ratio (CLR) transformation of bacterial relative abundances.
A Principal Component Analysis (PCA) was performed to explore the relationships among various maternal dietary indices, including the DQI, DII, MMDS, and HEI. This analysis was executed using the prcomp function from the stats package in R [50], which facilitated the reduction of dimensionality by transforming the original variables into a new set of uncorrelated principal components. The PCA results were visualized through a biplot, which was generated using the ggplot2 [51] and ggpubr [52] libraries. Additionally, cluster analysis was incorporated by applying the k-means method to the PCA scores to discern distinct nutritional behavior patterns. The optimal number of clusters was determined using the elbow method, implemented via the fviz_nbclust function from the factoextra package [53].
Linear regression models
To explore associations between dietary quality and gut microbiota composition, we performed a series of linear regression models using each dietary score, DII, MMDS, DQI, and HEI, as independent variables. Microbial taxa (normalized relative abundances via CLR transformation) were treated as dependent variables in univariate models. We used the statsmodels Python package [54] to fit ordinary least squares (OLS) regression models of the form genus (core member of the maternal and infant gut microbiota) ~ diet score, for each combination of microbial taxon and dietary index. Raw p-values were calculated for each regression, and false discovery rate (FDR) correction was applied across all tests using the Benjamini–Hochberg method to account for multiple comparisons. Associations with p < 0.05 before and after correction were considered for further interpretation. All analyses were conducted in Python (v3.12.7) [55] using pandas [56], statsmodels [54], statsmodels.stats.multitest [54], and joblib [57] for parallel computation.
Causal mediation analysis (CMA)
To investigate whether the elements previously identified as significant in preliminary univariate regressions (p < 0.05) were associated with infant gut core microbiota and diversity, we conducted a CMA using a two-model regression framework (Electronic Supplementary Material Table S3).
For each selected maternal taxon as a potential mediator, we fit two regression models:
- Mediator model:
where MiM_iMi is the CLR-transformed abundance of the mediator taxon, XiX_iXi is the standardized maternal dietary score (e.g., MMDS), and CiC_iCi is the covariate for delivery mode.
-
a-path (a = β1) represents the effect of the dietary score on the mediator taxon.
-
2.Outcome model:
where YiY_iYi is the CLR-transformed outcome (e.g., infant Simpson diversity or Veillonella), MiM_iMi is the mediator taxon, and CiC_iCi is the delivery mode covariate.
- b-path (b = β3) represents the effect of the mediator on the outcome, controlling for the dietary score.
- Direct effect (c′-path) (γ1) captures the effect of the dietary score on the outcome independent of the mediator.
The indirect effect was calculated as the product of the a and b paths (a × b), quantifying the mediation effect of each taxon. The total effect was derived as the sum of the direct and indirect effects:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$Total Effect= \gamma 1+(a\times b)$$\end{document}To assess the significance of the indirect effect, we employed nonparametric bootstrapping with 1,000 iterations to generate empirical 95% confidence intervals and two-tailed pseudo p-values. Only models with complete data for all included variables were used. All analyses were adjusted for delivery mode.
Results
Maternal and infant characteristics and gut microbiota core one month post-partum
General characteristics of participants are available in Table 1. This study evaluated maternal and infant characteristics considering one-month maternal and infant gut microbiota core. Approximately 25% of the deliveries were via cesarean section, and the majority were vaginal (74%). Infant gender distribution was more female than male. Antibiotic use during pregnancy and labor was reported in about 28 of the cases. Maternal dietary indices such as DII, DQI, MMDS, and HEI showed minimal variation over the time points.
Maternal dietary indices explain different patterns.
