Diagnostic challenges of histidine-rich protein 2-based rapid diagnostic tests due to pfhrp2 and pfhrp3 gene deletions in asymptomatic malaria in Tanzania
Ernest Mazigo, Hojong Jun, Wang-Jong Lee, Johnsy Mary Louis, Fadhila Fitriana, Jadidan Hada Syahada, Fauzi Muh, Wanjoo Chun, Won Sun Park, Se Jin Lee, Sunghun Na, Eun-Taek Han, Feng Lu, Winifrida Kidima, Jin-Hee Han

TL;DR
This study in Tanzania shows that HRP2-based malaria tests are mostly effective for asymptomatic cases, but gene deletions in some parasites can cause false negatives, highlighting the need for ongoing monitoring.
Contribution
The study evaluates the impact of pfhrp2/3 gene deletions on HRP2-based RDT performance in asymptomatic malaria in Tanzania.
Findings
HRP2-based RDTs detected 77.6% of qPCR-positive cases, with higher sensitivity than light microscopy.
Gene deletions were found in 11.8% of samples, with higher prevalence in high-transmission areas.
RDTs showed substantial agreement with qPCR, but dual-deletion isolates were all RDT-negative.
Abstract
Histidine-rich protein 2 (HRP2)-based rapid diagnostic tests (RDTs) are critical for malaria diagnosis in Africa, particularly in resource-limited settings. However, the spread of Plasmodium falciparum parasites with pfhrp2 and pfhrp3 gene deletions challenges their effectiveness, raising concerns in affected areas. Therefore, this study aimed to assess the prevalence of pfhrp2 and pfhrp3 gene deletions and evaluate the diagnostic performance of HRP2-based RDTs in detecting asymptomatic malaria infections in Tanzania. This cross-sectional community survey study aimed to determine the prevalence of pfhrp2/3 gene deletions from asymptomatic malaria population in Tanzania. Moreover, the study intended to assess the performance of HRP2-based RDT and light microscopy (LM) in detecting asymptomatic malaria infections. The survey was conducted from December 2022 to July 2023 in twelve…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
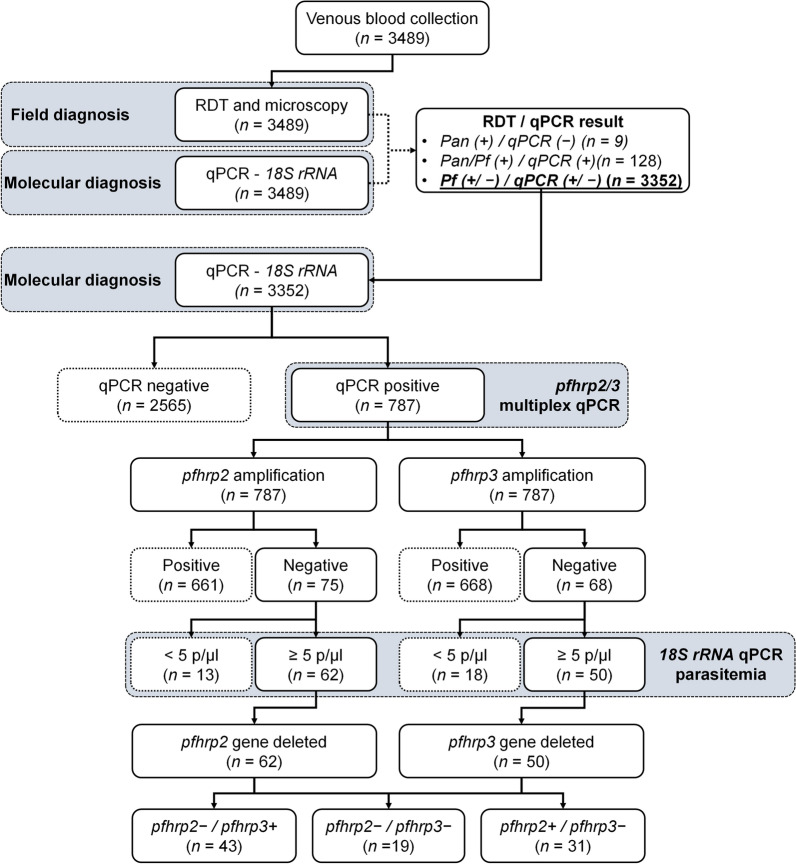 Figure 1
Figure 1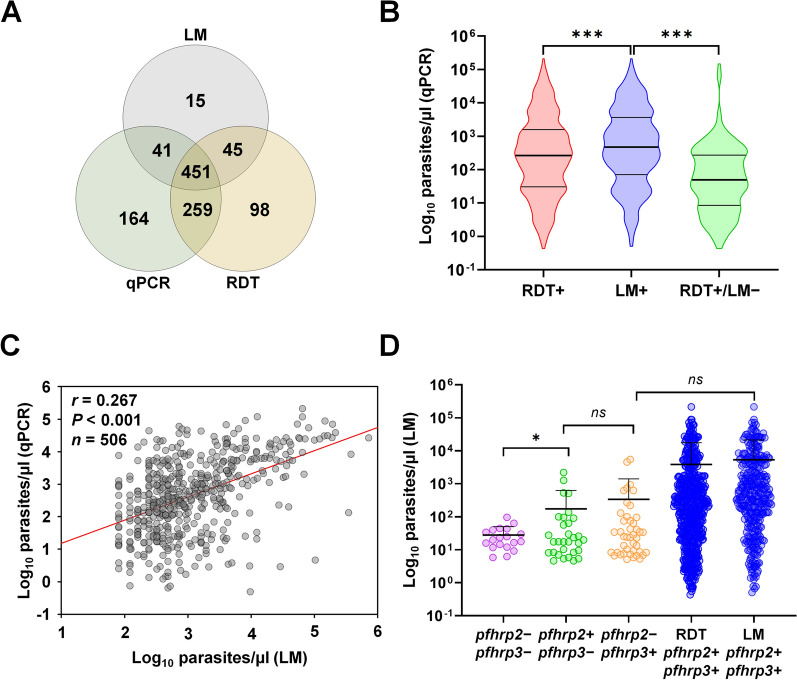 Figure 2
Figure 2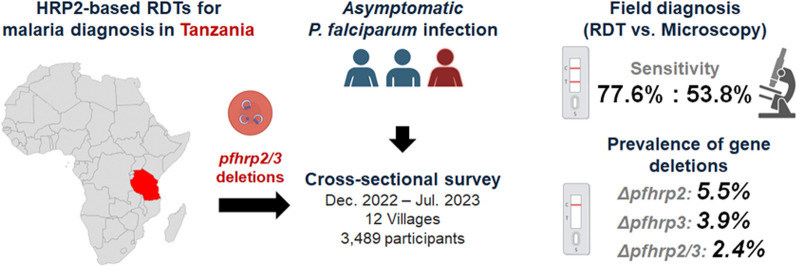 Figure 3
Figure 3- —http://dx.doi.org/10.13039/501100003710Korea Health Industry Development Institute
- —National Research Foundation of Korea
- —http://dx.doi.org/10.13039/501100003725National Research Foundation of Korea
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Genetics, Bioinformatics, and Biomedical Research · Mosquito-borne diseases and control
Background
Malaria is still a public health problem globally, particularly in sub-Saharan Africa (SSA) countries, which account for 94% of the global malaria cases and 95% of deaths, respectively [1]. Evidence shows a resurgence of malaria in areas where the disease has significantly declined [2, 3]. This observation calls for assessment of the effectiveness of malaria control strategies in use [2]. Light microscopy (LM), a standard assay for malaria diagnosis, faces several operational challenges, including limited expertise, power constraints, and time-consuming procedures, leading to delays in reporting results [4, 5]. These challenges necessitate the use of rapid diagnostic tests (RDTs) particularly in rural areas [6]. RDTs typically yield results within 15–20 min, enabling faster clinical decisions compared to LM. Moreover, modern RDTs are sensitive enough to detect low levels of parasitaemia, sometimes even below the threshold detectable by LM [7].
