Modeling the potential distribution of Wesselsbron, Sindbis, and Middelburg viruses and their vectors in Africa under future climatic and land-use changes
Maureen Nabatanzi, Selina L. Graff, Kigai E. E. Bigala, Peter Z. Sabakaki, Teddy A. Tindyebwa, Julius J. Lutwama, Innocent B. Rwego, Anthony M. Nsubuga, Sandra Junglen, Lisa Biber-Freudenberger

TL;DR
This study maps the future spread of three African arboviruses and their mosquitoes under climate and land-use changes, highlighting risks for human and animal outbreaks.
Contribution
The study provides novel ecological risk maps for Wesselsbron, Sindbis, and Middelburg viruses and their vectors under future climate and land-use scenarios in Africa.
Findings
High ecological suitability for five mosquito vectors is predicted in Equatorial and Southern Africa, with future hotspots emerging in Southern and Eastern regions.
Changing precipitation patterns, urbanization, and climate are key drivers of vector and arbovirus geographic expansion.
Ecological risk for arbovirus presence may shift from rural to urban areas with future ecological changes.
Abstract
Outbreaks of zoonotic arboviruses originating in Africa occur amidst complex ecological changes and are increasingly emerging as important neglected tropical diseases. Despite sporadic epizootics and human cases of Wesselsbron virus (WSLV), Sindbis virus (SINV), and Middelburg virus (MIDV) in Africa, knowledge of associated risks remains insufficient for prevention. We developed species distribution models for the three viruses alongside five key vectors, Aedes circumluteolus and Aedes mcintoshi for WSLV; Culex univittatus and Culex pipiens for SINV; and Mansonia africana and Aedes mcintoshi for MIDV that indicate areas with ecological suitability for the arboviruses in Africa. We integrated virus and mosquito species occurrence data with climate and land-use data for current (2015) and future (2021 – 2040) scenarios under two shared socioeconomic pathways of emission and climate…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