K-means clustering revealed three distinct dietary patterns, characterized by higher scores on the MMDS, elevated DQI and HEI, and a predominance of DII. The PCA biplot (Fig. 3) illustrates the relationships among dietary indices, with DQI and HEI closely aligned, reflecting their positive correlation (r = 0.59), and shared focus on overall dietary quality. In contrast, DII is negatively correlated with MMDS (r = –0.61), HEI (r = –0.40), and DQI (r = –0.29). These patterns are further supported by food-level correlations, where pro-inflammatory foods (e.g., processed meats, refined products) align with DII, and anti-inflammatory, nutrient-dense foods (e.g., legumes, fruits, vegetables, whole grains) align with MMDS, DQI, and HEI.Fig. 3PCA biplot illustrating sample clustering and the contributions of dietary indices (black arrows) and the union of food items ranked among the eight most positively correlated variables for each dietary index (turquoise arrows). Samples are colored according to dietary pattern clusters (K-means, K = 3), with 95% confidence ellipses illustrating the separation among clusters. Arrow directions indicate the alignment of variables with the principal components
Maternal diet and infant gut microbiota linear regressions
Significant associations were observed between maternal and infant gut microbiota composition and several dietary indices (Table 2). In infants, Veillonella was inversely associated with the DII (β = − 0.783, 95% CI: [− 1.520, − 0.047], p = 0.037) and positively associated with the MMDS (β = 0.785, 95% CI: [0.049, 1.522], p = 0.037). Infant alpha diversity metrics also showed positive associations with MMDS, including Shannon diversity (β = 0.211, 95% CI: [0.018, 0.403], p = 0.032) and Simpson diversity (β = 0.194, 95% CI: [0.001, 0.387], p = 0.049).
Among maternal gut microbiota genera, Coprococcus was inversely associated with DII (β = − 0.655, 95% CI: [− 1.155, − 0.155], p = 0.011), and positively with MMDS (β = 0.658, 95% CI: [0.159, 1.159], p = 0.010). Additional taxa, including Fusicatenibacter, Faecalibacterium, Incertae Sedis, and Bifidobacterium, showed significant relationships with HEI and other diet scores. Maternal diversity indices such as Chao1 (β = 0.204, 95% CI: [0.011, 0.397], p = 0.038), Shannon (β = 0.249, 95% CI: [0.059, 0.441], p = 0.011), and Inverse Simpson (β = 0.269, 95% CI: [0.079, 0.459], p = 0.006) were also positively associated with HEI.
However, none of the associations remained statistically significant after FDR correction, indicating potential Type I error due to multiple testing.
CMA of maternal diet and infant gut microbiota: the mediating role of maternal microbiota
To evaluate whether specific maternal microbial taxa mediated the relationship between DII and the relative abundance of Veillonella, we conducted CMA across core maternal genera and diversity indices. Among the top eight coefficients ranked by the magnitude of the indirect effect (a × b), three genera; Coprococcus, Lachnospiraceae FCS020 group, and Eubacterium coprostanoligenes group, showed consistent and relatively large mediation effects (unadjusted a × b ≥ 0.12), with comparable results after adjusting for delivery mode.
Notably, Coprococcus exhibited the strongest indirect effect (a × b = 0.15), driven by a strong inverse association between DII and Coprococcus (a = –0.66, p < 0.05), and a negative link between Coprococcus and Veillonella (b = –0.23). Adjustment for delivery mode did not substantially alter these coefficients (a × b = 0.15; a = –0.65, b = –0.23) (Fig. 4).Fig. 4. Mediation path diagram illustrating associations between maternal DII and the relative abundance of Veillonella in infants, via maternal gut microbial genera. Each node represents a mediator genus (center), the exposure (DII, left), or the outcome (Veillonella, right). Arrows represent estimated regression coefficients from the mediation models: a-path: effect of DII on the mediator genus, b-path (right): effect of the mediator genus on Veillonella, c′-path (center): direct effect of DII on Veillonella, adjusted for the mediators. For each path a, b, and a × b (indirect effect), both unadjusted and delivery-mode, adjusted estimates are shown as: estimate_unadj / estimate_adj. Asterisks (*) indicate statistical significance (p < 0.05) for the corresponding coefficient. The c′path at the center reflects the mean direct effect of DII on Veillonella across all models. Indirect effects (a × b) are displayed in blue above each mediator. Only the top 8 genera with the largest unadjusted indirect effects (by absolute value) are shown