Of the five Plasmodium parasites that are infective to humans, P. falciparum accounts for more than 95% of malaria infections in SSA [8]. Based on this observation, the majority of RDTs distributed in SSA are primarily designed using histidine-rich protein 2 (HRP2) as an antigen target to detect P. falciparum parasites. The HRP2 is a water-soluble protein with a molecular size of 60–105 kDa which is secreted by asexual trophozoite stages of P. falciparum parasites [9, 10]. HRP2 is located on chromosome 8 of the parasite and is secreted in abundance, and circulates in peripheral blood even when the parasites are sequestered in the microvasculature of organs [10, 11]. This makes the protein an ideal target for diagnosis of the P. falciparum parasites. The HRP2 is known to have 85%–90% similarity of nucleotide sequences with the HRP3. This homology renders a cross-reactivity with monoclonal antibodies in HRP2-based RDTs especially in higher parasite densities [12]. Thus, assessing the prevalence of both proteins is crucial as only parasites with dual deletions can be missed by the HRP2-based RDTs [13, 14].
In Tanzania, the National Malaria Control Program (NMCP) initiated the use of RDT for the diagnosis of malaria in 2009. Individuals suspected of malaria cases were required to be confirmed by the HRP2-based RDT before treatments. By 2014, the coverage of RDT across public and private health facilities in Tanzania was about 90%, and RDT is currently the first-line diagnostic assay for the screening of symptomatic malaria in all health facilities [15]. Five different brands of RDTs were approved by the Tanzania Medical Device Authority (TMDA), and in 2015, each outlet in mainland Tanzania sold approximately 40 RDT units per week, with more than 6000 outlets in total [16–18]. Nevertheless, evidence indicates that pfhrp2/3 gene deletions are present in P. falciparum parasites in several countries, including Tanzania [19–21]. Parasites with such deletions are missed by HRP2-based RDTs resulting in false-negative results [20, 22]. Nonetheless, most studies on pfhrp2/3 gene deletions focus on clinical malaria infections and less consideration is given to asymptomatic individuals with malaria.
Reports on the prevalence of individuals with malaria parasites without symptoms are increasing, particularly in areas with high malaria transmission including Tanzania [23–25]. Asymptomatic malaria carriers are infectious and serve as silent precursors to symptomatic infections [23, 25]. Thus, if these silent infections involve parasites with pfhrp2/3 gene deletions, they may also be missed by the HRP2-based RDTs and continue circulating undetected within the population. This selection pressure can increase the frequency of these RDT-evading strains, particularly in regions where RDTs are the primary diagnostic tool, potentially complicating malaria control and elimination efforts.
