The unsung hero: ntnh gene as complementary botulism marker
Sylvia Valdezate, Mónica Valiente, Sergio Díaz-Ramón, Gema Carrasco, María J. Medina-Pascual, Javier Pardos, María Isabel Hurtado, Noelia Garrido, Pilar Villalón, Antonio J. Martín-Galiano

TL;DR
The ntnh gene is a reliable complementary marker for diagnosing botulism, improving detection when traditional methods fail.
Contribution
This study introduces the ntnh gene as a novel complementary diagnostic tool for botulism, demonstrating its high conservation and co-occurrence with botulinum toxin genes.
Findings
The ntnh gene detection confirmed botulism cases in 35.62% of confirmed cases where standard methods were inconclusive.
NTNH sequences showed higher conservation than BoNT sequences, making it a more reliable biomarker.
Two foodborne botulism cases were identified solely through ntnh gene detection despite negative results from other tests.
Abstract
Botulism is a rare but severe neurological disease caused by botulinum neurotoxins (BoNTs). Standard diagnostic methods including the mouse bioassay (SMB) and bont gene type-specific PCR, are often limited by the high genetic diversity among bont subtypes, which can lead to false-negative results. The nontoxin-nonhemagglutinin (ntnh) gene is highly conserved and exclusively co-located with the bont gene complex. Thus, this study evaluates the use of ntnh gene as a complementary diagnostic tool for botulism and assesses its association with BoNT types. The ntnh gene was detected in a prospective BoNT-diagnostic group (n=88) and a BoNT-historical group (n=54). Toxin cluster proteins were identified in GenBank and RefSeq Clostridium proteomes using MMSeqs2. ntnh gene detection reinforced positive results from SMB or bont gene tests in 26 cases (35.62% of the total confirmed cases) of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
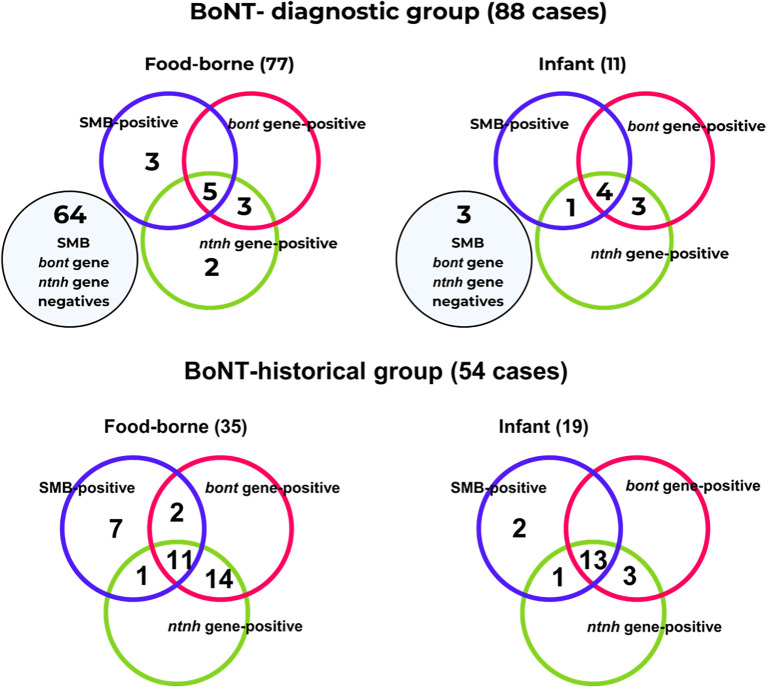 Figure 1
Figure 1 Figure 2
Figure 2| Group | Sex | Age range (median) | Location |
|---|---|---|---|
| BoNT1-diagnostic group (88) | |||
| FB2 (77) | 29 females | 27–75 yr. (52 yr.) | Andalusia (19), Aragon (1), Asturias (3), Canary Islands (3), Cantabria (1), Catalonia (14), Castile and León (13), Castile-La Mancha (2), Galicia (3), Madrid (12), Melilla (1), Murcia (1), Navarre (1), Valencian Community (8), Basque Country (6) |
| 48 males | 43–75 yr. (55 yr.) | ||
| IB3 (11) | 6 females | 1–12 months (4 months) | |
| 5 males | 3–11 months (5 months) | ||
| BoNT1-historical group (54) | |||
| FB2 (35) | 13 females | 14 months-80 yr. (61 yr.) | Andalusia (15), Aragon (3), Baleares (1), Catalonia (13), Castile and León (6), Castile-La Mancha (2), Galicia (1), Madrid (6), Rioja (2), Valencian Community (1), Basque Country (4) |
| 22 males | 19–79 yr. (56 yr.) | ||
| IB3 (19) | 9 females | 20 days-6 months (4 months) | |
| 10 males | 1–9 months (5 months) | ||
- —Ministerio de Ciencia, Innovación y Universidades10.13039/100014440
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBotulinum Toxin and Related Neurological Disorders · Hereditary Neurological Disorders · Neurological disorders and treatments
Introduction
1