The MMDS was significantly associated with lower maternal Christensenellaceae abundance (path a: β = −0.66, p = 0.0014), and Christensenellaceae was positively associated with infant Shannon diversity (path b: β = 0.09, p = 0.047), although after adjustment for delivery mode significance was lost (Fig. 5). In contrast, although diet was significantly associated with maternal microbiota, no significant mediators were identified for Simpson diversity or Veillonella abundance, as no microbial genera demonstrated simultaneous significance on both path a and path b (Fig. 6). The direct effect of the DII on Veillonella abundance remained statistically significant after adjusting for delivery mode (cʹ = − 0.82 unadjusted / − 0.77 adjusted, p < 0.05 for both models). Similarly, in models using the MMDS as the exposure, direct effects were also significant for Veillonella (c′ = 0.83 / 0.77, p < 0.05 for unadjusted model), Simpson diversity (c′ = 0.21 / 0.20, p < 0.05 for unadjusted model), and Shannon diversity (c′ = 0.23 / 0.21, p < 0.05 for both models) (Fig. 7). These findings suggest that the observed associations between maternal diet and infant microbiota or diversity metrics are not fully mediated by maternal gut microbial genera and may reflect direct influences of maternal diet or dietary-associated host factors.Fig. 5. Mediation path diagram illustrating associations between maternal MMDS and the relative abundance of Veillonella in infants, via maternal gut microbial genera. Each node represents a mediator genus (center), the exposure (MMDS, left), or the outcome (Veillonella, right). Arrows represent estimated regression coefficients from the mediation models: a-path: effect of MMDS on the mediator genus, b-path (right): effect of the mediator genus on Veillonella, c′-path (center): direct effect of MMDS on Veillonella, adjusted for the mediators. For each path a, b, and a × b (indirect effect), both unadjusted and delivery-mode, adjusted estimates are shown as: estimate_unadj / estimate_adj. Asterisks () indicate statistical significance (p < 0.05) for the corresponding coefficient. The c′path at the center reflects the mean direct effect of DII on Veillonella across all models. Indirect effects (a × b) are displayed in blue above each mediator. Only the top 8 genera with the largest unadjusted indirect effects (by absolute value) are shownFig. 6Mediation path diagram illustrating associations between maternal MMDS and the relative abundance of Simpson diversity in infants, via maternal gut microbial genera. Each node represents a mediator genus (center), the exposure (MMDS, left), or the outcome (Simpson diversity, right). Arrows represent estimated regression coefficients from the mediation models: a-path: effect of MMDS on the mediator genus, b-path (right): effect of the mediator genus on Simpson diversity, c′-path (center): direct effect of DII on Simpson diversity, adjusted for the mediators. For each path a, b, and a × b (indirect effect), both unadjusted and delivery-mode, adjusted estimates are shown as: estimate_unadj / estimate_adj. Asterisks () indicate statistical significance (p < 0.05) for the corresponding coefficient. The c′path at the center reflects the mean direct effect of DII on Simpson diversity across all models. Indirect effects (a × b) are displayed in blue above each mediator. Only the top 8 genera with the largest unadjusted indirect effects (by absolute value) are shownFig. 7Mediation path diagram illustrating associations between maternal MMDS and the relative abundance of Shannon diversity in infants, via maternal gut microbial genera. Each node represents a mediator genus (center), the exposure (MMDS, left), or the outcome (Shannon diversity, right). Arrows represent estimated regression coefficients from the mediation models: a-path: effect of MMDS on the mediator genus, b-path (right): effect of the mediator genus on Shannon diversity, c′-path (center): direct effect of DII on Shannon diversity, adjusted for the mediators. For each path a, b, and a × b (indirect effect), both unadjusted and delivery-mode, adjusted estimates are shown as: estimate_unadj / estimate_adj. Asterisks (*) indicate statistical significance (p < 0.05) for the corresponding coefficient. The c′path at the center reflects the mean direct effect of DII on Shannon diversity across all models. Indirect effects (a × b) are displayed in blue above each mediator. Only the top 8 genera with the largest unadjusted indirect effects (by absolute value) are shown
Discussion
We observed that maternal diet influences the infant gut microbiota at one month of age, and this effect is partially mediated by maternal microbiota changes related to diet during gestation. Specifically, the Veillonella genus, Shannon and Simpson diversity indices in the infant gut were partially mediated by members of the maternal gut core and diversity. A pro-inflammatory maternal diet was associated with a direct reduction in Veillonella abundance. At the same time, maternal gut microbiota at the genus level partially mediated this effect. For instance, genera such as Coprococcus appeared to partially mediated an increase in Veillonella in response to DII, while others, such as Romboutsia, partially mediated a decrease. Additionally, opposing effects of maternal MMDS and DII on Veillonella were observed in the CMA model. MMDS was positively associated with Shannon and Simpson diversity indices. However, when mediation models included specific members of the maternal gut microbiota, opposing effects emerged, further highlighting the complexity of microbial interactions.