It is against this background that the present study aimed to assess the prevalence of pfhrp2/3 gene deletions in asymptomatic malaria carriers across villages with varying endemicity in Tanzania. A previous survey of the same cohort reported asymptomatic malaria prevalence using RDT and LM [25]. The current study extends these findings by integrating molecular diagnostics to evaluate pfhrp2/3 deletions, assess RDT performance in asymptomatic infections, and examine associations with demographic factors. By focusing on asymptomatic carriers, this study provides evidence on the contribution of asymptomatic infections to malaria transmission and the potential impact of pfhrp2/3 deletions on RDT-based surveillance and control strategies.
Methods
Geographic coverage and study design
Blood samples were obtained from a cross-sectional community-based study. The study was undertaken from December 2022 to July 2023 and involved 12 villages with different levels of malaria endemicity in Tanzania [25]. Of the twelve villages, eight were from high-malaria transmission hotspots and four were from low-transmission settings. Through multi-stage sampling, two regions (Kigoma and Geita) were purposively selected to represent areas with high malaria cases, and one region (Arusha) was selected to represent areas with low prevalence (Supplementary Table S1). Thereafter, two districts from each region were randomly selected, followed by the selection of two villages per district. The two districts and two villages from Geita region were Nyang’hwale district (Nyangalamila and Kayenze) and Chato district (Ihanga and Rwantaba). From Kigoma region, the districts were Kibondo district (Kumuhasha and Bunyambo) and Kasulu district (Nyamnyusi and Mugombe) while from Arusha, Meru district (Maji ya Chai and Ngurudoto) and Arusha District Council (Bwawani and Themi ya Simba) (Table 1).Table 1. Distribution of collected samples in study sites and isolates selected for detection of pfhrp2 and pfhrp3 gene deletionsqPCR results, n (%)RDT results, n (%)P. falciparum mono-infections selected for hrp2/3 detection, n (%^a^)Site (n)qPCR + Pan + Selected isolatesqPCR + RDT + & qPCR + LM + & qPCR + qPCR +, LM + & RDT + Nyangalamila (276)102 (37.0)8 (2.9)94 (34.1)22 (23.4)25 (26.6)3 (3.2)44 (46.8)Kayenze (327)81 (24.8)4 (1.2)77 (23.5)7 (9.1)11 (14.3)6 (7.8)53 (68.8)Rwantaba (278)116 (41.7)30 (10.8)86 (30.9)6 (7.0)39 (45.3)6 (7.0)35 (40.7)Ihanga (339)126 (37.2)27 (8.0)99 (29.2)6 (6.1)41 (41.4)3 (3.0)49 (49.5)GEITA (1220)425 (34.8)69 (5.7)356 (29.2)41 (11.5)116 (35.6)18 (5.1)181 (50.8)Kumuhasha (293)94 (32.1)21 (7.2)73 (24.9)9 (12.3)29 (39.7)4 (5.5)31 (42.5)Bunyambo (303)113 (37.3)13 (4.3)100 (33.0)30 (30.0)31 (31.0)5 (5.0)34 (34.0)Nyamnyusi (259)124 (47.9)9 (3.5)115 (44.4)35 (30.4)11 (9.6)4 (3.5)65 (56.5)Mugombe (290)112 (38.6)11 (3.8)101 (34.8)25 (24.8)11 (10.9)6 (5.9)59 (58.4)KIGOMA (1145)443 (38.7)54 (4.7)389 (34.0)99 (25.4)82 (21.1)19 (4.9)175 (45.0)Maji ya Chai (279)7 (2.5)1 (0.4)6 (2.2)0 (0.0)5 (83.3)1 (16.7)0 (0.0)Ngurudoto (282)12 (4.3)0 (0.0)12 (4.3)7 (58.3)2 (16.7)0 (0.0)3 (25.0)Bwawani (284)12 (4.2)4 (1.4)8 (2.8)4 (50.0)3 (37.5)0 (0.0)1 (12.5)Themi ya Simba (279)16 (5.7)0 (0.0)16 (5.7)13 (81.3)1 (6.3)0 (0.0)2 (12.5)ARUSHA (1124)47 (4.2)5 (0.4)42 (3.7)24 (57.1)11 (26.2)1 (2.4)6 (14.3)TOTAL (3489)915 (26.2)128 (3.7)787 (22.6)164 (20.8)209 (26.6)38 (4.8)376 (47.8)^a^% of selected isolates; qPCR Quantitative polymerase chain reaction; RDT Rapid diagnostic tests; LM Light microscopy; hrp histidine rich protein
Participant selection and collection of blood samples
Blood samples were collected from individuals aged one year and older residing in selected villages (Supplementary Table S1). A team of researchers visited participants’ households, obtained informed consent, and screened individuals for asymptomatic P. falciparum infection. Asymptomatic individuals were defined as those who were afebrile, body temperature of 37 ℃ or below, had not experienced any malaria symptoms in the preceding 4–5 days, and had not taken antimalarial medication in the past 7 days. Household selection was based on lists provided by village administrative offices, and participants were enrolled using a randomized sampling approach. To ensure even representation, no more than five eligible individuals were selected per household.
Sample collection, malaria diagnosis, and DNA extraction
From each participant, venous blood was collected, followed by malaria diagnosis performed using HRP2-based RDT kits (Bioline™ Malaria Ag P.f/Pan: Abbott, Chicago, USA) and LM, following standard diagnostic guidelines. For molecular analysis, 50 µl of whole blood was spotted onto Whatman 3MM filter paper to prepare dried blood spots (DBS). The DBS cards were dried overnight, individually sealed in plastic bags with desiccants, and transported at ambient temperature to Kangwon National University (KNU), Republic of Korea. Subsequently, DNA was extracted at KNU from a single DBS spot (corresponding to 50 µl of whole blood), using the QIAamp DNA Mini Kits (Qiagen, Hilden, Germany), following the manufacturer’s instructions. DNA was eluted in a final volume of 50 µl.
