Molecular serotyping and virulence assessment of Avibacterium paragallinarum strains circulating in Chinese poultry flocks
Jiaolong Wen, Zhangmin Liao, Lijuan Yin, Lin Zhu, Yanhua Xu, Xiaoling Wang, Gao Fan, Jianping Qin, Qingfeng Zhou, Wencheng Lin

TL;DR
This study examines the spread and virulence of Avibacterium paragallinarum in Chinese poultry, revealing regional and seasonal patterns and the dominance of certain serotypes.
Contribution
The study provides updated epidemiological data and molecular serotyping of A. paragallinarum strains in China.
Findings
A. paragallinarum prevalence was highest in Anhui, Guangxi, and Guangdong provinces.
Serotype A was the most common, followed by C and B, with phylogenetic clustering matching serotype classification.
Both serotype A and C isolates caused infectious coryza in SPF chickens, with direct inoculation leading to more severe symptoms.
Abstract
Infectious coryza, caused by Avibacterium paragallinarum (A. paragallinarum), is a highly contagious respiratory disease that poses a significant threat to poultry production in China. Updated information on the epidemiology, serotype distribution, and virulence of circulating strains is essential for formulating effective regional control and vaccination strategies. A total of 200 clinical samples from nine provinces were analyzed to determine the prevalence, serotype distribution, and seasonal dynamics of A. paragallinarum. The bacterial isolation, characterization, species-specific PCR, and molecular serotyping based on the hmtp210 gene were performed. Phylogenetic relationships were inferred using the neighbor-joining method. The pathogenicity of representative isolates (serotype A and C) was assessed using SPF chickens. A. paragallinarum prevalence varied geographically, with the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
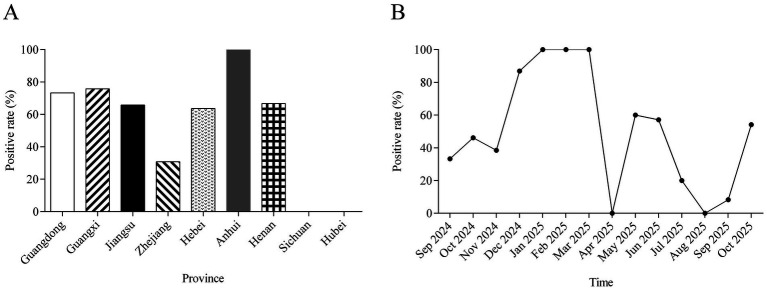 Figure 1
Figure 1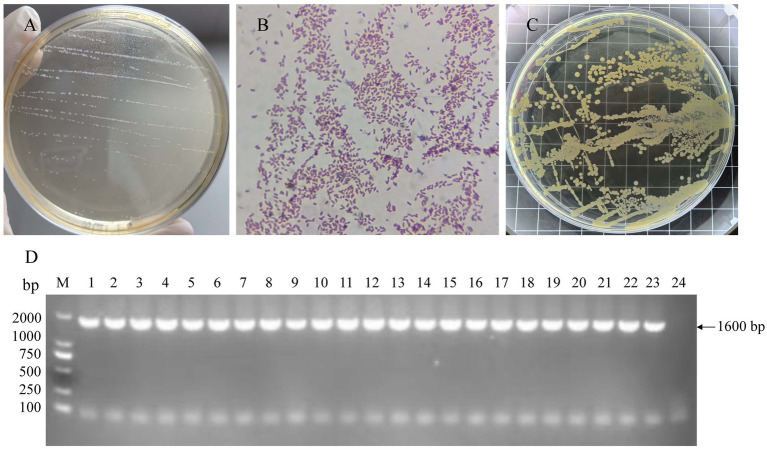 Figure 2
Figure 2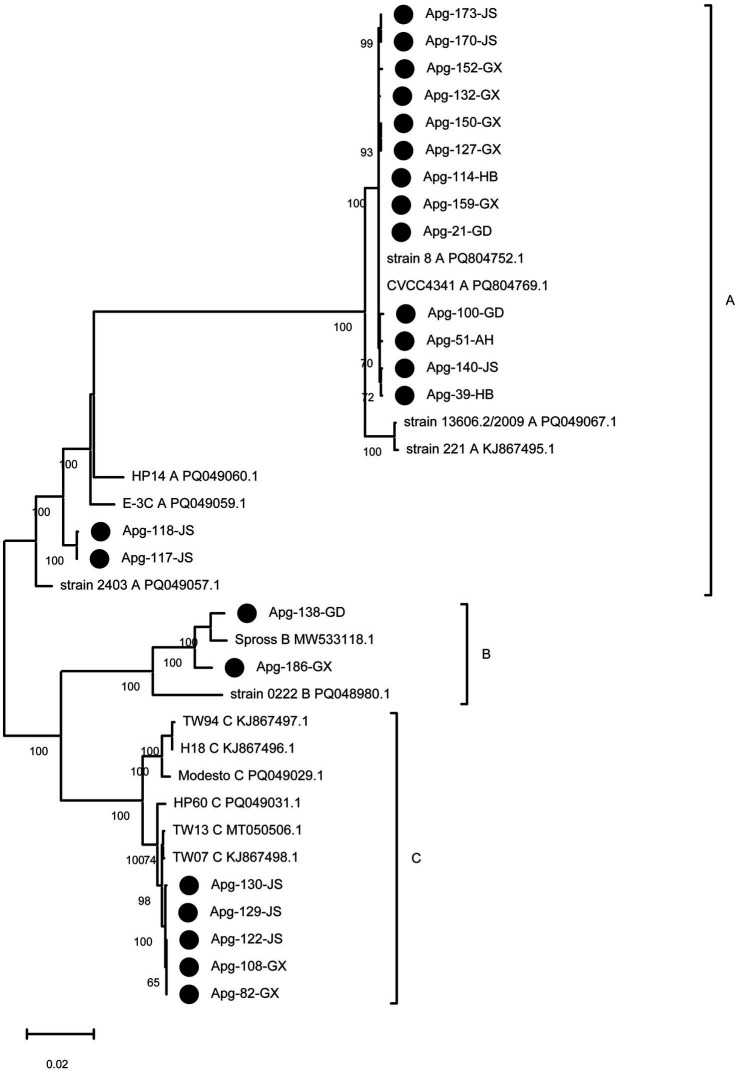 Figure 3
Figure 3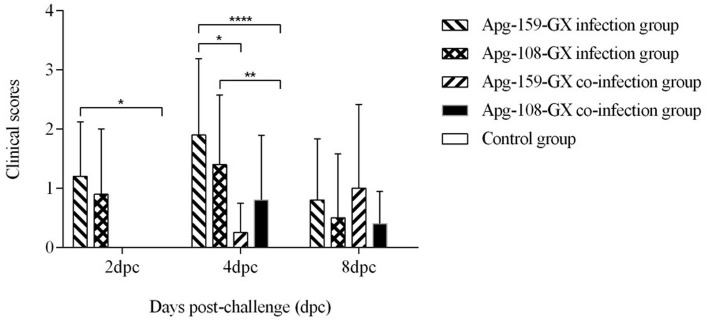 Figure 4
Figure 4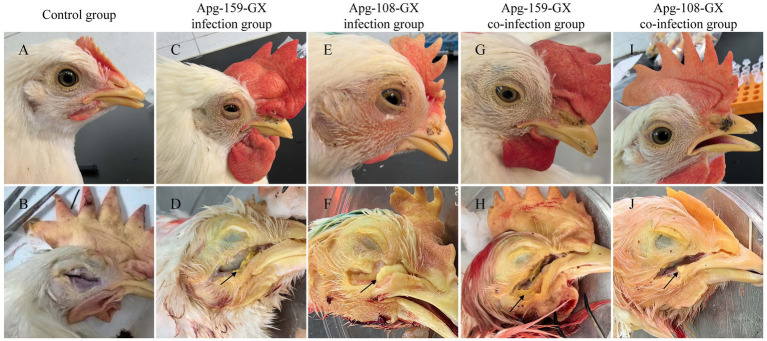 Figure 5
Figure 5| Province | Apg positive rate | Serotype A | Serotype B | Serotype C |
|---|---|---|---|---|
| Guangdong | 73.33% (11/15) | 8 | 1 | 2 |
| Guangxi | 75.81% (47/62) | 24 | 1 | 22 |
| Zhejiang | 30.77% (4/13) | 0 | 0 | 4 |
| Jiangsu | 65.82% (52/79) | 29 | 0 | 23 |
| Sichuan | 0% (0/7) | 0 | 0 | 0 |
| Hubei | 0% (0/6) | 0 | 0 | 0 |
| Hebei | 63.64% (7/11) | 7 | 0 | 0 |
| Anhui | 100% (4/4) | 4 | 0 | 0 |
| Henan | 66.67% (2/3) | 2 | 0 | 0 |