Botulism is a potentially fatal neurotoxin-mediated disease characterized by a symmetrical, descending flaccid paralysis affecting voluntary and autonomic muscles. Many patients develop respiratory compromise, frequently requiring emergent airway support, intensive care, and antitoxin administration. Different forms are distinguished by the route of the toxin entry, such as foodborne botulism (FB), intestinal or infant botulism (IB), as well as wound, inhalational, and iatrogenic (cosmetic or therapeutic) botulisms (Rao et al., 2021). The botulinum neurotoxin (BoNT) is one of the most lethal known substances, produced by seven distinct Clostridium species: Clostridium parabotulinum, Clostridium sporogenes, Clostridium botulinum, Clostridium novyi sensu lato, Clostridium argentinense, Clostridium butyricum, and Clostridium baratii. But, over the latter half of the 20th century, the use of C. botulinum to identify any BoNT-producing bacterium gained widespread acceptance (Smith et al., 2023). Based on amino acid variations and antigenic activity, eight toxin types (BoNT/A-BoNT/H) and at least 41 subtypes have been identified in clusters with ha or orfX conformation (Hill et al., 2007).
Laboratory diagnostic criteria for botulism are based on the isolation of BoNT-producing clostridia, the detection of BoNT, or the identification of their encoding genes (bont) https://www.ecdc.europa.eu/en/botulism/facts. A variety of technologies and their derivatives—including cell-based assays, endopeptidase mass spectrometry (Endopep-MS), immunoassays, nucleic acid testing, comparative genomic methods, and electrochemical biosensors—have been developed to detect and characterize the causative agents of botulism (Centurioni et al., 2022; Wang et al., 2025). Diagnosis mainly relies on the mouse bioassay (SMB) and the bont gene amplification (Lindström et al., 2001; Pillai et al., 2024). Diagnosis methods possess serious drawbacks. Specifically, SMB requires animal facilities and the bont type-specific PCR assays are limited by sequence diversity within subtypes (≥23.7%) (Peck et al., 2017; Pillai et al., 2024). Single nucleotide polymorphisms can affect primer or probe matches, potentially resulting in detection failure (Pillai et al., 2024).
Difficulties in detecting BoNT may be mitigated by identifying the nontoxin-nonhemagglutinin (ntnh) gene using a single PCR assay (Raphael and Andreadis, 2007; Fach et al., 2009; Hill et al., 2010; Williamson et al., 2017). All BoNTs are naturally co-expressed with a protective partner, the NTNH protein, forming together the minimal progenitor toxin complex (Lam and Jin, 2015). The role of NTNH in the disease is crucial, as it stabilizes and protects BoNT from the acidic environment and proteases present in the host’s gastrointestinal tract (Gao and Jin, 2024). The ntnh gene is highly conserved and is associated with the BoNT complex across all types, subtypes, and variants, but is absent in non-toxigenic strains (Raphael and Andreadis, 2007; Fach et al., 2009; Hill et al., 2010; Williamson et al., 2017). This study evaluates ntnh gene detection as a strategy to enhance the molecular diagnosis of botulism.
Materials and methods
2
Botulism diagnosis and ntnh gene detection
2.1
Botulism was diagnosed through SMB and BoNT multiplex PCR on chromosomal extracts obtained from prior enrichment of anaerobic cultures of patient stool samples (Centers for Disease Control and Prevention, 1998; Lindström et al., 2001) at the National Centre of Microbiology (Spain). The ntnh gene was studied in suspected botulism cases (Williamson et al., 2017) from a prospective BoNT-diagnostic group (01/2023-08/2025) and retrospectively searched in confirmed botulism cases from a BoNT-historical group (09/2010-12/2022) (Valdezate et al., 2023). This assay also identified the BoNT cluster type (ha or orfX) (Williamson et al., 2017). Patient characteristics (sex, age, and location) were recorded, stratified by botulism type for both groups.