Molecular diagnosis of P. falciparum infections
To confirm P. falciparum mono-infections and assess DNA quality, a quantitative polymerase chain reaction (qPCR) targeting the small subunit of the 18S rRNA gene was performed using previously published species-specific primers (Forward: ATTGCTTTTGAGAGGTTTTGTTACTTT, Reverse: GCTGTAGTATTCAAACACAATGAACTCAA) and a probe (FAM-CATAACAGACGGGTAGTCAT) [26]. Reactions were carried out in 10 µl volumes containing 5 µl of 2 × Prime Time Gene Expression Master Mix with ROX reference dye (Integrated DNA Technologies, Coralville, USA), 1 µl of gDNA, 5 pmol of each primer and 2.5 pmol of the P. falciparum-specific probe. Amplifications were performed on an AriaMx Real-Time PCR system (Agilent, Santa Clara, USA) under the following conditions; initial polymerase activation at 95 ℃ for 3 min, followed by 40 cycles of denaturation at 95 ℃ for 15 s and annealing/extension at 58 ℃ for 1 min. All reactions were conducted in duplicate. Double-distilled water (DDW) was used as a negative control, and 3D7 strain cultures containing 5% ring-stage parasites, as described previously [27], were used as the positive control for both standard curve generation and estimation of parasite density (relative parasitaemia). Only samples confirmed to be P. falciparum mono-infections were included in the subsequent analysis of pfhrp2 and pfhrp3 gene deletions. To ensure selection of mono-infection cases, samples that tested positive by qPCR but showed non-falciparum antigens (Pan-positive) by RDT were excluded. The limit of detection (LoD) of the qPCR assay, based on the 3D7 18S rRNA gene standard curve, was 5 parasites/µl. Samples below this threshold were considered analytically negative and excluded from pfhrp2 and pfhrp3 gene deletion analysis.
For comparative analyses of parasite density and diagnostic performance, parasite densities estimated by qPCR were categorized into five levels: very low (< 100 parasites/µl), low (100–999 parasites/µl), moderate (1000–4999 parasites/µl), high (5000–99,999 parasites/µl), and hyper parasitaemia (≥ 100,000 parasites/µl), following previously established classifications [28].
Detection of pfhrp2 and pfhrp3 gene deletions
Multiplex qPCR for the detection of pfhrp2 and pfhrp3 gene deletions was confirmed using a laboratory reference strain of P. falciparum 3D7 (both pfhrp2 and pfhrp3 present), Dd2 (pfhrp2 deleted and pfhrp3 present), and HB3 (pfhrp2 present and pfhrp3 deleted) [27]. Gene-specific amplification was conducted using previously published primers and probes. For pfhrp2, the forward primer was GTATTATCCGCTGCCGTTTTTGCC, and the reverse primer was CATCTACATGTGCTTGATTTTCGT. For pfhrp3, the forward was ATATTATCCGCTGCCGTTTTTGCT and the reverse primer was CCTGCATGTGCTTGACTTTCGT. The probes used were FAM-labelled probe (TTCCGCATTTAATAATAACTTGTGTAGC) for pfhrp2, and HEX-labelled probe (CTCCGAATTTAACAATAACTTGTTTAGC) for pfhrp3 [29]. The reaction conditions and volumes used for the multiplex qPCR were identical to those described for the 18S rRNA qPCR assay. Samples with a relative parasite density below 5 parasites/µl were excluded from gene deletion analysis [29]. For samples suspected to have deletions in pfhrp2, pfhrp3, or both genes, amplification was repeated in three independent reactions. A gene was considered deleted only if concordant results were obtained in at least two of the three replicates (Fig. 1).Fig. 1. Flowchart of sample processing and determination of pfhrp2 and pfhrp3 gene deletions. The diagram illustrates the stepwise analysis of field-collected blood samples, beginning with initial malaria diagnosis using RDTs and LM. Samples then underwent molecular confirmation by qPCR to detect Plasmodium falciparum infection. qPCR-positive samples were further analysed using multiplex qPCR to assess the presence or absence of the pfhrp2 and pfhrp3 genes. Proportions of samples retained at each stage of processing are shown, culminating in the final distribution of samples with single and dual gene deletions. This flowchart highlights sample attrition across steps and the relative prevalence of gene deletions among confirmed P. falciparum infections.* qPCR* Quantitative polymerase chain reaction; RDT Rapid diagnostic tests; LM Light microscopy; *pfhrp2 Plasmodium falciparum *histidine rich protein 2; *pfhrp3 Plasmodium falciparum *histidine rich protein 3; p/µl parasites/µl
Statistical analysis
Amplification curves were evaluated using AriaMx 1.8 software (Agilent Technologies), and data visualization was performed using GraphPad Prism 8 (GraphPad Prism Software, San Diego, USA). Parasitaemia was log_10_-transformed, and normality was assessed using the Shapiro-Wilk test. Parasite densities measured by LM and estimated by qPCR were then compared using Pearson’s correlation analysis in SigmaPlot 12 (Systat Software Inc., San Jose, USA). Deletion data were organized in Microsoft Excel (Microsoft, Redmond, USA) using double-entry tables to ensure clarity and minimize entry errors. Unpaired t-tests and one-way analysis of variance (ANOVA) were performed to evaluate associations between log_10_-transformed parasitaemia levels and gene deletion profiles.