This is the second study to use dietary indices in the MAMI cohort [14]. Compared to other populations, our cohort shows similar adherence to dietary indices (Table 3). In terms of the HEI, other populations have also shown high adherence, indicating a lower preference for Western dietary patterns [58–61]. However, one study noted that 80% of its U.S. cohort exhibited lower adherence to the HEI [62]. Regarding the DII, which measures the inflammatory potential of dietary components, the scores are distributed more evenly across studies, reflecting varied dietary influences within the same populations [63].Table 3. Table comparison of studies reporting maternal dietary index during gestationReferencesPopulationSample size (n)CohortDIIDQIMMDSHEIRodriguez et al. [66]Spain54INMANANANA48–51Shapiro et al. [67]United States1079The Healthy StartNANANA54.2 (13.6)Asadi et al. [68]Iran350NANANANA70.1 (11.11)Sivula et al. [69]Finland1330KuBiCo − 0.095 (2.3)NANA58.1 (10.1)Monthé et al. [70]United States1459Project Viva − 2.6 (1.4)NA4.6 (2)61 (10)Almulla et al. [71]United Arab Emirates122Mutaba’ah StudyNANA4.5 (0.5)62.5 (2.5)Garcia et al. [72]Spain86MAMINANA8.8 (1.7)NALeda et al. [73]Spain2461INMANANA3.8 (1.6)NALeda et al. [73]Greece889RHEANANA4 (1.7)NAHayat et al. [74]Iran90NANA52.64(7.94)NANASouza et al. [75]Brazil260NA0.04 (3.2)68.82 (80)NANAThis studySpain246MAMI0.58 (4.61)64.5 (11.41)4 (1)71.92 (11)NA not available, DII dietary inflammatory index, MMDS modified mediterranean dietary score, HEI healthy eating index, MAMI maternal microbe cohort, INMA Infancia y Medio Ambiente—environment and childhood, RHEA mother–child cohort in Crete, and KuBiCo Kuopio birth cohort*Values correspond to median (interquartile range)
The lactate-utilizing bacterium Veillonella is a prominent early colonizer of the infant gut microbiota. Studies have identified vertical transmission events of Veillonella strains, particularly via human milk (72) and maternal fecal samples, contributing to initial gut colonization [64]. However, in our cohort, Spearman correlation tests between maternal and infant Veillonella gut abundance at one month postpartum did not reveal a significant association (ρ (rho) = –0.016, p = 0.872). Moreover, Veillonella has not been detected as the most frequently transmitted bacterial genera [65].
Despite this, Veillonella plays a beneficial role in shaping the infant gut microbial community. Its metabolic functionality, primarily driven by the methylmalonyl-CoA pathway along with other reductive pathways, enables energy production under the low-redox conditions typical of the infant colon. This allows Veillonella to metabolize lactate, derived from other microbial taxa and dietary sources, into short-chain fatty acids (SCFAs) such as propionate and acetate, which support gut health [76]. A decreased abundance of Veillonella has been associated with a higher risk of asthma [77]. As an obligate anaerobic, Gram-negative bacterium, Veillonella contributes to gut homeostasis, playing a key role in immune modulation, epithelial barrier integrity, and luminal pH regulation [78, 79].