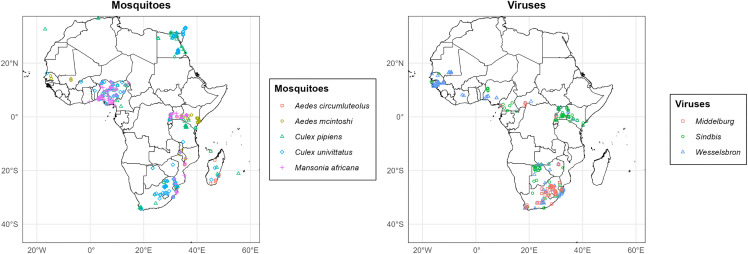 Fig 1
Fig 1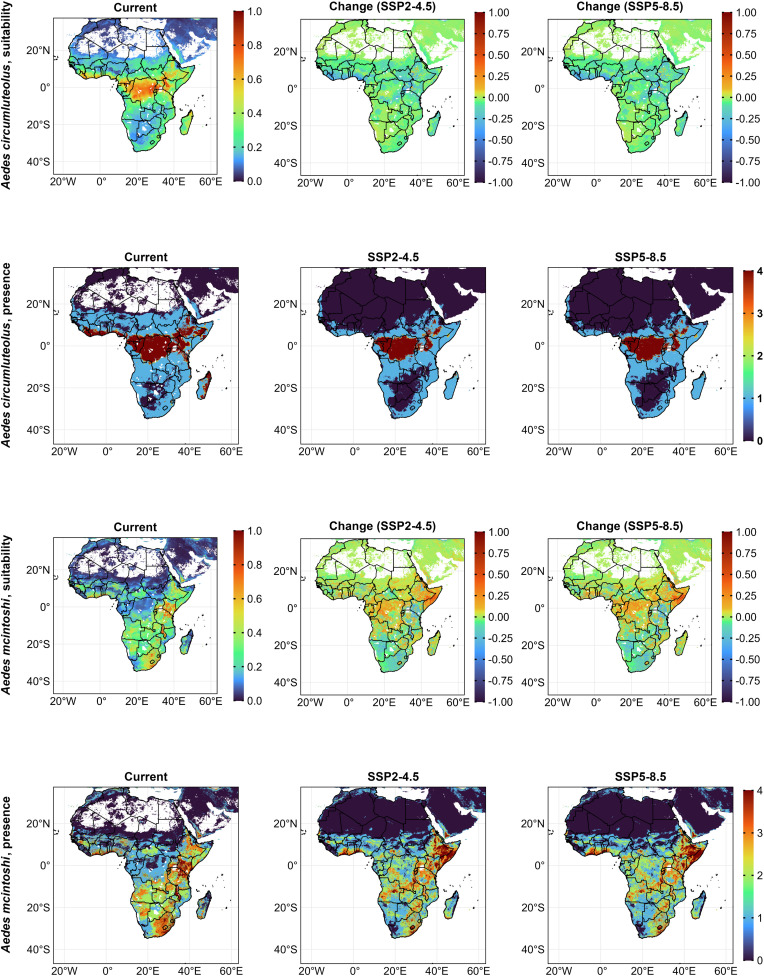 Fig 2
Fig 2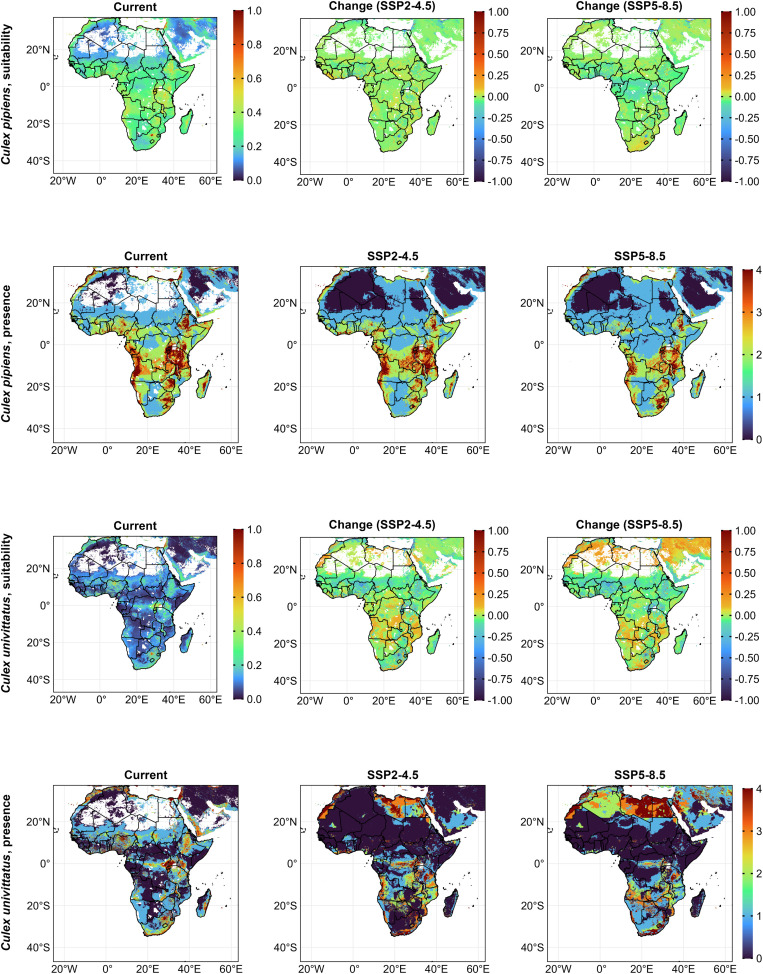 Fig 3
Fig 3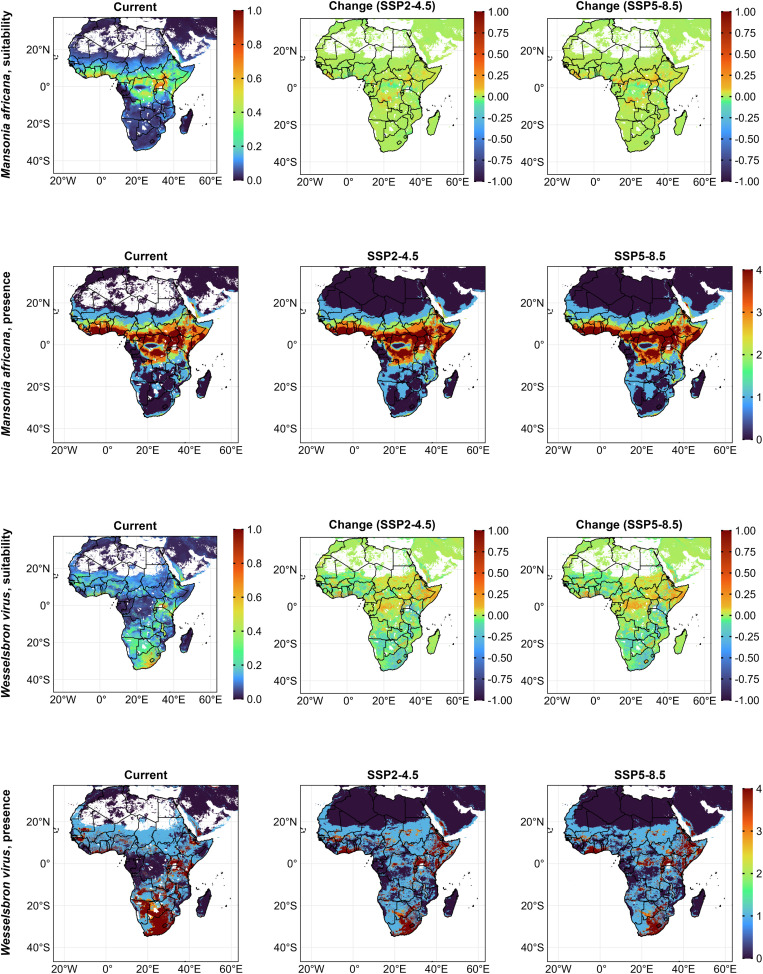 Fig 4
Fig 4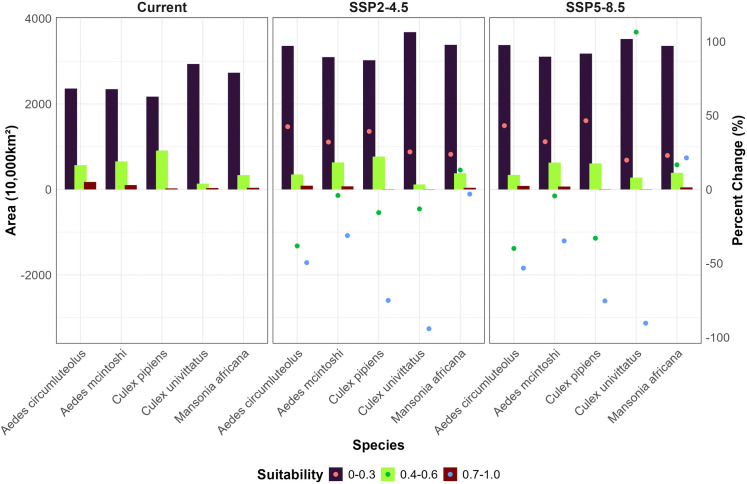 Fig 5
Fig 5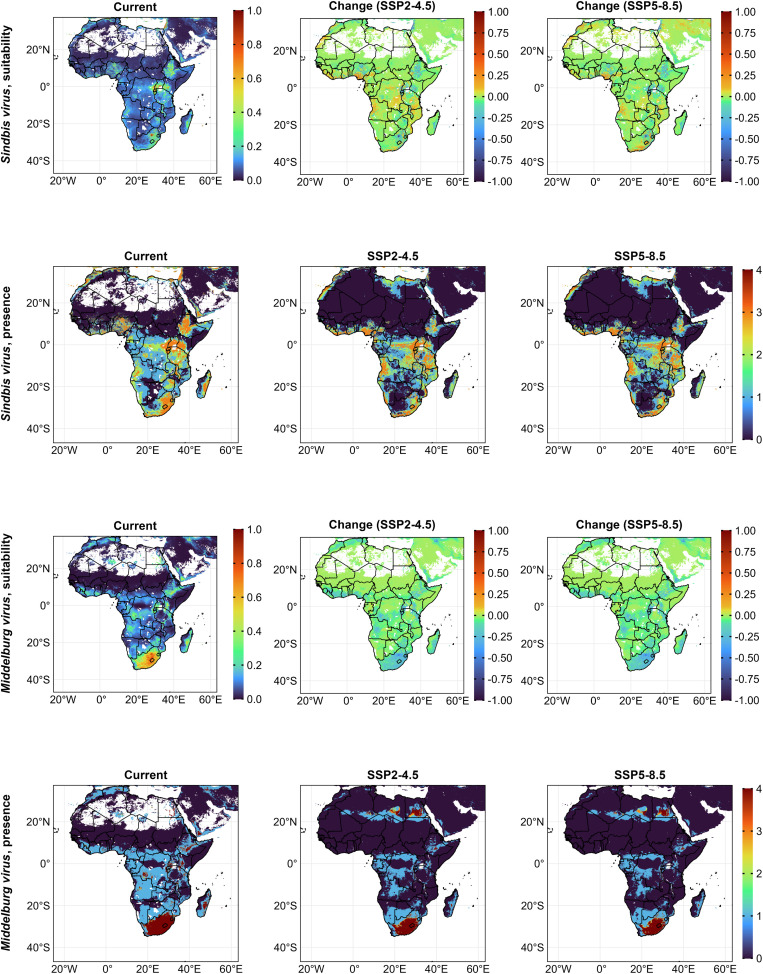 Fig 6
Fig 6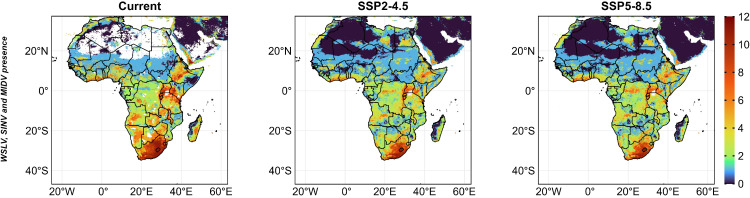 Fig 7
Fig 7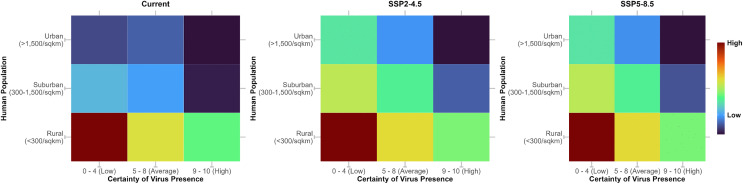 Fig 8
Fig 8- —http://dx.doi.org/10.13039/501100001659Deutsche Forschungsgemeinschaft
- —Collaborative Research Centre
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Zoonotic diseases and public health · Species Distribution and Climate Change
Introduction
Arthropod-borne infections account for almost half of the recent emerging infectious diseases, particularly in countries of the global south [1]. Wesselsbron virus (WSLV), belonging to the family Flaviviridae and genus Flavivirus, was first identified in 1955 in South Africa [2]. Human disease is characterized by fever, headache, arthralgia, and myalgia [3]. The reported geographical distribution is primarily in Africa and Thailand [4], with sporadic human cases and antibodies to WSLV detected in South Africa, Uganda, Nigeria, Mauritania, and Senegal [3,5–8]. WSLV has been detected from multiple mosquito taxa, including several Aedes (Ae), Anopheles, Culex (Cx), and Mansonia (Ma) species, but its main vectors are freshwater breeding Aedes species, notably, Ae. circumluteolus, Ae. mcintoshi and Ae. caballus [4,9,10]. In Africa, WSLV poses a substantial zoonotic threat to humans, exemplified by its broad geographical circulation in mosquitoes, amplification in multiple vertebrates such as rodents, other wild animals, and epizootic potential in livestock [5,11–13]. Domestic sheep are highly susceptible (abortions, neonatal mortality), with cattle and goats also acting as amplifying hosts, while humans are incidental hosts [9,14].