Diagnostic performance metrics, including sensitivity, specificity, positive predictive value (PPV), negative predictive value (NPV), and accuracy, were calculated using qPCR as the reference standard with the MedCalc’s diagnostic test calculator (https://www.medcalc.org/calc/diagnostic_test.php). The differences in specificity and sensitivity between RDT and LM were statistically analysed using the McNemar test. All statistical tests were conducted at a 95% confidence interval (CI) and P < 0.05 was considered statistically significant.
Results
Study population
A total of 3489 participants were enrolled in the study, including 2365 (67.8%) from high-transmission villages and 1124 (32.2%) from low-endemicity villages. All participants were tested in the field using HRP2-based RDTs and LM. P. falciparum mono-infections were subsequently confirmed using species-specific qPCR and RDT results. Among participants from high-transmission regions, 1220 (51.6%) were from Geita and 1145 (48.4%) were from Kigoma (Table 1).
Field diagnostic performance of RDTs and LM
To evaluate the performance of HRP2-based RDTs and LM in detecting asymptomatic P. falciparum infections, their sensitivity, specificity, and accuracy were assessed using qPCR as the reference standard. Both mono- and co-infections with P. falciparum were included in the analysis. Nine out of 3489 samples (0.3%) showed Pan-positive results by RDT but were negative by P. falciparum-specific qPCR, suggesting mono-infections with non-falciparum species (Fig. 1). These samples were excluded from further analysis. Detection rates of asymptomatic P. falciparum varied among the three diagnostic methods. A total of 451 (12.9%) participants tested positive by all three methods. RDTs detected 853 (24.4%) participants as positive, whereas LM detected 552 (15.8%) (Fig. 2A). Overall, qPCR identified P. falciparum infections in 915 (26.2%) participants, including 128 (3.7%) that were RDT Pan/Pf double-positive (Fig. 1 and Table 1).Fig. 2. Detection of Plasmodium* falciparum* infections across diagnostic assays and analysis of parasite densities. A Venn diagram showing the overlap in P. falciparum detection among three diagnostic assays: LM, RDT, and qPCR. A majority of samples (n = 451) were positive by all three methods, followed by those detected by both RDT and qPCR. Among samples detected by a single method, qPCR identified the highest number of infections (n = 164), exceeding those detected by RDT or LM alone. B Comparison of relative parasite densities (as determined by qPCR) between diagnostic outcomes. Parasite densities are shown as log_10_ parasites/μl, and normality of the transformed data was assessed using the Shapiro-Wilk test. Samples positive by both RDT and LM showed higher relative parasitaemia (Unpaired t-test, P > 0.05), whereas samples missed by LM had significantly lower parasite densities (P < 0.001). C Correlation analysis between parasitaemia estimated by LM and relative parasite density measured by qPCR (log_10_-transformed). Pearson correlation was used to assess the association (r = 0.267, P < 0.001). D Comparison of relative parasite densities among isolates with different pfhrp2 and pfhrp3 deletion statuses (log_10_-transformed). Mean parasite densities were 5534 ± 17,479 parasites/μl for pfhrp2 + /pfhrp3 + isolates, 2580 ± 9184 for pfhrp2 − /pfhrp3 + , 5347 ± 16,305 for pfhrp2 + /pfhrp3 − , and 1654 ± 3654 for pfhrp2 − /pfhrp3 − isolates. No statistically significant differences were observed among groups (One-way ANOVA, P > 0.05). qPCR Quantitative polymerase chain reaction; RDT Rapid diagnostic tests;* LM* Light microscopy;* pfhrp2 Plasmodium falciparum *histidine rich protein 2; pfhrp3 Plasmodium falciparum histidine rich protein 3
Among the 915 qPCR-confirmed positive cases, the RDT detected 710 [sensitivity: 77.6% (95% CI 74.8–80.3)], whereas LM identified only 492 [sensitivity: 53.8% (95% CI 50.5–57.0)]. The RDT also produced more false positives (143 cases, 5.6%) than LM (60 cases, 2.3%), while LM missed more infections (423 cases, 46.2%) compared with the RDT (205 cases, 22.4%) (Table 2). Overall accuracy was slightly higher for RDTs [90.0% (95% CI 89.0–91.0)] than for LM [86.2% (95% CI 85.0–87.3)]. A McNemar test confirmed that the difference in sensitivity between RDTs and LM was statistically significant (P < 0.001). In terms of specificity, LM showed slightly higher values [97.7% (95% CI 97.0–98.2)] than RDTs [94.4% (95% CI 93.5–95.3)], and this difference was also statistically significant (P < 0.001). PPV was higher for LM [89.1% (95% CI 86.4–91.4)] than RDTs [83.2% (95% CI 80.8–85.4)], whereas the NPV was higher for RDTs [92.2% (95% CI 91.3–93.1)] than for LM [85.6% (95% CI 84.7–86.4)]. Overall, RDTs exhibited "substantial" agreement with qPCR [κ = 0.736 (95% CI 0.710–0.763)], while LM demonstrated only "moderate" agreement [κ = 0.590 (95% CI 0.556–0.624)] (Table 2).Table 2. Diagnostic accuracy of microscopy and RDT using qPCR as reference standardRDTMicroscopyTP (qPCR = 915)710492FP (qPCR negative)14360TN (qPCR = 2574)24312514FN (qPCR positive)205423Sensitivity % (95% CI)77.6 (74.8–80.3)53.8 (50.5–57.0)Specificity % (95% CI)94.4 (93.5–95.3)97.7 (97.0–98.2)PPV % (95% CI)83.2 (80.8–85.4)89.1 (86.4.8–91.4)NPV % (95% CI)92.2 (91.3–93.1)85.6 (84.7–86.4)Accuracy %90.0 (89.0–91.0)86.2 (85.0–87.3)kappa value (95% CI)0.736 (0.710–0.763)0.590 (0.556–0.624)qPCR Quantitative polymerase chain reaction; RDT Rapid diagnostic tests; TP True positive; FP False positive; TN True negative; FN False negative; PPV Positive predictive value; NPV Negative predictive value