| Apg strain | Accession no. | Country | Serotype | Year | Note |
|---|---|---|---|---|---|
| 221 | Japan | A | Unknown | Reference strains | |
| 13606.2/2009 | USA | A | 2009 | ||
| 8 | China | A | 2011 | ||
| CVCC4341 | China | A | 2020 | ||
| 2,403 | USA | A | 2024 | ||
| E-3C | Brazil | A | Unknown | ||
| HP-14 | USA | A | 2024 | ||
| Spross | Unknown | B | Unknown | ||
| 0222 | USA | B | 2024 | ||
| strain H18 | Japan | C | Unknown | ||
| TW94 | Taiwan | C | 1994 | ||
| Modesto | USA | C | 1999 | ||
| TW07 | Taiwan | C | 2007 | ||
| TW13 | Taiwan | C | 2013 | ||
| HP60 | USA | C | 2024 | ||
| Apg-21-GD | PX735107 | China | A | 2024 | This study isolates |
| Apg-39-HB | PX735108 | China | A | 2024 | |
| Apg-51-AH | PX735109 | China | A | 2024 | |
| Apg-82-GX | PX735110 | China | C | 2024 | |
| Apg-100-GD | PX735111 | China | A | 2024 | |
| Apg-108-GX | PX735112 | China | C | 2024 | |
| Apg-114-HB | PX735113 | China | A | 2024 | |
| Apg-117-JS | PX735114 | China | A | 2025 | |
| Apg-118-JS | PX735115 | China | A | 2025 | |
| Apg-122-JS | PX735116 | China | C | 2025 | |
| Apg-127-GX | PX735117 | China | A | 2025 | |
| Apg-129-JS | PX735118 | China | C | 2025 | |
| Apg-130-JS | PX735119 | China | C | 2025 | |
| Apg-132-GX | PX735120 | China | A | 2025 | |
| Apg-138-GD | PX735121 | China | B | 2025 | |
| Apg-140-JS | PX735122 | China | A | 2025 | |
| Apg-150-GX | PX735123 | China | A | 2025 | |
| Apg-152-GX | PX735124 | China | A | 2025 | |
| Apg-159-GX | PX735125 | China | A | 2025 | |
| Apg-170-JS | PX735126 | China | A | 2025 | |
| Apg-173-JS | PX735127 | China | A | 2025 | |
| Apg-186-GX | PX735128 | China | B | 2025 |
- —Fourth Round of Guangdong Provincial Modern Agricultural Industry Technology System Innovation Team Construction Project
- —Science and Technology Plan Program of Guangdong Province
- —Science and Technology Plan Program of Yunfu city
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial infections and disease research · Aquaculture disease management and microbiota · Coccidia and coccidiosis research
Introduction
1
Infectious coryza (IC) is an acute, highly contagious respiratory disease of chickens that poses a significant threat to global poultry production. The causative agent, Avibacterium paragallinarum (A. paragallinarum), is a Gram-negative, capsulated, nicotinamide adenine dinucleotide (NAD)-dependent bacterium of the family Pasteurellaceae (1, 2). Clinically, the disease is characterized by sneezing, facial swelling, lacrimation, conjunctivitis, and serous-to-mucopurulent nasal discharge (1, 3, 4). While chickens of all ages are susceptible, the economic impact is most pronounced in commercial settings (1, 5). Broilers suffer from reduced feed intake and growth retardation, while layers can experience egg production drops of up to 40% (1). Environmental stressors and concurrent infections often exacerbate clinical signs and severity, contributing to widespread flock outbreaks (6, 7).