Sindbis virus (SINV), an alphavirus (family Togaviridae) which was first isolated in Egypt in 1952, is widely distributed across Africa, Europe, the Middle East, and parts of Asia [15–17]. In humans, SINV infections are characterized by febrile illness, widespread maculopapular rash, myalgia, and prolonged arthralgia [18,19]. In northern Europe, SINV is endemic in Sweden and Finland, causing thousands of symptomatic human cases during peak years; persistent low-level circulation is linked to resident bird populations and Culex vectors [19,20]. In South Africa, epidemics associated with Cx. univittatus have been documented for decades, with large outbreaks recorded in the 1970s – 80s affecting thousands of humans and sporadic activity continuing thereafter [21,22]. Sporadic human cases were reported in Uganda, China, Australia, and Spain [15,18,23]. Human infections arise from bites by infectious mosquitoes, an effect of spill-overs from enzootic circulation in birds [24]. Transmission events are dominated by multiple ornithophilic Culex mosquitoes, notably, Cx. univittattus and Cx. pipiens in Africa and Cx. torrentium/pipiens in Europe where Ae. cinereus is a bridge vector [4,18,22,24,25]. The enzootic cycle relies on avian amplifying hosts such as thrushes (Turdus spp.) and crows (Corvus corone sardonius), implicated as reservoirs [18,24,26]. Cross-species spill-over is likely enhanced by a broad vertebrate host range, including multiple wild and domestic animals like horses [15,16,27].
Middelburg virus (MIDV) of the family Togaviridae, genus Alphavirus was first isolated in South Africa in 1957. The virus is thought to circulate primarily in enzootic transmission cycles involving mosquitoes and vertebrate hosts, with episodic spill-over into humans and domestic animals [28–30]. In a South African study, Fourie et al (2022) suggested that humans infected with MIDV disease could present with fever and neurologic symptoms [31]. Overall, formal vector competence and vertical transmission studies for MIDV remain scarce. Recent entomological investigations in South Africa detected MIDV in several mosquito genera, including Aedes and Culex, supporting the likelihood of a multi-vector system [32]. In Uganda, MIDV was detected in Mansonia africana and Eretmapodites intermedius, suggesting possible enzootic transmission in forested ecosystems [33]. Floodwater Aedes species, particularly Aedes mcintoshi, have been repeatedly highlighted as ecologically important due to their association with rainfall-driven emergence, potential involvement in vertical transmission, and frequent detection in arbovirus surveillance studies [9,28]. Serological and virologic evidence indicate MIDV exposure in a range of wild and domestic animals, including livestock, implicating mammals as amplifying hosts during periods of increased vector abundance [27]. MIDV has demonstrated epizootic potential, especially among sheep, and has been associated with severe neurological disease in horses, livestock, and wildlife, indicating its pathogenicity in mammals [27,29,34].
Anthropogenic environmental change and climate variability increasingly influence the epidemiology of arboviral diseases by reshaping the distribution of mosquito vectors, vertebrate hosts, and transmission dynamics [35,36]. Land-use change, urbanization, invasion of forests, and socioeconomic activities affect vegetation cover, mosquito density, and biting behaviour, increasing contact among wildlife, livestock, and humans [36,37]. At the same time, globalization has facilitated the spread of mosquitoes beyond their historical ranges, accompanied by ecological and behavioural adaptations that enable persistence in diverse urban and peri-urban environments [38].
These processes are well documented for Ae. aegypti and Ae. albopictus, whose urban adaptation has driven the global expansion of Dengue, Zika, and Chikungunya viruses [39,40]. In contrast, many arboviruses of African origin remain poorly characterized despite evidence that Africa’s diverse ecosystems host numerous competent or suspected vectors [8,41–43]. Rapid environmental change may further alter vector distributions and transmission pathways, increasing the potential for emergence within and beyond Africa [40]. Wesselsbron virus (WSLV), Middelburg virus (MIDV), and Sindbis virus (SINV) were first described in Africa between the 1950s and 1970s, yet their contemporary epidemiology and epidemic potential remain poorly understood [2,4,7,11,23,29]. Although detected in sub-Saharan and East Africa, recognized outbreaks have been largely confined to Southern Africa (WSLV and MIDV) and Europe (SINV), and their potential distribution across Africa remains uncertain [9,12,15,29,31,33,44]. Surveillance gaps, exclusion from neglected tropical disease monitoring, and limited differential diagnostics further hinder understanding of these viruses, leading to underdetection or misclassification. For example, sporadic detections of WSLV were made during Rift Valley Fever (RVF) outbreaks [3,5,12]. In addition, human cases may also be misdiagnosed due to similar symptom presentation with other arboviruses and limited capacities for differential diagnosis [31].
While spatial modeling has effectively guided surveillance and prevention for Dengue, Zika, and Yellow fever [45–47], comparable continental-scale assessments are limited for WSLV, SINV, and MIDV.
We modeled the environmental suitability of WSLV, SINV, and MIDV and their mosquito vectors across Africa under current and future climatic and land-use conditions. By focusing on ecological suitability rather than realized transmission, this study provides a macroecological baseline to identify areas of potential virus presence and human exposure, supporting targeted surveillance and future ecological and epidemiological investigations [47,48].
Materials and methods
Ethics statement
This study received ethical clearance from the University of Bonn Center for Development Research and the Charité Universitätsmedizin Berlin. No human subjects were involved in this study. We used openly accessible sources for secondary data, which had no person identifiable data, and we acknowledge the authors/sources.
Study site
Africa is the main source of mosquito-borne viruses of global public health importance [4]. In this study, we combined data from different sources across Africa, including online databases, literature, surveys, and primary data from Uganda, with ecological data covering Africa, to model the habitat suitability of mosquito and arbovirus species under current and future climatic conditions.
Species and presence records
Viruses
This study assesses the habitat suitability for three neglected arboviruses, Wesselsbron, Sindbis, and Middelburg viruses. We selected these viruses due to their epidemic potential and the insufficient attention in Africa given to potential human exposure amidst ecological change. We obtained presence records for WSLV, SINV, and MIDV. During February – March 2024, we searched the National Center for Biotechnology Information (NCBI) Virus database for all published nucleotide sequences using the following taxonomic identifications: Wesselsbron virus (164416), Sindbis virus (11034), and Middelburg virus (11023) [49]. We also searched NCBI’s PubMed, a literature database using the same scientific names, for studies in Africa that identified these viruses. To limit the risk of misclassification, we selected studies where one of the samples underwent virus isolation (for older studies) and polymerase chain reaction (PCR) or genome sequencing (for recent studies). We extracted the date, location, and host of the sample collection. Where location coordinates were missing, we used the sampling site description to identify proximal coordinates in Google Maps [50]. We removed duplicate records and coordinates resulting in: WSLV (n = 154), SINV (n = 109), and MIDV (n = 49) (Fig 1). S2 Table in the Supporting Information shows the sources of the presence points.
Presence points of the mosquito and virus species used in species distribution modeling; S2 Table shows the sources of the presence points.Map was plotted in R on a base layer derived from the openAFRICA “Africa Shapefiles” dataset: https://open.africa/dataset/africa-shapefiles/resource/dcdadd25-0137-4c93-ae5a-82b39d424d60, released under a Creative Commons 4.0 International License.
Mosquitoes
During the literature review conducted in February–March 2024, we collated the main vector species implicated in virus epidemiology, ranking them by evidence and viral detections (S1 Table). However, the final selection of the vectors to use as predictors was constrained to those that fulfilled the former criteria but also had robust presence points in Africa sufficient to generate vector niche models that could be applied in the virus models. In Africa, WSLV has been detected in multiple Aedes species including Ae. caballus, Ae. juppi, Ae. circumluteolus, Ae. mcintoshi, Ae. luridus, Ae. vexans, Ae. unidentatus, Ae. gibbinsi and Ae. tricholabis as well as Ma. uniformis, An. coustani and Culex species. [4,8–10]. Of the 154 WSLV detections we identified, 44 were from mosquitoes, with at least 12 different species. Ae. vexans (7) had the highest detections, followed by Ae. circumluteolus (6) and Ae. mcintoshi (4) (S1 Table). Vector modeling was restricted to Ae. circumluteolus and Ae. mcintoshi, which are consistently identified as the primary enzootic vectors and as the epizootic vectors associated with livestock outbreaks and floodwater-driven transmission across Africa [3,9,13,51].