Parasite densities and diagnostic concordance
Analysis of diagnostic performance according to parasite density showed that both RDT and LM were more effective in detecting infections with higher parasite densities. However, the detection rate of RDT was significantly higher than that of LM (P < 0.001). Furthermore, RDTs identified a significantly greater number of infections with low parasite densities compared to LM (P < 0.001) (Fig. 2B). As expected, the sensitivity of both RDT and LM increased in parallel with rising parasite densities. Notably, RDT consistently demonstrated higher sensitivity than LM across all parasitaemia categories (Table 3). A comparison between parasite densities measured by LM and relative parasitaemia estimated by qPCR revealed a significant positive correlation (r = 0.267, P < 0.001), indicating a weak but statistically meaningful relationship between the two quantification methods (Fig. 2C).Table 3. Sensitivity of RDTs and microscopy across different parasite densitiesParasite densityNo. of casesSensitivity, n (%)Parasites/μl of bloodqPCRLMRDTVery low (< 100)371124 (33.4)238 (63.2)Low (100–999)283165 (58.3)239 (84.5)Moderate (1000–4999)12792 (72.4)109 (85.8)High (5000–99,999)129106 (82.2)119 (92.2)Hyper parasitemia (≥ 100,000)55 (100.0)5 (100.0)Total915492 (53.2)710 (76.8)qPCR Quantitative polymerase chain reaction; RDT Rapid diagnostic tests; LM Light microscopy
Prevalence and distribution of pfhrp2 and pfhrp3 gene deletions
Only isolates confirmed as P. falciparum positive were analysed for pfhrp2 and pfhrp3 gene deletions. Of the 3489 samples, 915 (26.2%) were positive by qPCR. Among these, 128 (3.7%) were also Pan-positive by RDT, indicating possible mixed-species infections. These samples were excluded from further multiplex qPCR analysis for pfhrp2/3 deletions, leaving 787 (22.6%) isolates confirmed as P. falciparum mono-infections for pfhrp2/3 gene deletion analysis (Fig. 1 and Table 1). Of the 787 isolates selected for deletion analysis, 376 (47.8%) were positive by all three diagnostic methods (RDT, LM, and qPCR), while 164 (20.8%) were positive only by qPCR (Table 1).
Multiplex qPCR identified at least one gene deletion (pfhrp2 or pfhrp3) in 93/787 (11.8%) isolates, including 76 (9.7%) from high-transmission villages and 17 (2.2%) from low-transmission villages. The deletions were unevenly distributed across study sites (Table 4). The single pfhrp2 deletions were detected in 43 (5.5%) isolates, with a mean relative parasitaemia of 101.0 parasites/μl (range: 5.6–25,106). Single pfhrp3 deletions were observed in 31 (3.9%) isolates, with a mean parasitaemia of 177.5 parasites/μl (range: 6.6–3214). Dual pfhrp2/3 deletions were found in 19 (2.4%) isolates, with a significantly lower mean parasitaemia of 27.8 parasites/μl (range: 5.8–96.1). No pfhrp2/3 dual deletions were detected in Maji ya Chai and Ngurudoto (both located in Arusha District, Meru Region), or Rwantaba (Chato District) (Table 4).Table 4. Prevalence and distribution of pfhrp2 and pfhrp3 gene deletions in studied sites and across different demographic parametersRegionDistrictVillage (n of selected isolates)pfhrp2 deletion (n, %^a^)pfhrp3 deletion (n, %^a^)Dual deletion (n, %^a^)GeitaNyang’hwaleNyangalamila (94)6 (6.4)4 (4.3)2 (2.1)Kayenze (77)2 (2.6)1 (1.3)2 (2.6)ChatoRwantaba (86)3 (3.5)1 (1.2)0 (0.0)Ihanga (99)6 (6.1)3 (3.0)1 (1.0)KigomaKibondoKumuhasha (73)2 (2.7)3 (4.1)4 (5.4)Bunyambo (100)9 (9.0)6 (6.0)2 (2.0)KasuluNyamnyusi (115)5 (4.3)3 (2.6)3 (2.6)Mugombe (101)1 (1.0)4 (4.0)3 (3.0)Total in high transmission (n = 745)34 (4.6)25 (3.4)17 (2.3)ArushaMeruMaji ya Chai (6)2 (33.3)0 (0.0)0 (0.0)Ngurudoto (12)3 (25.0)2 (1.7)0 (0.0)Arusha DCBwawani (8)1 (12.5)3 (37.5)1 (12.5)Themi ya Simba (16)3 (18.8)1 (6.3)1 (6.3)Total in low transmission (n = 42)9 (21.4)6 (14.3)2 (4.8)Total in studied regions (n = 787)43 (5.5)31 (3.9)19 (2.4)^a^% of analyzed isolates; pfhrp2 Plasmodium falciparum histidine rich protein 2; pfhrp3 Plasmodium falciparum histidine rich protein 3