Since its initial description in 1931, A. paragallinarum has been identified worldwide (1, 8). In China, Infectious coryza was first documented in Beijing and subsequently detected across numerous provinces, including Jiangsu, Hebei, Anhui, and Ningxia (4). The capsular polysaccharide is a primary virulence factor and the key protective antigen, making it a major focus for vaccine development (9, 10). The classical Page serotyping scheme based on capsular antigenicity classifies A. paragallinarum into three serovars (A, B, and C) without cross-protect (11). This scheme was later refined by Kume, who defined serogroups I, II, and III as equivalents to Page’s serovars A, B, and C, respectively (12). To date, nine recognized serovars have been identified (A-1 to A-4, B-1, and C-1 to C-4) (12, 13).
Levels of cross-protection within serovars vary considerably. Serovar A strains generally induce strong mutual protection, serovar B provides only partial protection, and serovar C offers minimal cross-protection. Furthermore, some field isolates remain untypable (1, 14). All three major serovars have been identified in China, with recent epidemiological data revealing an increasing incidence of IC. The current landscape is characterized by the predominance of serovars A and C, alongside the occasional co-circulation of all three types (3, 8, 15).
Accurate serotyping is essential for epidemiological investigation and vaccine selection. However, conventional haemagglutination-inhibition (HI) assays, which rely on high-quality standard antisera, are hampered by challenges in reagent availability and standardization, often yielding ambiguous results (14). To overcome these limitations, molecular serotyping based on sequence polymorphisms in the hmtp210 gene has been established as a reliable alternative. Previous reports indicated that variability within the N-terminal 1–400 amino acid region of hmtp210 protein accurately differentiates Page serovars A, B, and C (14). The discriminatory power was further enhanced by Li et al. (16), who showed that analysis of both the 1–400 and 1,100–1,600 regions provide superior resolution. Consequently, sequencing of hmtp210 gene has become the molecular gold standard for serotyping A. paragallinarum.