Multiple Culex species (e.g., Cx. pipiens, Cx. torrentium, Cx. univittatus, Cx. perexiguus, Cx. quinquefasciatus, Cx. modestus), as well as Culiseta morsistans and, less consistently, certain Aedes and Anopheles taxa have been reported infected with or competent for SINV [17,24,25,52]. Although many Culex species are implicated as vectors, SINV isolations are dominated by Cx. univittatus and Cx. pipiens in Africa, while in Europe Cx. torrentium/Cx. pipiens, Cx. perexiguus, Culiseta morsistans, Ae. cinereus and Ae. Ochlerotatus caballus play important roles [17,24,53,54]. Of the 109 SINV presence points we identified, 44 were from at least 17 different mosquito hosts; these were dominated by the Culex genus (31), featuring multiple species, led by Cx. univittatus (15) and Cx. pipiens (4) (S1 Table). Therefore, we selected Cx. univittatus and Cx. pipiens as focal vectors.
Field and molecular studies have detected MIDV in multiple Aedes species, including Ae. caballus, Ae. mcintonshi, Ae. circumluteolus, Ae. dalzieli, and members of the Aedes (Aedimorphus) group, as well as other mosquito taxa [29,30,32,33,55]. Earlier entomological investigations in southern Africa suggested that floodwater Aedes species, including Aedes (Ochlerotatus) juppi, Ae. caballus, and Ae. mcintoshi, may contribute to MIDV maintenance and episodic amplification, potentially through vertical transmission and co-circulation with other arboviruses such as WSLV and SINV [9,28]. Given this evidence and the absence of consistent competence data across species, we adopted a conservative modeling strategy. Among Ae. mosquitoes, we prioritized Ae. mcintoshi due to its ecological association with floodwater-driven transmission systems, repeated detection in recent surveillance studies, and sufficient availability of occurrence data for spatial modeling [32,55] (S1 Table). We complemented this with Ma. africana to represent a non-Aedes mosquito potentially involved in enzootic transmission [4,33].
Presence records for mosquitoes.
We searched the Global Biodiversity Information Facility (GBIF) database using the following GBIF taxon identifications: Ae. circumluteolus (Theobald, 1908) (1651224), Ae. mcintoshi Huang, 1985 (1651403), Cx. univittatus Theobald, 1901 (1653264), Cx. pipiens Linnaeus, 1758 (1652991), Cx. perexiguus Theobald, 1903 (1653140), Ma. africana (Theobald, 1901) (5089851) [56]. Additional presence points were obtained from mosquito sampling in Uganda [33]. The extracted data were cleaned to remove duplicates resulting in Ae. circumluteolus (n = 121), Ae. mcintoshi (n = 89), Cx. univittatus (n = 172), Cx. pipiens (n = 241) and Ma. africana (n = 157) (Fig 1).
Ecological variables
This study integrated climatic, environmental, and land-use data to represent the ecologies of mosquitoes and viruses which contribute to disease transmission [57]. We selected precipitation, temperature, forest cover, normalized difference vegetation index, NDVI (NDVI), agriculture, human population, settlements, and urbanization as key predictors. These variables have been found to influence mosquito survival, breeding, and virus transmission [47,48,58]. Where available, we chose temporal scales of 2015 (current) and 2040 (future) to align species presence records with available environmental data [59].
Climatic variables.
We used WorldClim version 2.1, whose data are derived from over 9,000 weather stations, to obtain 19 bioclimatic variables for 1970–2000; the variables represent key ecological processes [60] (S3 Table). Shared Socioeconomic Pathways (SSPs) are the Intergovernmental Panel on Climate Change’s projections of future emissions and climate. We chose SSP2-4.5 for moderate and SSP5-8.5 for severe conditions [61]. We selected IPSL - CM6A - LR and HadGEM - GC31 - LL Global Climate Models (GCM) from the Coupled Model Intercomparison Project Phase 6 for their strong performance in Africa [62,63]. We downloaded data for 2021–2040 for these models and SSPs from WorldClim [60].
Livestock.
We accessed livestock data from the Food and Agriculture Organisation’s Gridded Livestock of the World (GLW) version 4 for 2015 [64]. Since high-resolution livestock datasets for future scenarios are not available, we used the GLW data version 4 from 2020, a high-resolution compilation of livestock densities [65], to model future suitability and assumed no change in densities.
Human population and urbanization.
We extracted human population and settlement model grids (urbanization) for 2015 and built-up areas for 2014 from the “Global Human Settlement Layer” dataset [66,67]. For future human populations, we accessed projections for SSP2-4.5 and SSP5-8.5 [68].
Cropland.
We extracted the Global Land Analysis and Discovery laboratory’s data for 2012–2015, comprising the percent of croplands per pixel [69]. We derived future land-use from the Global Change Analysis Model and Demeter datasets for SSP2-4.5 and SSP5-8.5 for 2040 at 0.05-degree resolution and extracted layers for crops, forests and urban land - use [70].
Forest.
We downloaded the Moderate Resolution Imaging Spectroradiometer (MODIS) Land Cover Type (MCD12Q1) Version 6.1 dataset of the International Geosphere-Biosphere Programme for 2015 and extracted forested areas [71,72]. To estimate future forested areas in 2040, we utilized the Global Change Analysis Model and Demeter datasets for SSP2-4.5 and SSP5-8.5 [70].
Normalized difference vegetation index.
We extracted the NDVI data for 2015 at 1 kilometer (km) resolution from the Terra MODIS Vegetation Indices (MOD13A3) Version 6.1 monthly dataset [72,73]. For future scenarios, high-resolution NDVI data for Africa was unavailable.
Suitability modeling
The ecological niche of vectors, hosts, or reservoirs indicates geographical areas with potential for virus transmission [74]. We applied the suitability distribution models of vectors as predictor variables in the models of the respective viruses.
Pre-modeling data processing.
The final predictor dataset comprised 26 environmental variables (Table 1); for detailed descriptions, see S3 Table). We processed all variables to cover a uniform spatial extent (Africa), with the same projection (EPSG: 4326-WGS 84), resolution (0.00833 × 0.00833 grid cell size where 1-degree latitude ~111.32 km^2^ per grid), alignment, and ASCII grid format [75]. We used R version 4.4.1 and the packages “sf” and “terra” to process the data [76–78].
Table 1: Ecological variables used in the study.
Species distribution modeling.
Species distribution models (SDMs) use ecological variables and presence points of species to predict the habitat suitability of a species across the landscape. They indicate where conditions are suitable for the species on a continuous scale (0–1). SDMs are widely applied in studying the current and future habitat suitability of vectors and diseases under climatic changes [46,47,58,79]. Maximum entropy (Maxent), a popular SDM method, predicts habitat suitability based on species presence and environmental conditions and projects to future environmental scenarios. The resulting species distribution maps indicate where conditions are suitable for the species on a continuous scale (0–1) [59,80]. Presence maps show where the species are estimated to occur based on thresholds applied to the suitability models. They convert suitability into stacked levels of certainty of species presence, indicating where the species are likely present with higher confidence. Species distribution models that use presence-only species data are prone to spatial bias that can be introduced by differing methods and intensity of species sampling across studies [59]. We utilized the target-group background approach to minimize sampling bias by using collective occurrence records of the genera to which our study species belong to reflect the general sampling effort [81,82]. Therefore, occurrences in Africa of the genera Aedes Meigen, 1818*(for species Ae. circumluteolus and Ae. mcintoshi), Culex Linnaeus,* 1758 (for species Cx. univittatus and Cx. pipiens), and Mansonia Blanchard*, 1901 (*for species Ma. africana) were extracted from GBIF (S2 Table). We converted the genera’s sampling points into gridded layers using kernel density estimations under the R package “MASS” and incorporated these in the respective Maxent models of the vectors [81,82].