Our mediation models show that maternal dietary patterns during pregnancy exert both direct and indirect influences on Veillonella abundance in infants at one month. The DII negatively impacted Veillonella levels, while a MMDS showed generally positive associations. These effects were partially mediated through specific members of the maternal gut microbiota at one month postpartum, including genera such as Coprococcus, Romboutsia, Lachnospiraceae, and Eubacterium, many of which are involved in upstream fermentation of complex carbohydrates [80–83] and can alter the availability of Veillonella’s primary substrate, lactate. These microbial shifts may also play a role in modulating infant gut microbial diversity, particularly in response to Mediterranean dietary patterns during pregnancy.
This reflects broader ecological and metabolic interdependencies: Veillonella thrives in environments where lactate is abundant and is likely to be in a metabolically active, quasi-exponential (log) phase under such conditions [84]. When lactate becomes limited, whether due to substrate scarcity or competitive uptake by co-colonizing taxa, Veillonella may shift to a stationary-like metabolic state, characterized by reduced SCFA production, stress-response gene expression, and metabolic reprogramming. Such phase-dependent shifts have been described in vitro, with V. dispar showing reduced propionate output and transcriptional changes in central metabolic genes under nutrient-limited conditions [78, 84].
In vivo, these dynamics are further shaped by strain-level variability. Different Veillonella strains differ in lactate transporter efficiency, SCFA production ratios, and biofilm formation capacity, all of which may influence their ability to colonize and persist in the infant gut [79, 85, 86]. Quorum sensing systems [87], although not well-characterized in Veillonella, may play a role in coordinating population-level behaviors such as biofilm formation and substrate utilization in response to local microbial density or host signaling molecules, and autoinducers, and may be affected indirectly by maternal microbial ecology.
Importantly, the maternal gut microbiota at one month postpartum may reflect longer-term dietary influences, which in turn modulate microbial metabolite profiles (e.g., lactate availability) that affect vertically transmitted or environmentally acquired Veillonella strains in the infant. The observed mediation by maternal genera and diversity indices reinforces the importance of microbial cross-feeding, ecological succession, and nutrient shaping in early life microbiota assembly, particularly for key fermenters like Veillonella that occupy metabolic niches downstream of primary saccharolytic fermenters, such as Coprococcus, Eubacterium coprostanoligenes group, Agathobacter, and Christensenellaceae R-7 group [80, 88, 89].
Gestational adherence to a MMDS was positively associated with infant gut microbial diversity at one month, with effects partially mediated by the maternal core gut microbiota at one month postpartum. Key mediators included Coprococcus, Eubacterium hallii group, and Christensenellaceae R-7 group, fiber-fermenting taxa enriched by plant-based diets and known for SCFA production [90–92]. Although direct effects of MMDS on infant diversity were stronger, the microbial mediation paths suggest that postpartum maternal microbiota composition, shaped by earlier dietary exposures, contributes to infant ecological outcomes. Maternal adherence to a Mediterranean diet may therefore influence the early phases of microbial assembly. Although diversity is physiologically lower in early infancy than later in life, several cohorts suggest that relatively reduced diversity or delayed diversification during the first year is associated with adverse immune and allergic outcomes [93–95], rather than being beneficial per se. In this context, the MMDS associated increase in Simpson diversity at 1 month is likely to reflect a more advanced yet still age-appropriate maturation trajectory rather than dysbiosis, consistent with patterns typically observed in vaginally delivered versus caesarean-born infants, and in breastfed infants compared with those who discontinue breastfeeding [96, 97]. The accompanying enrichment of Veillonella spp., a characteristic early-life taxon that participates in cross-feeding networks, may indicate the emergence of a more functionally interconnected ecosystem shaped in part by maternal diet. However, as we did not measure microbiota function or long-term clinical outcomes, these implications remain speculative, and our findings should be interpreted as hypothesis-generating.