Given the substantial economic impact of IC in China and the dynamic, regionally variable distribution of A. paragallinarum serovars, continuous and targeted epidemiological surveillance is crucial. This study aimed to characterize the contemporary epidemiological profile of A. paragallinarum in China. Clinical samples were screened for A. paragallinarum, and confirmed by species-specific PCR, and subsequently molecularly serotyped based on hmtp210 gene sequencing. Furthermore, the pathogenicity of representative strains from the predominant serotypes was assessed. The findings provide updated data on the geographical distribution, molecular characteristics, and virulence of circulating strains. These insights are critical for informing evidence-based prevention strategies and guiding the development of effective, regionally tailored vaccines.
Materials and methods
2
Ethics statement
2.1
All animal procedures were reviewed and approved by the Animal Care Committee of South China Agricultural University (approval ID: SYXK-2024-0136). All procedures complied with the Guidelines for the Care and Use of Laboratory Animals issued by the Ministry of Science and Technology of China. To minimize distress, birds were euthanized following deep isoflurane anesthesia and subsequent intravenous pentobarbitone administration, according to established broiler anesthesia protocols (17).
Sample collection
2.2
A systematic epidemiological investigation of IC was conducted from September 2024 to October 2025 across nine provinces in China (Guangdong, Guangxi, Jiangsu, Zhejiang, Sichuan, Hubei, Hebei, Henan, and Anhui). Swabs from the infraorbital sinuses and/or nasal exudates were collected from chickens exhibiting characteristic clinical signs of IC, including facial swelling and nasal discharge. Samples were transported under cold-chain conditions and refrigerated at 4 °C until processing. A total of 200 samples were obtained, and their geographic distribution is detailed in Table 1. The number of samples obtained from each province was determined by the availability of clinically suspected IC cases and voluntary diagnostic submissions during the study period, rather than by a predefined or evenly stratified sampling design. Consequently, sample sizes varied substantially among provinces.
Isolation and identification of Avibacterium paragallinarum
2.3
Following surface disinfection of the head, infraorbital sinuses were aseptically incised. Exudates were collected using sterile inoculating loops and streaked onto tryptic soy agar (TSA) supplemented with 10% heat-inactivated chicken serum and 0.02% β-nicotinamide adenine dinucleotide (β-NAD). Plates were incubated at 37 °C in a 10% CO_2_ atmosphere for 24 h. Presumptive colonies exhibiting smooth, dew-drop-like morphology were subcultured in tryptic soy broth (TSB) containing 10% chicken serum and 0.01% NAD, and incubated at 37 °C for 6 h with shaking (180 rpm). For purification, 3–5 single colonies were streaked to obtain pure cultures. Colony and cellular morphology were examined visually and via Gram staining and microscopy, respectively. Species-level identification was confirmed by PCR amplification of an ~1,600 bp fragment from the hmtp210 gene using established primers (18). Isolates yielding the expected amplicon were identified as A. paragallinarum. A previously characterized A. paragallinarum reference strain (CVCC4341) was included as a positive control in each PCR run to verify assay specificity and performance, and its identity had been confirmed by sequencing. Due to limitations related to sample quality, transport conditions, and biosafety requirements, bacterial isolation was attempted only for a subset of PCR-positive samples with sufficient volume and appropriate storage conditions.
PCR amplification and phylogenetic analysis
2.4
Bacterial genomic DNA was extracted exclusively from purified A. paragallinarum colonies obtained as described in this study. DNA extraction was conducted using the FastPure Microbiome DNA Isolation Kit (Cat. no. DC502; Vazyme Biotech, Nanjing, China) according to the manufacturer’s instructions (Version 24.1). Briefly, approximately 400 μL of bacterial culture was transferred to a lysis tube and centrifuged at 10,000 rpm for 3 min to pellet the cells. After supernatant removal, the pellet was resuspended and incubated at 70 °C for 5 min to ensure efficient cell lysis. Following centrifugation (10,000 rpm, 1 min), the lysate was mixed with absolute ethanol and loaded onto a silica-based DNA binding column. The column was washed sequentially with Buffer WP and Buffer WB, dried by high-speed centrifugation, and genomic DNA was eluted in 50 μL of elution buffer. DNA concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA), and extracted DNA was stored at −20 °C until further use.
The hmtp210 gene was amplified using specific primers: HMTp210F (5’-ATGAATAAAGTTTTTAAAATTAAATATTCTGTTATTAAACAAGAAAT-3′) and HMTp210R (5’-CTAAAAGGTAAATCCAACACTCATTGCTGCCC-3′), which generate an approximately 6100 bp amplicon (19). PCR reactions (50 μL) consisted of 25 μL PrimeSTAR HS Premix (Takara Bio), 10 pM of each primer, and 2 μL of template DNA. Reactions were carried out on a T100 Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, United States) under the following cycling conditions: an initial denaturation at 98 °C for 3 min; 32 cycles of denaturation at 98 °C for 15 s, annealing at 55 °C for 10 s, and extension at 72 °C for 35 s; followed by a final extension at 72 °C for 5 min. PCR products were held at 4 °C until further analysis. Amplified PCR products were separated by 1% agarose gel electrophoresis. Target bands were excised and purified using a gel extraction kit (Solarbio). Purified amplicons were A-tailed using a DNA A-Tailing Kit (Takara Bio), ligated into the pMD19-T vector (Takara Bio), and transformed into Escherichia coli DH5α competent cells. For each isolate, three independent positive clones were selected for bidirectional sequencing by Sangon Biotech (Shanghai, China). Sequences were assembled and analyzed using the SeqMan program of DNASTAR Lasergene 7.1 software. The hmtp210 sequences from the 22 obtained isolates were aligned with those from 15 reference strains (Table 2) using the ClustalW algorithm in MEGA11. A phylogenetic tree was constructed via the neighbor-joining method with 1,000 bootstrap replicates to assess genetic relationships and molecular serotype classification.