In the first step, we ran Maxent version 3.4.4 [59,80], using all ecological variables, a genus-specific bias grid, automatic feature classes, and split the occurrence data into 75% training and 25% testing. We used the jackknife test, set the regularization multiplier to 3, conducted 5 cross-validation runs, set a convergence threshold of 1.0e-5, used a maximum of 10,000 background points and 500 maximum iterations. In a second step, we considered the averages of the cross-validation runs to drop variables of low importance and high Pearson correlation (over 0.7) [75]. See the correlation matrix in S1 Fig. This enabled the exclusion of less important predictors and addressed multicollinearity in subsequent species modeling.
Parameter setting optimization.
Selection of an appropriate combination of settings in Maxent is challenging. In the third step, we used the “kuenm” approach to optimize the calibration of multiple candidate models, selection, evaluation, and projection [83]. We organized occurrence data into four sets: random independent points (10), joint (minus independent points), training (70% of joint), and testing (30% of joint). Per species, we combined the variables chosen in step 2 with one of two regularization multipliers (3 or 5) and a basic combination of feature classes to produce sets of variables and candidate models. During each calibration, we created a candidate model using each joint and training set of occurrences.
Candidate model evaluation.
We measured statistical significance using the partial receiver operating characteristic (pROC), 50% of data for bootstrapping, and 500 iterations [84]. We assessed predictive ability by the omission rate (OR) and allowed a 5% omission error (E) [85]. We evaluated model complexity using the Akaike Information Criterion corrected (AICc) for small sample sizes [83]. We selected best-performing candidate models based on statistical significance, OR below 5%, and delta AICc less than 2. S4 Table shows the model settings and results.
Final models.
We generated final models using the best - performing settings with five bootstrap replicates, projecting them with extrapolation and clamping based on future GCM and SSP variables. Each model was evaluated using independent points (S4 Table). Output models presented as maps show predicted habitat suitability probabilities ranging from 0 (least suitable) to 1 (most suitable). We calculated suitability differences between current and future models. For mosquitoes, we reclassified suitability into low (0 - 0.3), average (0.4 - 0.6), and high (0.7 - 1) categories, estimating the area (km^2^) per class. We converted suitability models into species presence models by assigning presence to cells with suitability above four thresholds: 1) Equal training sensitivity and specificity; 2) Maximum training sensitivity plus specificity; 3) Balanced training omission; and 4) Ten-percentile training presence. This created a stack of four presence/absence models per species and scenario, showing the number of threshold criteria (0–4) under which a given cell is predicted as suitable. For viruses, we combined these stacks to produce hotspot maps highlighting areas with the highest certainty of species presence (value 12) and least certainty (value 0). We overlaid hotspot maps with human population data for 2015 and 2040 to visualize population as a predictor of virus presence and likely exposure. We used R Studio and the packages “ggplot2”, “terra”, “viridis”, and “gridExtra” to visualize figures [76,78,86–88]. Administrative boundary data were obtained from the openAFRICA “Africa Shapefiles” dataset released under a Creative Commons 4.0 International License.
Results
We modeled habitat suitability for three viruses (WSLV, SINV, and MIDV) and five mosquito vectors (Ae. circumluteolus, Ae. mcintoshi, Cx. univittatus, Cx. pipiens and Ma. africana). We analyzed at least 100 candidate models for each mosquito or virus species, most of which were significantly better in predicting species presence than a randomized prediction (S4 Table). The best candidate models had above-random habitat suitability predictions with high mean Area Under the Curve (AUC) ratios (Range = 1.45 - 1.88). Final models produced using the best candidate model settings resulted in higher mean AUC ratios (Range = 1.67 - 1.97) implying improved fitness of training data and better predictive performance (S4 Table). To facilitate interpretation, habitat suitability outputs (continuous probabilities from 0 to 1) were converted into threshold-based presence maps summarizing agreement across four presence–absence criteria. The resulting presence index (0–4) reflects increasing certainty of environmentally suitable conditions for virus presence.
Habitat suitability for vectors
Under current ecological conditions, we predicted varied suitability for the five mosquitoes, particularly across Equatorial and Southern African countries. Suitability was predicted to remain relatively stable under both scenarios during 2021–2040 with small geographical expansions for Cx. univittatus, Ma. africana, Ae. mcintoshi, and Cx. pipiens. Areas with high precipitation in warm (1,300mm) and cold quarters (3,000mm) were suitable for Ae. cicumluteolus but not Ae. mcintoshi. Both Aedes species required at least 23°C for average to high suitability and preferred temperatures less than 34°C, while Culex species’ suitability was sensitive to extremely low (below 7°C) and high temperatures (above 32°C). For Aedes and Culex species, we found average suitability values in areas that received at least 400 mm of precipitation in dry quarters. Suitability was positively influenced by built-up areas (for Culex species) and livestock (for all mosquitoes, except Ma. africana).
We predicted areas with highly suitable habitats for Ae. circumluteolus primarily in equatorial countries from the West African coast to Central and Eastern Africa, including the North-Eastern horn (Fig 2). Currently, areas with low suitable habitats cover 2,360,000 km^2,^ increasing under SSP2-4.5 (42%) and SSP5-8.5 (43%). In contrast, highly suitable habitats were limited to 176,000 km^2^, declining under SSP2-4.5 (-49%) and SSP5-8.5 (-53%) (Fig 5 and S5 Table). Current highly suitable habitats were concentrated in Central and Eastern Africa and Madagascar. Future projections showed the highest certainty of presence was limited to Central and Eastern Africa (Fig 2). The key predictive variables were isothermality (47%) and precipitation of the driest month (35%). The least important were built-up areas (0.12%) and forests (0.03%). Suitability increased with isothermality, minimum temperature (from 0.2 at -30°C to 0.5 at 23°C), precipitation of the driest month (from 0.6 at 8mm to 0.8 at 140mm), warmest quarter precipitation (0.5 at 200mm to 0.9 at 1,300mm), coldest quarter precipitation (steady at 0.5 from 1mm to 3,500mm), and livestock (from 0.7 at 0 animals to 0.95 at 550,000 animals per 111km^2^).
Habitat suitability and presence for Ae. circumlutelous and Ae. mcintoshi currently and under future (2021 – 2040, SSP2-4.5 and SSP5-8.5) ecology.For suitability scenarios, positive values indicate an increase, and negative values, a decrease in suitability. Presence maps display a threshold-based certainty index (from least certainty (0) to highest certainty (4)) representing the number of presence–absence models in which suitability exceeded predefined thresholds. Africa base layer derived from the openAFRICA “Africa Shapefiles” dataset: https://open.africa/dataset/africa-shapefiles/resource/dcdadd25-0137-4c93-ae5a-82b39d424d60, released under a Creative Commons 4.0 International License.
We found areas with high suitability for Ae. mcintoshi mainly in East Africa, Malawi, and Southern Africa, with increased suitability across Sub-Saharan Africa (SSA), particularly in the Democratic Republic of Congo (DRC), Ethiopia, and Somalia under SSP2–45 and SSP5–85 (Fig 2). Areas with low suitability covered the most area (2,346,000 km^2^) and were expected to increase by 32%, while highly suitable habitats (101,000 km^2^) were projected to decline under SSP2-4.5 (-31%) and SSP5-8.5 (-34%) (Fig 5, and S5 Table). Future presence was predicted to spread to Central Africa, Ethiopia, and Somalia (Fig 2). Precipitation of the warmest quarter (34%) and human population (33%) were the best predictors for suitability, while livestock (1.5%) and built-up areas (0.5%), the least. Suitability responded positively to the minimum temperature of the coldest month (from 0.4 at 20 °C - 0.95 at 23 °C), precipitation of the warmest quarter (peaked at 0.6 at 100mm), built-up areas, and livestock. It responded negatively to the maximum temperature of the warmest month (plateaued at 0.55 until 34 °C, then declined), precipitation of the coldest quarter (peaks at 0.5 at 300mm, then declined to 0 at 3,000mm), forest cover, and human population.