Strengths and limitations
Several limitations of this study must be acknowledged. First, a comprehensive understanding of microbiota modulation and the identification of microbial signatures would ideally require a multi-omics approach [98–100]. Second, more data are needed to elucidate the initial seeding of the infant gut microbiota, particularly the role of other influencing factors such as human milk composition. Finally, the cross-sectional design limits causal inference, as it captures only a single time point, one month postpartum, providing a snapshot rather than a dynamic view of microbiota development [101].
Nevertheless, an strength of the study is its focus on gestational exposures, a time window considered etiologically relevant for early-life development [102–105]. Methodologically, the use of the 'abundance-occurrence' method to define the core microbiota aligns with accepted practices [106]. However, this approach tends to emphasize the most consistently detectable and abundant taxa, potentially overlooking less abundant but potentially biologically significant taxa.
The application of CMA is another strength, as it enables exploration of potential biological mechanisms, particularly highlighting the maternal microbiota as a mediator of the relationship between maternal diet and infant gut microbiota [107–109].
Furthermore, the study utilized multiple dietary indices to assess maternal dietary intake, which enhances the robustness of dietary characterization and helps to address some methodological biases [110, 111]. However, dietary data were obtained via a FFQ administered during the early postpartum period, relying on retrospective recall of the previous year’s diet. This introduces recall bias and measurement noise, limiting the precision of dietary exposure assessment [112].
The dietary indices were based on international standards, which, while comprehensive, may not fully capture local dietary patterns. Given that the study population was based in a Mediterranean region (Valencia, Spain), this raises concerns about external validity and the generalizability of the findings to other populations or food environments.
Another limitation lies in the 16S rRNA gene sequencing approach used for microbial profiling. While it allows taxonomic classification, it has known limitations, including potential false positives, with up to 20% of predicted genera possibly being incorrect when using the SILVA database [113]. This introduces taxonomic bias and may reduce the accuracy of microbial composition data. Furthermore, strain-level variation within the same genus is often overlooked, meaning that associations observed at the genus level may not hold true when examined at a strain-level [114, 115]. Finally, 16S sequencing does not allow for accurate functional inference, limiting insights into metabolic pathways and microbial functionality [116]. As such, infant microbiota abundance may not directly reflect functional capacity.
Conclusion
This study demonstrates that maternal diet during gestation exerts both direct and microbiota-mediated effects on the infant gut microbiota at one month postpartum. Notably, dietary patterns such as MMDS and DII were associated with significant changes in Veillonella abundance and infant microbial diversity, effects partially mediated by maternal gut taxa including Coprococcus, Eubacterium, and Christensenellaceae R-7 group. These findings emphasize the ecological complexity of maternal-infant microbial transfer and underscore the importance of upstream saccharolytic fermenters in shaping substrate availability for early colonizers like Veillonella. While cross-sectional design and methodological constraints limit causal inference, this study highlights the value of incorporating microbial mediation models to disentangle the pathways by which maternal diet influences early-life microbiota development. Future research should employ metagenomic techniques and adopt longitudinal study designs to validate findings and enhance the understanding of these associations and better inform public health recommendations.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1Supplementary file2
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tablas de composición de alimentos del CESNID. In: SEÑ - Sociedad Española de Nutrición. https://www.sennutricion.org/es/2013/05/13/tablas-de-composicin-de-alimentos-del-cesnid. Accessed 8 Jan 2025
- 2Python Release Python 3.12.7. In: Python.org. https://www.python.org/downloads/release/python-3127/. Accessed 30 Apr 2025
- 3Pandas - Python Data Analysis Library. https://pandas.pydata.org/. Accessed 6 May 2025
- 4Joblib: running Python functions as pipeline jobs — joblib 1.5.0 documentation. https://joblib.readthedocs.io/en/stable/. Accessed 6 May 2025