Animal experiment
2.5
The serotype A isolate Apg-159-GX and the serotype C isolate Apg-108-GX used in this study were clinical isolates obtained in the present investigation and were selected from the 22 confirmed Avibacterium paragallinarum isolates for challenge experiments. To assess the pathogenicity and transmissibility, an experimental infection study was performed using 50-day-old specific pathogen-free (SPF) chickens. Birds were randomly assigned into three groups: Group 1 (n = 15) was inoculated with serotype A isolate Apg-159-GX, Group 2 (n = 15) with serotype C isolate Apg-108-GX, and Group 3 (n = 15) served as the unchallenged control. For Groups 1 and 2, ten birds per group were directly challenged via inoculation into the left infraorbital sinus with 0.2 mL of a bacterial suspension containing 1 × 10^4^ CFU/mL. The remaining five birds per group were housed as in-contact sentinels to assess horizontal transmission. Control group birds received 0.2 mL of phosphate-buffered saline (PBS) via the same route. The challenge dose was selected based on previously published experimental infection models of infectious coryza, in which an inoculum of approximately 10^4^ CFU was demonstrated to reliably induce typical clinical signs without causing excessive mortality (20).
All chickens were monitored daily for 8 days post-challenge (dpc). Clinical signs were evaluated using a standardized clinical scoring system adapted from a previously published method for infectious coryza, with minor modifications to better reflect the severity of infraorbital sinus involvement observed under the present experimental conditions (8). Briefly, clinical signs were scored as follows: 0 = no signs; 1 = mild facial swelling/serous exudate; 2 = moderate swelling with mucous nasal discharge; 3 = severe swelling with copious exudate and/or transient blindness. At 8 dpc, all surviving birds were humanely euthanized. Necropsy was performed, with a specific focus on the infraorbital sinuses to confirm the presence and severity of caseous exudate characteristic of infectious coryza.
Statistical analysis
2.6
Statistical analyses were conducted using GraphPad Prism 9 (GraphPad Software Inc., San Diego, United States). Differences in clinical scores between groups across time points were analyzed using two-way analysis of variance (ANOVA). A p-value < 0.05 was considered statistically significant.
Results
3
Spatiotemporal distribution and serotype diversity of Avibacterium paragallinarum
3.1
A total of 200 clinical samples collected from nine major poultry-producing provinces were analyzed to determine the geographic distribution, seasonal patterns, and serotype composition of A. paragallinarum. A. paragallinarum detection varied substantially among provinces (Figure 1A; Table 1). The highest positivity rates were observed in Anhui (100.0%, 4/4), followed by Guangxi (75.8%, 47/62) and Guangdong (73.3%, 11/15). Moderate detection rates were recorded in Jiangsu (65.8%, 52/79), Henan (66.7%, 2/3), and Hebei (63.6%, 7/11), while a relatively low rate was found in Zhejiang (30.8%, 4/13). All samples from Sichuan (0/7) and Hubei (0/6) tested negative, revealing a pronounced regional heterogeneity with circulation concentrated primarily in southern and eastern China. These provincial detection rates should be interpreted with caution, particularly for regions with limited sample numbers, as the present study was not designed to estimate province-level prevalence.
Spatiotemporal distribution of A. paragallinarum in China. (A) Geographic distribution of A. paragallinarum positivity across nine provinces. (B) Monthly positivity rates of A. paragallinarum.
Serotyping identified clear geographic differences in serotype distribution (Table 1). Serotype A was the most widespread, detected in Jiangsu (29 positive samples), Guangxi (24 positive samples), Guangdong (8 positive samples), Hebei (7 positive samples), Anhui (4 positive samples), and Henan (2 positive samples). In contrast, serotype B was identified in only one sample from Guangxi and one from Guangdong. Serotype C exhibited regional clustering, predominating in Jiangsu (23 positive samples) and Guangxi (22 positive samples), with lower occurrence in Zhejiang (4 positive samples) and Guangdong (2 positive samples). Serotype C was absent from Sichuan, Hubei, Hebei, Anhui, Henan. These results indicate co-circulation of serotypes A and C in key provinces (e.g., Jiangsu, Guangxi, Guangdong), a more restricted distribution of serotype A in northern/central regions, and an apparent absence of serotype B during the study period.