The Cx. pipiens model predicted low suitability across most areas (21,720,000 km^2^) with average suitability spanning 9,100,000 km^2^. High-suitability zones covered approximately 250,000 km^2^ in select regions of West Africa, the Nile Delta, eastern Africa, Angola, South Africa, and Madagascar (Figs 3 and 5). Under SSP2-4.5 and SSP5-8.5, low suitability areas increased by 39% and 46%, while highly suitable Areas declined by 75% (S5 Table). We observed the highest presence in East Africa, including Malawi, Zambia, Angola, Namibia, South Africa, and Madagascar. Built-up areas (54%) were the primary contributors to habitat suitability, followed by livestock (13%), cropland (2.6%), while precipitation in the driest quarter (1.1%) was the least significant. Suitability increased with the precipitation of the driest quarter (rising from 0.25 at 0 mm to 0.64 at 420 mm), livestock (plateauing at 0.95 with 600,000 animals per 111 km^2^), built-up areas, and human population. Conversely, suitability declined with the maximum temperature of the warmest month (starting at 0.9 from 0 °C to 15 °C, then dropping to 0.3 at 45 °C), the annual temperature range (declining from 0.6 at 8 °C to 0.1 at 46 °C), and cropland. Current habitat suitability for Cx. univittatus was rather low in most areas (29,370,000 km^2^) (S5 Table). High-suitability areas (310,000 km^2^) lay along the Sahara coastline, Nile Delta, South-Western Uganda, Rwanda, South Africa and Reunion Island (Fig 3). Under SSP2-4.5, areas with low suitability increased by 25% and average suitability by 13%. Under SSP5-8.5, highly suitable areas increased by 106%, particularly in the Sahara and Central/Southern Africa (Figs 3 and 5). Current high certainty of presence includes Western Africa, the Nile Delta, and parts of East and Southern Africa. Future projections under SSP2-4.5 show spread into Morocco, Libya, Egypt, and parts of Central Africa. SSP5-8.5 further expands this to Libya and Egypt (Fig 3). The best predictors for suitability are built-up areas (58%) and temperature annual range (20%), while croplands (2.5%) and forests (1%) are the lowest. Suitability increased with building footprint, and livestock (peaking at 0.9 at 1 million animals per 111 km^2^) but declined with forest and cropland covers. Negative responses were noted for minimum temperature of the coldest month (peaked at 0.6 at 7^o^C then declined), temperature annual range (from 0.3 at 14^o^C - 0.6 at 32^o^C, then declined), annual precipitation (from 0.9 at 0 mm - 0.4 at 1,500mm, then declined), and precipitation of driest quarter (peaked at 0.5 at 450mm).
Habitat suitability and presence for Cx. pipiens and Cx. univittatus currently and under future (2021 – 2040, SSP2-4.5 and SSP5-8.5) ecology.For suitability scenarios, positive values indicate an increase, and negative values, a decrease in suitability. Presence maps display a threshold-based certainty index (from least certainty (0) to highest certainty (4)) representing the number of presence–absence models in which suitability exceeded predefined thresholds. Africa base layer derived from the openAFRICA “Africa Shapefiles” dataset: https://open.africa/dataset/africa-shapefiles/resource/dcdadd25-0137-4c93-ae5a-82b39d424d60, released under a Creative Commons 4.0 International License.
Most of Africa (27.33 million km^2^) had low suitability for Ma. africana with highly suitable areas around Uganda, South Sudan, Ethiopia, and Kenya (400,000 km^2^). Average suitability spanned 3.34 million km^2^ in Equatorial Africa. Under both scenarios, suitability increased around Sierra Leone, Liberia, and Central Africa. Under SSP2-4.5, low and average suitability areas increased by 13%, while high suitability decreased by 3%. Under SSP5-8.5, low suitability increased by 22%, average by 16%, and high by 21% (Figs 4, 5 and S5 Table). High certainty of presence was stable across equatorial countries in both future scenarios (Fig 4). Key predictors were isothermality (46%) and human population (24%), while mean temperature of the warmest quarter (0.034%) and precipitation seasonality (4.08%) had minimal influence. Suitability responded positively to isothermality, mean temperature of the warmest quarter (peaked at 0.6 at 17^o^C, declined to 0.15 at 37^o^C), mean temperature of the coldest quarter (peaked at 0.75 at 29^o^C), precipitation of the warmest quarter (peaked at 0.65 at 300mm then declined to 0 at 700mm), precipitation of driest quarter (peaked at 0.75 at 150mm but declined to 0 at 400mm). Suitability decreased with precipitation seasonality, human population (beyond 15,000 people per 111 km^2^), and livestock (beyond 200,000 animals per 111 km^2^).
Habitat suitability and presence for Ma. africana and WSLV currently and under future (2021 – 2040, SSP2-4.5 and SSP5-8.5) ecology.For suitability scenarios, positive values indicate an increase, and negative values, a decrease in suitability. Presence maps display a threshold-based certainty index (from least certainty (0) to highest certainty (4)) representing the number of presence–absence models in which suitability exceeded predefined thresholds. Africa base layer derived from the openAFRICA “Africa Shapefiles” dataset: https://open.africa/dataset/africa-shapefiles/resource/dcdadd25-0137-4c93-ae5a-82b39d424d60, released under a Creative Commons 4.0 International License.
Predicted area for habitat suitability in 10,000 km2 per mosquito species and climate change scenario.Habitat suitability is classified into, low (0 - 0.3), average (0.4 - 0.6) and high (0.7 - 1.0). On the right y-axis, the percent change in the area of habitat suitability for future scenarios is denoted by points.
Predicted ecological suitability for virus presence
Current ecological suitability for WSLV was highest in South Africa, Lesotho, and East Africa. Future habitat suitability was predicted to increase in Lesotho and the equatorial countries from the west to the east of Africa. Current ecological suitability for WSLV presence was highly certain in areas of southern and eastern Africa, increasing in Ivory Coast, Ghana, Nigeria, and the Horn of Africa while declining in South Africa under future scenarios (Fig 4). Key predictors were built-up areas (33%), and habitat suitability of Ae. mcintoshi (25%) and Ae. circumluteolus (11%), with minimal contributors from livestock (0.46%) and cropland (0.81%).
Under the current ecological conditions, limited areas had above-average suitability for SINV, primarily in Nigeria, central Ethiopia, eastern Africa, in South Africa and central Madagascar. Under the future scenario SSP2-4.5, predicted ecologically suitable areas slightly increased across Africa, particularly in coastal West Africa and in countries below the equator; the increases were more obvious under SSP5-8.5 for South Africa and Lesotho. We predicted high ecological suitability with low to medium certainty (suitability above 1–3 threshold values) for SINV presence in the Nile Delta, western coast, east, and southern Africa. Under SSP2-4.5 and SSP5-8.5, the presence shrunk in those areas but slightly increased in coastal Nigeria, DRC, and Angola (Fig 6). Cx. univittattus (73%) contributed most to SINV suitability, followed by Cx. pipiens (9.6%), with minor influence from livestock (1.2%) and human population (0.6%).
Habitat suitability and presence for SINV and MIDV currently and under future (2021 – 2040) SSP2-4.5 and SSP5-8.5 ecology.For suitability scenarios, positive values indicate an increase, and negative values, a decrease in suitability. Presence maps display a threshold-based certainty index (from least certainty (0) to highest certainty (4)) representing the number of presence–absence models in which suitability exceeded predefined thresholds. Africa base layer derived from the openAFRICA “Africa Shapefiles” dataset: https://open.africa/dataset/africa-shapefiles/resource/dcdadd25-0137-4c93-ae5a-82b39d424d60, released under a Creative Commons 4.0 International License.
We predicted highly suitable habitats for MIDV primarily in South Africa and Lesotho. Under both future scenarios, suitability was stable, albeit with slight declines, notably in South Africa. Current ecological suitability for MIDV presence was highly certain in South Africa, Lesotho, Swaziland, and parts of Eastern and Central Africa. Under future scenarios, high certainty expanded slightly to areas in Egypt, Libya, and Algeria (Fig 6). Key predictors were the mean temperature of the driest quarter (31%) and built-up areas (14%), while Ma. africana suitability (1.9%), cropland (1.03%), and human population (0.08%) had minimal impact. Notably, the best model selected for final modeling did not consider Ae. mcintoshi among the most important variables.
Predicting hotspots for risk of zoonotic disease exposure
We analysed WSLV, SINV, and MIDV presence-absence models to map potential exposure risk. High-certainty hotspots (presence scores 9–0) were concentrated in South Africa, Lesotho, Uganda, Kenya, and Ethiopia, while average risk (presence scores 5–8) occurred along the North African coast and parts of SSA. Under SSP2-4.5 and SSP5-8.5, average risk expanded slightly, with low risk potentially emerging in the Sahara (Fig 7).