Of the 93 isolates with pfhrp2/3 gene deletions, 53 (57.0%) were from female participants, while the remaining 40 (43.0%) were from males. Proportions of deletions were consistently higher among females across all deletion types, though gender-based differences were not statistically significant. When stratified by age, children under 15 years showed a higher prevalence of overall deletions at 59 (63.4%) cases, compared to participants aged 16 and above, who accounted for 34 (36.6%) of gene deletion cases. This age-related difference was statistically significant (P < 0.01), suggesting a higher deletion prevalence in younger individuals (Table 5).Table 5. Comparison of pfhrp2/3 gene deletions by gender and age of participantspfhrp2 deletion (n = 43, %^a^)pfhrp3 deletion (n = 31, %^a^)Dual deletion (n = 19, %^a^)P-valueGender Males19 (44.2)13 (41.9)8 (42.1)P > 0.05 Females24 (55.8)18 (58.1)11 (57.9)Age in years < 516 (37.2)8 (25.8)3 (15.8)P < 0.05 5–1517 (39.5)10 (32.3)5 (26.3) 16–406 (14.0)9 (29.0)4 (21.1) > 404 (9.3)4 (12.9)7 (36.8)^a^% of analyzed isolates; *pfhrp2 Plasmodium falciparum *histidine rich protein 2; pfhrp3 Plasmodium falciparum histidine rich protein 3
Impact of gene deletions on diagnostic outcomes
Overall, parasites with gene deletions had lower parasitaemia than those without deletions. Notably, isolates with dual deletions exhibited significantly lower parasitaemia compared with non-deleted samples (P < 0.05), while no significant difference was observed for single deletions (Fig. 2D). All dual-deletion isolates were undetected by RDTs, yielding false-negative results compared with qPCR. The impact of pfhrp2 and pfhrp3 gene deletions on diagnostic outcomes was evaluated by comparing the proportion of deletion-carrying isolates detected by RDT and LM. Among 43 isolates with a pfhrp2 − /pfhrp3 + genotype, 24 (55.8%) were detected by RDT and 16 (37.2%) by LM (Table 6). In 31 isolates with a pfhrp2 + /pfhrp3 − genotype, 27 (87.1%) were RDT positive and 12 (38.7%) were detected by LM (Table 6). Of the 19 isolates with dual pfhrp2/pfhrp3 deletions, 15 (78.9%) were detected only by LM (Table 6).Table 6. Positivity rates of samples with pfhrp2 and pfhrp3 deletions by RDT and microscopyqPCR (pfhrp2/3 gene deletion)RDT (n, %^a^)Microscope (n, %^a^)PositiveNegativePositiveNegativepfhrp2 − /pfhrp3 + (n = 43)24 (55.8)19 (44.2)16 (37.2)27 (62.8)pfhrp2 + /pfhrp3 − (n = 31)27 (87.1)4 (12.9)12 (38.7)19 (61.3)pfhrp2 − /pfhrp3 − (n = 19)0 (0.00)19 (100)15 (78.9)4 (21.1)^a^% of analyzed isolatesqPCR Quantitative polymerase chain reaction; RDT Rapid diagnostic tests; pfhrp2 Plasmodium falciparum histidine rich protein 2; pfhrp3 Plasmodium falciparum histidine rich protein 3
Discussion
Elimination of malaria in SSA countries is challenged by several factors including the increase in asymptomatic malaria and parasites carrying gene deletions in the pfhrp2 and pfhrp3 genes. Here, we present data from asymptomatic malaria patients sampled from twelve villages with varying malaria endemicity in Tanzania, including eight villages within high malaria transmission areas and four villages within low transmission areas [25]. Although pfhrp2 and pfhrp3 deletions have been documented in symptomatic patients, the present study specifically examines their prevalence among asymptomatic carriers in Tanzania [30, 31]. To ensure reliability of results, samples that failed to amplify in the multiplex qPCR were independently re-tested. Results were considered valid only if at least three replicates yielded concordant results [30].
As suggested by WHO, the performance of malaria diagnostic tools should be frequently assessed, as lowered parasitaemia, particularly in asymptomatic cases, is expected to reduce their efficiency [32, 33]. The present study revealed that LM missed almost 46.2% of infections, whereas RDT missed nearly 22.4% of qPCR-positive P. falciparum infections. These findings are consistent with previous studies on clinical samples, which also showed that LM missed more cases than RDT [34–36]. Overall, the results indicate that RDTs are more sensitive than LM in detecting asymptomatic infections with low parasite densities. False negatives are a significant concern, especially when associated with asymptomatic cases, as failure to detect and treat these individuals allows for continued silent transmission [36]. Although false-negatives in HRP2-based RDTs can result from gene deletions, a large proportion of the current RDT false-negative results (17.5%) were due to other factors, while only 2.6% were attributed to dual pfhrp2 and pfhrp3 gene deletions. Previous studies have shown that submicroscopic malaria infections, characterized by low parasitaemia, largely account for the high false-negative rates of LM and RDT [37, 38]. Similarly, in the present study, a comparison of relative parasitaemia in samples detected by RDT and LM showed that many infections with fewer than 100 parasites/µl were detected by RDT but missed by LM, contributing to its lower sensitivity.