Seasonal dynamics revealed that a clear seasonal pattern was observed (Figure 1B). The positive rate increased from 33.3% in September 2024 to a peak of 87.0% in December 2024. A sustained high-incidence phase occurred from January to March 2025 (100.0%), followed by a sharp decline to 0% in April 2025. Positivity remained low throughout May–September. A distinct resurgence was observed in October 2025 (54.2%), establishing a stable, annual epidemic cycle with peak activity during late autumn and winter.
Isolation and identification of Avibacterium paragallinarum
3.2
A. paragallinarum was isolated from infraorbital sinus exudates collected from clinically suspected IC cases. From the 200 field samples, 22 isolates with phenotypic characteristics consistent with A. paragallinarum were recovered. Although a substantially larger number of samples tested positive by PCR, successful bacterial isolation was limited to these 22 isolates. On TSA supplemented with 10% chicken serum and NAD, colonies appeared as grey-white, round, smooth, and dew-drop-like (Figure 2A). Microscopic examination after Gram staining confirmed Gram-negative coccobacilli (Figure 2B). All isolates exhibited the characteristic satellite growth around Staphylococcus aureus streaks, confirming their NAD dependency (Figure 2C). To ensure clonal purity, isolates were subcultured, and 3–5 well-isolated colonies from each were subjected to species-specific PCR targeting the hmtp210 gene. All selected colonies yielded the expected ~1,600 bp amplicon (Figure 2D), confirming their identity as A. paragallinarum and verifying the successful isolation of genetically representative strains.
Isolation and characterization of A. paragallinarum field isolates. (A) Characteristic grey-white, dew-drop-like colonies of A. paragallinarum on supplemented TSA. (B) Gram-negative coccobacilli morphology of A. paragallinarum isolates. (C) Satellite growth around Staphylococcus aureus. (D) PCR amplification of the hmtp210 gene. M, DL2000 DNA marker; lanes 1–22 correspond to PCR-positive clinical isolates confirmed in this study, and lane 23 represents the positive control (A. paragallinarum reference strain [CVCC4341]); lane 24, negative control.
Phylogenetic analysis of the hmtp210 gene
3.3
A phylogenetic tree was constructed based on the hmtp210 gene sequences from the 22 isolates and 15 reference strains representing serotypes A, B, and C. The GenBank accession numbers of the clinical isolates and reference strains are listed separately in Table 2. The resulting phylogeny (Figure 3) revealed that all isolates clustered into three distinct, well-supported clades corresponding to serotypes A, B and C, demonstrating strong concordance between molecular genotyping and conventional serotyping.
Phylogenetic analysis based on the hmtp210 gene sequences. The phylogenetic tree was constructed using MEGA 7.0 based on hmtp210 sequences from 22 isolates obtained in this study and 15 reference strains. Black circles denote the filed strain isolated in this study. Bootstrap values were calculated with 1,000 replicates.
The majority of isolates (15 out of 22) formed a monophyletic clade with 7 reference serotype A strains, indicating a genetically homogeneous lineage circulating in the sampled regions. Two isolates clustered closely with a reference serotype B strain Spross. The remaining 5 isolates clustered with 2 reference serotype C strains (TW07, TW13), reflecting their close evolutionary relationship. These results confirm the predominance of serotype A among circulating strains in the sampled regions during the study period.
Clinical manifestations and pathogenicity of Avibacterium paragallinarum
3.4
Among the 22 confirmed A. paragallinarum isolates, Apg-159-GX (serotype A) and Apg-108-GX (serotype C) were selected for experimental challenge. Following direct infraorbital sinus inoculation of SPF chickens, both representative strains—Apg-159-GX (serotype A) and Apg-108-GX (serotype C)—induced typical infectious coryza. Clinical signs emerged at 2 dpc, peaked at 4 dpc, and largely resolved by 8 dpc. As shown in Figure 4, directly inoculated birds from both serotype groups exhibited significantly higher clinical scores than controls at 2 and 4 dpc (p < 0.05). At 4 dpc, clinical scores in the Apg-159-GX direct-challenge group were also significantly higher than in its contact-exposed group (p < 0.05), whereas no significant difference was observed between the Apg-108-GX challenge and contact groups (p > 0.05). By 8 dpc, clinical scores did not differ significantly among any groups (p > 0.05). No significant differences in mean clinical scores were observed between the two serotypes under direct-challenge conditions, indicating comparable overall clinical virulence in this model.
Clinical severity following experimental infection. Chickens were challenged with serotype A strain APG-159-GX or serotype C strain APG-108-GX via direct inoculation or contact exposure. Clinical signs were scored at 2, 4, and 8 days post-challenge (dpc).