Hotspots for virus exposure risk currently, and during future (2021 – 2040) SSP2-4.5 and SSP5-8.5 scenarios.Risk is based on the highest (12) and the least (0) certainty of virus presence. Africa base layer derived from the openAFRICA “Africa Shapefiles” dataset: https://open.africa/dataset/africa-shapefiles/resource/dcdadd25-0137-4c93-ae5a-82b39d424d60, released under a Creative Commons 4.0 International License.
We overlaid the combined presence-absence model outputs of the three viruses with gridded human population density across Africa for each scenario. Under current conditions, environmentally suitable areas for low certainty of virus presence (certainty scores 0–4) were more frequently associated with sparsely populated rural areas (<300 people per km^2^), shown in darker red tones, compared with suburban and urban areas, which are predominantly shown in blue (Fig 8). Under future climate scenarios (SSP2-4.5 and SSP5-8.5), environmentally suitable areas for low to moderate certainty of virus presence were projected to expand into increasingly populated suburban and urban settings, illustrated by yellow and green tones (Fig 8).
Current, and future (2021 – 2040, SSP2-4.5 and SSP5-8.5) human population density and ecological suitability for potential arboviral exposure.Human population density is presented as people per square kilometre. Ecological suitability represents the combined predicted presence of WSLV, SINV and MIDV. Warmer colors indicate greater overlap between ecological suitability for virus presence and human populations.
Discussion
The recent devastating outbreaks of Zika and Dengue underscore the need to understand mosquito-borne virus distribution under climate change [40,46,89]. This study predicts areas in Africa where current and future ecology may support five vectors (Ae. circumluteolus, Ae. mcintoshi, Cx. pipiens, Cx. univittatus, and Ma. africana) and the potential exposure of WSLV, SINV, and MIDV to humans. We present predicted species’ habitat suitability and presence, summarizing the consistency of habitat suitability exceeding multiple threshold criteria.
Our models predict that most SSA countries show variable suitability for the analyzed mosquito species, a finding consistent with mosquito collections since the 1970s which identified arbovirus vectors in multiple countries [42,51]. High suitability is predicted in southern Africa, particularly Lesotho and Northern South Africa, where recent surveys report abundant Culex, Aedes, and Mansonia in conservation areas [42,55]. Unlike dry semi-deserts, conservation areas have more ecological integrity, standing water, and wildlife to support mosquito populations [42]. In arid regions of Kenya, wetlands and seasonal flooding enable breeding for Ae. mcintoshi and Ma. africana [90,91]. Ma. africana, Ae. mcintoshi and Ae. circumluteolus are found in savannas and grasslands, while Cx. pipiens occurs near lake shores, but also in urban areas, villages and human settlements [51,55,91]. The adaptability of arbovirus vectors across ecosystems highlights the need for diverse entomological surveillance.
Aedes species were more tolerant of the coldest and driest seasons compared to Ma. africana and Culex species, but all mosquitoes preferred maximum temperatures below 32–45°C. In South Africa, Aedes species are reported to thrive better between 18–27°C. High temperatures and rainfall typically increase mosquito breeding, serving as an early warning for risk of infection exposure [55,90]. Seasonal mosquito control is essential even during short rains of dry seasons [91]. In addition to climate, built-up areas, human population, and livestock density were key predictors of mosquito habitats. Ma. Africana, Ae. mcintoshi, and Ae. circumluteolus prefer savanna and grasslands but also thrive in urban areas [42,51,55]. Mosquitoes are adapting to breeding in containers and feeding on livestock and humans near buildings [39]. Urbanization also increases transference risks via mobility and trade [4].
Our virus models did not include niche models for all known mosquito-virus detections; instead, focal vector species were chosen. We acknowledge that omitting other species could bias spatial predictions where those other suspected vectors dominate local transmission cycles. However, detecting a virus in a mosquito does not necessarily establish it as a competent vector in viral maintenance. Notably, for SINV, while enzootic transmission in Europe is sustained mainly by Cx. torrentium, in Africa, Cx. univittatus is the principal vector with Cx. pipiens, Cx. neavei, Cx. quinquefasciatus and Cx. perexiguus playing a supportive role [22,41,54,92,93]. Although WSLV has a broad vector range, enzootic and epizootic cycles rely on floodwater Aedes mosquitoes (notably Ae. circumluteolus and Ae. mcintoshi), which are also the primary vectors in livestock outbreaks after heavy rains [3,9,13,51]. Surveillance studies associate MIDV with several mosquito genera, suggestive of a heterogeneous assemblage of mosquito vectors rather than a single dominant species. However, much of this evidence is based on field isolations rather than controlled vector competence studies [4,32,33,55]. Future studies should explore the relevance of specific floodwater Aedes species and Mansonia species in MIDV flood-driven transmission, mammal host amplifying and enzootic circulation roles [4,9,28,33]. We also recommend robust mosquito surveys across Africa to investigate the role of different species, local abundance, infection rates and competence, as has been done for SINV in South Africa and Europe [22,54,92,93].
We predicted low ecological suitability for WSLV, SNV, and MIDV presence in Africa, with hotspots in Southern, Eastern, and Western coastal regions. Future scenarios predicted small expansions in North Africa, the Horn of Africa, and the west coast. These findings align with reported sporadic cases and highlight ecological risks for potential human exposure to aid prevention and early warning [3,5,15,30].
We applied the individual niches of the mosquito vectors and included the ecology of known livestock hosts as predictors in our models. The best predictors of the viruses were the ecological niches of their mosquito vectors, a finding corroborated by models of Dengue using its vector, Ae. aegypti [57]. However, arboviral disease epidemiology is often complicated by the multitude of vectors, hosts, ecological and human susceptibility factors at work which can’t be fully modelled. Furthermore, unlike Dengue with established primary vectors - Aedes albopictus and/or Aedes aegypti and humans as the main hosts, the arboviruses in our study are a part of multi-vector multi-host transmission networks that are still under investigation [94]. For example, in Sweden, the seasonal abundance of and SINV prevalence in the main enzootic vector Cx. torrentium and in the bridge vector Ae. cinereus, are important predictors of outbreaks [24]. Similarly, in South Africa, SINV detections in the enzootic vector Cx. univittatus has been linked to increased human infections. However, Cx. univittatus and other vectors’ presence alone doesn’t necessarily indicate SINV infection, but incidence is rather seasonal, coinciding with the warm, wet season [21,54]. Furthermore, our study points to the role of climatic predictors in vector and virus ecology, corroborating findings in South Africa linking SINV and Rift Valley Fever (RVF) surges to above-average rainfall and vector breeding [21]. In settings where the evidence for vector-host-virus ecology is underdeveloped, our ecological risk maps can indicate areas that could be targeted for mosquito sampling and virus screening as part of early-warning.
The vectors we analyzed are also important vectors for other viruses of public health importance, i.e., Ae. circumluteolus (Spondweni, Zika, Lebombo, Kedougou, Bunyamwera, Ndumu, Pongola and RVF); Ae. mcintoshi (Babanki, Ndumu, Bunyamwera, Ngari, Pongola, RVF); Cx. univittatus (West Nile, Usutu, WSLV, RVF); Cx. pipiens (West Nile, Usutu, Semliki Forest, RVF, Japanese Encephalitis, Venezuelan Equine Encephalitis), Ma. africana (Spondweni, Usutu, RVF) [4,95,96]. The diversity of niches suitable for these vectors implies that where suitable blood meals or potential hosts (bird, livestock, or human) are present, the transmission, maintenance, and spread of multiple arboviral diseases is likely [4]. Since vector-virus interactions for emerging diseases are often not well understood in the African context, broad mosquito control measures are recommended over species-specific approaches [51,97]. However, mosquito surveillance to detect virus circulation has the best potential for assessing potential risk for human exposure to strengthen early warning and justify resource allocation for intensive control interventions [24,98].