Interestingly, the positivity rates for P. falciparum infections were slightly lower for RDT (853 cases, 24.4%) than for qPCR (915 cases, 26.2%). Among the RDT positives, 143 cases (4.1% of all samples) were false positives. This may be due to the persistence of malaria antigens in the blood even after the parasites have been cleared [35, 39]. The presence of other infections or non-falciparum parasites that can cross-react with RDT may also contribute to the false-positive results [39]. Conversely, 205 RDT-negative cases were positive by qPCR, resulting in a total of 348 (10.0%) discordant cases. McNemar’s test confirmed that the difference in individual-level positivity between RDT and qPCR was highly significant (P < 0.001), suggesting that reliance on RDTs alone may underestimate true infection prevalence, particularly in low-density or asymptomatic infections. In this study, the RDT demonstrated higher sensitivity than LM, detecting a greater proportion of asymptomatic P. falciparum infections and showing clear advantage in identifying true infections. Although the PPV of the RDT (83.2%) was slightly lower than that of LM (89.1%), the higher NPV (92.2%) indicates strong reliability in ruling out infection. The overall accuracy of RDT (90.0%) exceeded that of LM (86.2%), and the higher kappa value (0.736 vs. 0.590) reflects more consistent performance across all cases. Collectively, these findings demonstrate that while both methods are generally reliable, RDTs provide a more sensitive and consistent tool for detecting asymptomatic P. falciparum infections, particularly in field settings where accurate identification of both positive and negative cases is critical. These findings are consistent with other studies conducted elsewhere [35, 36]. The combination of RDT and the molecular qPCR in this study provides evidence that most asymptomatic P. falciparum infections in Tanzania produce sufficient HRP2/3 antigen levels to yield positive results on HRP2-based RDTs.
While the gender-based difference in pfhrp2/3 gene deletion prevalence was not statistically significant, the consistently higher rates observed among females may be influenced by both behavioural and biological factors. In many malaria endemic settings, women, particularly mothers and caregivers, tend to seek health care for themselves and their children more frequently, increasing the likelihood of parasite detection and treatment, which can drive the selection of deletion carrying strains [40]. Additionally, although less studied, hormonal and immunological differences may influence susceptibility and parasite dynamics in females [41]. More notably, our study found a significantly higher prevalence of pfhrp2/3 deletions among children under 15 years of age. This finding aligns with previous research indicating that younger children, having not yet developed sufficient immunity, are more susceptible to higher-density infections and may be more likely to harbour deletion-carrying parasites [42]. In regions where HRP2-based RDTs are heavily relied upon, especially for febrile children, selective pressure may favour the survival and transmission of HRP2-negative strains, contributing to their increased prevalence in both women and young children.
Although the overall prevalence of pfhrp2/3 gene deletions was low (11.8%), a key finding was that both single and dual deletions are circulating among asymptomatic P. falciparum infections in Tanzania. Regions with higher malaria transmission had a greater proportion of deletions (9.7%) than those within low transmission (2.2%). This study aligns with findings from a study conducted in the northern part of Tanzania, where pfhrp2/3 deletions were also observed among asymptomatic malaria patients [19]. In contrast to the previous study, which found no pfhrp2/3 dual deletions, the present study identified dual pfhrp2/3 gene deletions in 2.4% of isolates, all of which showed discordance between RDT and qPCR results [19, 20]. The prevalence of pfhrp2/3 gene deletions, particularly in asymptomatic individuals, highlights the importance of surveying and documenting cases across different stages of malaria infection to understand their role in transmission. In this analysis, Pan-only RDT-positive samples were excluded to minimize the risk of misclassification from mixed-species infections. Nevertheless, it is possible that some of these cases represented P. falciparum with pfhrp2 deletions, which would lead to a modest underestimation of deletion prevalence in our study. This potential source of bias should be considered when interpreting the findings and indicates the importance of including sensitivity analyses in future surveillance efforts.
These findings challenge the WHO master protocol for surveillance of pfhrp2/3 deletions, which emphasizes a health center-based approach [43], thereby increasing the likelihood of missing deletions in asymptomatic individuals. We anticipate that continued use of HRP-based RDTs may exert selective pressure on the parasite population, potentially driving the spread of pfhrp2/3-deleted strains [44]. Considering the increasing prevalence of pfhrp2/3 gene deletions, we recommend that Tanzania’s national malaria diagnostic policy incorporate alternative tools, such as pLDH-based RDTs, which are unaffected by these deletions. Although slightly less sensitive than HRP2-based tests [45], pLDH-based diagnostics offer a viable alternative, particularly in high-risk regions. A combined diagnostic approach using both HRP2 and pLDH-based RDTs could serve as an effective interim solution while long-term policy adjustments are developed.
Conclusions
This study provides important evidence on the performance of HRP2-based RDTs and LM in detecting asymptomatic P. falciparum infections in Tanzania, highlighting both their utility and limitations in the context of emerging pfhrp2/3 gene deletions. While RDTs demonstrated higher sensitivity and better concordance with qPCR than LM, the prevalence of pfhrp2/3 gene deletion-carrying infections remains a concern. Notably, the detection of single and dual pfhrp2/3 deletions, especially in high transmission areas, underscores the silent risk posed by these asymptomatic carriers to malaria control efforts. As a cross-sectional study, these findings are limited by the inability to evaluate temporal trends or causality. In addition, the exclusion of RDT-Pan only infections from molecular deletion analysis may have underestimated the true prevalence of pfhrp2/3 deletions. Despite these limitations, the study emphasizes the need for continued molecular and longitudinal surveillance and calls for more inclusive diagnostic strategies to ensure effective malaria control and elimination in Tanzania.
Supplementary Information
Supplementary Material 1.