Gross pathology. Gross lesion assessment corroborated the clinical findings (Figure 5). Control birds showed no abnormalities (Figures 5A,B). Directly challenged birds from both serotype groups developed classic IC lesions, including periocular swelling, conjunctivitis, and accumulation of yellow caseous exudate in the infraorbital sinus (Figures 5C–F). Contact-exposed birds displayed similar but milder lesions (Figures 5G–J), consistent with a lower infectious dose during natural transmission.
Gross pathology of SPF chickens infected with A. paragallinarum. Control chickens without clinical signs (A) and gross lesions in the infraorbital sinus (B). (C,D) Serotype A strain Apg-159-GX—partially closed eyes with conjunctivitis and moderate periocular swelling (C), and accumulation of yellow caseous exudate in the sinus (D). (E,F) Serotype C strain Apg-108-GX—nasal discharge, lacrimation, and moderate swelling (E), with the sinus filled with yellow caseous material (F). (G,H) Contact-exposed birds with small amounts of caseous exudate in Apg-159-GX inoculated group (H). (I,J) Contact-exposed birds with small amounts of caseous exudate in Apg-108-GX group.
Discussion
4
A. paragallinarum, the etiological agent of infectious coryza (IC), has re-emerged repeatedly in China over the past decade, imposing significant economic losses on the poultry industry. IC is a highly contagious respiratory disease characterized by conjunctivitis, sinusitis, nasal discharge, facial swelling, and depression (1). Beyond causing reduced feed intake and growth retardation in broilers, the disease can lead to egg production drops of up to 40% in laying flocks (1, 21). Furthermore, recovered birds often become chronic carriers, and subsequent environmental or infectious stressors can readily trigger recurrent outbreaks, substantially complicating long-term disease control efforts.
A. paragallinarum has a global distribution and is particularly prevalent in intensively managed poultry operations across Asia, the Americas, Europe, and Africa (6, 22, 23). In China, the first confirmed reports of serotypes A, C, and B date from 1986, 1993, and 2003, respectively (15). Epidemiological investigations have consistently identified serotypes A and C as the predominant circulating types, with serotype B detected only sporadically (3, 24). Surveys conducted from 2019–2020 confirmed the co-circulation of all three serotypes, with a notable predominance of serotype C during that period (4). The epidemiology of A. paragallinarum in China exhibits distinct winter–spring seasonality and higher prevalence in southern provinces compared to northern regions (25, 26). Collectively, these findings underscore the dynamic and evolving nature of A. paragallinarum epidemiology in China, emphasizing the critical need for continuous surveillance.
This multi-provincial survey provides updated data on the spatial and temporal patterns of A. paragallinarum circulation in China. IC occurred year-round but exhibited strong seasonality, with distinct peaks in winter and spring—consistent with prior observations that cool, humid conditions favor bacterial survival and transmission (25). Positive samples were mainly concentrated in Jiangsu, Guangxi, and Guangdong, while Sichuan, Zhejiang, and Hubei contributed only sporadic detections. The higher prevalence in southern coastal provinces likely reflects intensive poultry production, frequent bird movements, and climatic conditions that favor pathogen persistence. Serotype A was predominated in Jiangsu and Guangdong, while both serotypes A and C co-circulated in Guangxi. Although serotype B and serotype C were detected at low frequency, their continued presence suggests ongoing, albeit limited, transmission cycles. These epidemiological patterns reaffirm the necessity for region-specific surveillance programs and evidence-based vaccine selection.
A notable observation in this study was the discrepancy between the number of PCR-positive samples and the relatively small number of successfully isolated strains. This outcome is expected and reflects fundamental methodological differences between molecular detection and bacterial isolation. PCR assays detect bacterial DNA regardless of organism viability, whereas successful isolation of A. paragallinarum is strongly influenced by sample freshness, transport conditions, prior antimicrobial treatment, and the organism’s fastidious growth requirements. Similar discrepancies between PCR detection rates and isolation success have been reported previously (24). Importantly, this limitation does not undermine the epidemiological significance of PCR-based surveillance but highlights the need for cautious interpretation when correlating molecular positivity with culture recovery.
Our findings on pathogenicity and transmission dynamics align closely with previous studies. Zhao et al. reported that clinical signs appeared earlier following direct infraorbital sinus inoculation than after natural contact exposure, with peak signs in the latter group delayed by 3–5 days (26). This pattern is consistent with the 4-day delay we observed in contact-sentinel birds compared to directly inoculated chickens. Similarly, Guo et al. documented rapid development of inflammatory lesions in the sinus and tracheal mucosa as early as 3 days post-infection (24). Earlier research demonstrated that A. paragallinarum adheres tightly to ciliated epithelial surfaces, inducing epithelial desquamation and airway narrowing (24, 27).