Incorporation of periodic surveys and arbovirus screening into public health systems could improve countries’ knowledge of circulating neglected arboviruses and improve early detection [7,99,100]. In addition, countries should operationalize a One Health approach with multi-sectoral coordinated interventions covering human health, vector control, human activities, animal (wild and domestic) health, as well as ecological monitoring [101]. Countries can leverage existing national and regional policy frameworks such as the Integrated Disease Surveillance and Response (IDSR), and One Health initiatives by the Africa Centers for Disease Control to expand surveillance beyond detection to pre-epidemic multi-host and multi-sectoral surveillance cognizant of ecological risks of arboviral emergence and human exposure risk [102,103].
Our projections suggest a future shift in the spatial overlap between ecological suitability for virus presence and human populations, from predominantly rural settings toward more densely populated urban areas under future socioeconomic and climate scenarios. While this does not imply increased transmission risk per se, it positions human population as another important attribute of ecological transitions that could modify virus presence and human exposure. In Sub-Saharan Africa, rapid population growth and land-use change—characterised by the conversion of forested or wetlands into agricultural, periurban, and built-up environments—may similarly reshape vector habitats and host availability. Such transitions have the potential to modify primarily sylvatic transmission cycles by influencing mosquito behavior, host-seeking patterns, and contact rates between anthropophilic vectors and humans, thereby altering the ecological landscape in which arboviruses persist. [4,35].
Livestock density was an important predictor of vector presence, signalling them as potential amplifying blood meals. In East Africa, Ae. mcintoshi and Ma. africana abundance were detected in livestock-dense communities where they influenced arboviral activity [41,90]. Syndromic and sentinel surveillance, especially in rural and peri-urban hotspots near livestock, could enhance early detection and understanding of arbovirus transmission and seasonality of risk. Animals like the African Thrushes (for SINV), horses (for SINV and MIDV), and sheep and goats (for WSLV and MIDV) implicated in arboviral maintenance can also contribute to sentinel surveillance [26,27]. For example, SINV sentinel studies in South Africa showed higher pigeon infection risks during warm, wet seasons [54]. Another South African study demonstrated that infected wild and non-equine animals present with neurologic and even fatal SINV infection that could constitute early warnings of cross-species spill overs [27].
Although we evaluated model performance using pROC, AICc, and bootstrap procedures, we acknowledge that bootstrapping with relatively small and spatially clustered species occurrence datasets could have underestimated model uncertainty. Additionally, to address model overfitting due to the limited viral occurrence records, several safeguards were implemented to enhance model robustness. Sampling bias was reduced using a target-group background approach derived from extensive Aedes, Culex, and Mansonia genus occurrence data, a recommended strategy for presence-only species distribution modeling [59,104]. Model complexity was further constrained through conservative regularization, restricted feature classes, variable selection, and AICc-based model evaluation, which explicitly penalizes over-parameterization in small-sample contexts [83]. Predictive performance was assessed using independent testing data, partial ROC, and omission rate thresholds, providing complementary measures of model reliability. Nevertheless, uncertainty remains in regions with sparse data, and model outputs should therefore be interpreted as indicators of ecological suitability for virus presence across ecological scenarios rather than precise outbreak-prone areas.
Many viral pathogens in this study are not notifiable or routinely monitored, limiting case data from many African countries. While serological tests offer basic diagnostic capacity, advanced methods like PCR, virus isolation, and sequencing are more reliable but underutilized due to limited resources, as reflected in the sparse virus sequences in public databases. Despite these challenges, we are confident that we compiled a representative dataset from surveillance, outbreaks, and research, incorporating diverse diagnostic methods and reputable sources. Additionally, we did not have access to a suitable dataset to represent the hosts that influence arboviral transmission, and we only included livestock as a predictor in our models. Where data are available, comprehensive models that account for stochastic introduction events, host susceptibility, host biting, and transmission rates could improve risk mapping [105,106].
Arboviral transmission depends on complex and dynamic processes—including vector–host interactions, vector competence, pathogen prevalence, immunity, and human mobility—that cannot be fully captured using presence-only ecological models. In this study, ecological suitability maps are intended to represent ecological conditions associated with documented virus occurrence rather than realized transmission or infection risk. Similar approaches have been used to map potential ecological suitability or exposure to viruses with complex or poorly resolved transmission systems, including Zika, Ebola Virus, and Leishmaniases [47,107,108]. Given the incomplete knowledge of vector competence and host amplification for many arboviruses, these outputs should be interpreted as screening tools to guide surveillance and ecological inquiry, not as definitive predictors of disease transmission.
Conclusion
This study presents a continental-scale assessment of the ecological suitability of selected African arboviruses and their mosquito vectors under current and future climatic and environmental conditions. Using presence-only data and macroecological modeling, we identify areas where ecological conditions may support virus presence and where potential human exposure could occur, while clearly distinguishing these patterns from realized transmission risk. The results reveal pronounced spatial heterogeneity and projected shifts in suitability, highlighting the influence of ecological change on arbovirus landscapes.
These outputs should be interpreted as screening tools rather than predictors of disease occurrence. Given the complex, multi-host and multi-vector transmission systems of most zoonotic arboviruses, the maps provide a baseline to guide surveillance, entomological studies, and ecological investigations in data-limited regions. Future integration of vector competence, host dynamics, and epidemiological data will be essential to translate ecological suitability into refined assessments of transmission risk.
Supporting information
S1 TableMosquito host per virus species identified in literature.(DOCX)
S2 TableSources of presence points of the mosquito and virus species.(DOCX)
S3 TableSource datasets for the ecological variables.(DOCX)
S4 TableModel settings and results per species.(DOCX)
S5 TableArea of mosquito species habitat suitability and percent change per scenario.(DOCX)
S1 FigCorrelation matrix of ecological variables.(PNG)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature. 2008;451(7181):990–3. doi: 10.1038/nature 06536 18288193 PMC 5960580 · doi ↗ · pubmed ↗
- 2Weiss KEH, Alexander RA, Clark R, Louw JG, De Kock VE. Wesselsbron virus - a virus not previously described, associated with abortion in domestic animals. Onderstepoort Journal of Veterinary Research. 1956;27.
- 3Weyer J, Thomas J, Leman PA, Grobbelaar AA, Kemp A, Paweska JT. Human cases of Wesselsbron disease, South Africa 2010-2011. Vector Borne Zoonotic Dis. 2013;13(5):330–6. doi: 10.1089/vbz.2012.1181 23473219 · doi ↗ · pubmed ↗
- 4Braack L, Gouveia de Almeida AP, Cornel AJ, Swanepoel R, de Jager C. Mosquito-borne arboviruses of African origin: review of key viruses and vectors. Parasit Vectors. 2018;11(1):29. doi: 10.1186/s 13071-017-2559-9 29316963 PMC 5759361 · doi ↗ · pubmed ↗
- 5Diagne MM, Faye M, Faye O, Sow A, Balique F, Sembène M, et al. Emergence of Wesselsbron virus among black rat and humans in Eastern Senegal in 2013. One Health. 2017;3:23–8. doi: 10.1016/j.onehlt.2017.02.001 28616499 PMC 5454166 · doi ↗ · pubmed ↗
- 6Henderson BE, Kirya GB, Hewitt LE. Serological survey for arboviruses in Uganda, 1967-69. Bulletin of the World Health Organization. 1970;42(5):797–805. 5311064 PMC 2427494 · pubmed ↗
- 7Kokernot RH, Szlamp EL, Levitt J, Mc Intosh BM. Survey for antibodies against arthropod-borne viruses in the sera of indigenous residents of the Caprivi Strip and Bechuanaland Protectorate. Trans R Soc Trop Med Hyg. 1965;59(5):553–62. doi: 10.1016/0035-9203(65)90158-6 5893148 · doi ↗ · pubmed ↗
- 8Diallo M, Nabeth P, Ba K, Sall AA, Ba Y, Mondo M, et al. Mosquito vectors of the 1998-1999 outbreak of Rift Valley Fever and other arboviruses (Bagaza, Sanar, Wesselsbron and West Nile) in Mauritania and Senegal. Med Vet Entomol. 2005;19(2):119–26. doi: 10.1111/j.0269-283X.2005.00564.x 15958020 · doi ↗ · pubmed ↗