The extensive tissue destruction observed in this study is likely attributable, in part, to toxin-mediated injury. Several studies have implicated RTX (Repeats-in-ToXin) family toxins in the pathogenesis of A. paragallinarum (27). These toxins form pores in host cell membranes, leading to osmotic lysis, ionic dysregulation, and necrotic cell death (28, 29). Putative RTX-encoding genes have been identified in the genome of A. paragallinarum, and RTX1-like proteins have been characterized in related Avibacterium species (28, 29). Furthermore, Gram-negative outer membrane vesicles (OMVs) contribute to virulence by facilitating biofilm formation, stimulating potent pro-inflammatory responses, and delivering cytotoxic cargo to host tissues (30). It is plausible that OMVs derived from A. paragallinarum function similarly in vivo.
Host species and age are critical determinants of A. paragallinarum pathogenicity. Although cross-species transmission can occur, field isolates demonstrate strong host preference, with chicken- and goose-origin strains exhibiting high virulence in their natural hosts but attenuated effects in heterologous species (23). Balouria et al. reported pronounced differences in clinical presentation between chickens and Japanese quail, further supporting this host-restricted virulence (23). Age-related susceptibility is another key factor. In our study, 50-day-old SPF chickens developed clinical signs at 24 hpc. In contrast, younger birds (2.5 weeks old) in Balouria’s work exhibited symptoms within 12 h (23), suggesting that juvenile birds may experience a more rapid and potentially more severe disease onset.
These observations reinforce that clinical outcome reflects the combined effects of strain virulence, host susceptibility, and environmental conditions. Efficient horizontal transmission observed for both serotypes further underscores the potential for rapid flock-to-flock spread under commercial conditions.
In conclusion, IC remains a significant challenge for Chinese poultry industry. A. paragallinarum is widely distributed, with serotypes A and C currently predominating and exhibiting high virulence and transmissibility. Although serotype B circulates sporadically, its persistence necessitates ongoing surveillance. Effective control of IC requires a multifaceted strategy combining regionally tailored vaccination—based on prevalent serotypes—with stringent biosecurity measures to interrupt transmission. Sustained epidemiological monitoring is essential to track serotype dynamics, inform vaccine updates, and enable the development of targeted, effective prevention strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1El-Gazzar M Gallardo R Bragg R Hashish A Sun HL Davison S . Avibacterium paragallinarum, the causative agent of infectious coryza: a comprehensive review. Avian Dis. (2025) 68:362–79. doi: 10.1637/aviandiseases-D-24-00105, 40249575 · doi ↗ · pubmed ↗
- 2Gallardo RA Da SA Egana-Labrin S Stoute S Kern C Zhou H . Infectious coryza: persistence, genotyping, and vaccine testing. Avian Dis. (2020) 64:157–65. doi: 10.1637/0005-2086-64.2.15732550616 · doi ↗ · pubmed ↗
- 3Xu Y Cheng J Huang X Xu M Feng J Liu C . Characterization of emergent Avibacterium paragallinarum strains and the protection conferred by infectious coryza vaccines against them in China. Poult Sci. (2019) 98:6463–71. doi: 10.3382/ps/pez 531, 31801310 PMC 8913951 · doi ↗ · pubmed ↗
- 4Guo M Chen X Zhang H Liu D Wu Y Zhang X. Isolation, serovar identification, and antimicrobial susceptibility of Avibacterium paragallinarum from chickens in China from 2019 to 2020. Vet Sci. (2022) 9:27. doi: 10.3390/vetsci 9010027 PMC 878176735051111 · doi ↗ · pubmed ↗
- 5Crispo M Sentíes-CuéCG Cooper GL Mountainspring G Corsiglia C Bickford AA . Otitis and meningoencephalitis associated with infectious coryza (Avibacterium paragallinarum) in commercial broiler chickens. J Vet Diagn Invest. (2018) 30:784–8. doi: 10.1177/1040638718792964, 30129392 PMC 6505790 · doi ↗ · pubmed ↗
- 6Shelkamy M Hashish A Srednik ME Chaves M Macedo NR Gadu E . Prevalence of nonpathogenic avibacterium paragallinarum in naive-healthy layer flocks across multiple states in the United States. Transbound Emerg Dis. (2025) 2025. doi: 10.1155/tbed/9994679, 40420864 PMC 12103973 · doi ↗ · pubmed ↗
- 7Morales-Erasto V Falconi-Agapito F Luna-Galaz GA Saravia LE Montalvan-Avalos A Soriano-Vargas EE . Coinfection of Avibacterium paragallinarum and Ornithobacterium rhinotracheale in chickens from Peru. Avian Dis. (2016) 60:75–8. doi: 10.1637/11265-082015-Res Note.1, 26953948 · doi ↗ · pubmed ↗
- 8Cui W Wang C Wu Y Wang T Wang Z Zhang W . Epidemiological investigation of infectious coryza in Central China and the effect of enterococcus faecium on improving vaccine immunity. Poult Sci. (2025) 104:105622. doi: 10.1016/j.psj.2025.105622, 40769017 PMC 12540443 · doi ↗ · pubmed ↗