Clinical samples were collected as part of standard patient care and all data were strictly anonymized prior to analysis, so this study was exempted from formal ethical review and approval by the institutional board. However, all procedures were conducted in accordance with the ethical standards of the Declaration of Helsinki, ensuring the protection of patients’ rights, confidentiality, and the ethical principles relevant to public health investigation. Bioassay was performed in accordance with authorization PROEX (no.252.4/21) following institutional and national animal care guidelines.
ntnh gene detection in NCBI genomes
2.2
Clostridium species proteomes (excluding atypical, metagenome-assembled and multi-isolate project ones), were downloaded from NCBI-Genome repository (https://www.ncbi.nlm.nih.gov/datasets/genome/?taxon=2913503) (Accession: 03/09/2025). Toxin cluster proteins were identified within these proteomes using MMSeqs2 (https://search.mmseqs.com) and representative sequences for toxin subtypes (Lindström et al., 2001) besides for NTNH (GenBank accession number: Q45914), H70 (Q9LBR5.1), HA17 (P46083), HA33 (P0DPR0.1), BotR (WP_011948509.1), P47 (WGZ47456.1), ORFX1 (WGZ47458.1), ORFX2 (WGZ4759.1) and ORFX2 (WGZ4760.1). Identity between homologs was calculated following alignment with Muscle v3.8.1551 (https://www.drive5.com/muscle/). Isolate metadata was downloaded from NCBI Biosample (https://www.ncbi.nlm.nih.gov/biosample). NTNH presence in non-Clostridium genomes was explored by BLAST against the RefSeq select database (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Results
3
Study population
3.1
Two distinct sample groups were collected: the prospective BoNT-diagnostic group, comprised 88 samples from 77 foodborne botulism (FB) and 11 infant botulism (IB) cases; and the BoNT-historical group, consisting of 54 samples from 35 FB cases and 19 IB cases (Table 1).
ntnh gene detection
3.2
In the prospective BoNT-diagnostic group, ten FB cases were ntnh positive. Of these, five cases were also positive for SMB and the bont gene, while three cases were only bont gene positive. The remaining two cases were negative for both the SMB and bont tests. Eight cases of IB were ntnh positive: four were also positive for SMB and the bont gene, one was positive only for the SMB, and three were positive only for the bont gene. Regarding the BoNT-historical group, ntnh detection was positive in twenty-six FB cases. Of these, eleven cases were positive for both SMB and the bont gene, one was positive only for the SMB, and fourteen were positive only for the bont gene. Seventeen IB cases were ntnh positive: thirteen were positive for both SMB and the bont gene, one was positive only for the SMB, and three were positive only for the bont gene.
Overall, positive ntnh gene detection reinforced the positive results obtained by either the SMB or bont gene detection in twenty-six cases (35.62%) respect to the positives cases by SMB and/or bont genes tests (confirmed botulism). Conversely, the ntnh gene failed to detect two FB cases in the historical group, despite positive results from both SMB and bont gene tests. Meanwhile, ntnh was detected in two FB cases from the BoNT-diagnostic group, that were negative by both the SMB and bont gene tests. Data are compiled in Figure 1 and in Supplementary Table S2.
Distribution of ntnh detection results across the BoNT-diagnostic group (suspected cases) and the BoNT-historical group (confirmed cases).
Consistent with the established associations between cluster category and bont type (Williamson et al., 2017), our findings showed the following distribution: the ntnh-ha cluster was associated with bont/A (n=5), bont/B (n=45), and bont/F (n=1); the ntnh-orfX cluster was linked to bont/A (n=1) and bont/F (n=2). In one case of bont/A(B), both cluster types were observed.
ntnh gene detection in public NCBI genomes
3.3
The presence, sequence conservation and co-existence of BoNT and NTNH were assessed using public genomes from updated repositories. Of the 3,250 RefSeq and 2,494 GenBank Clostridium genome-based annotated proteomes, 474 and 395, respectively, contained BoNT variants sharing at least 50% identity and 50% alignment length with one BoNT representative. These involved five species previously associated with the BoNT presence. No significant NTNH hits were detected in non-Clostridium species.
Most BoNT cluster proteins showed a strong type association pattern, (i.e. HA proteins with BoNT/A, B, C and D; and OrfX/P47 with BoNT/E, F and G types). The exception was NTNH, which co-existed in 90-100% of strains of all types as appears in color-ranked in the “NTNH” labelled column of Figure 2A.
Genome analysis of bont/ntnh containing genomes. (A) Co-presence of cluster proteins in toxin subtype. (B) Inter-subtype sequence identity of bont and ntnh variants. (C) Venn diagrams showing genomic samples with exclusive and overlapping detection of bont and ntnh genes in RefSeq and GenBank repositories.
Regarding sequence conservation, NTNH and BoNT homologs showed comparable average identity percentage within BoNT subtypes (90.0 ± 10.6% vs 88.3 ± 11.9%, respectively). In contrast, when homologs from different BoNT subtypes were compared (all-against-all), NTNH was far more conserved than BoNT, particularly when different types were involved (67.2 ± 8.9% vs 39.7 ± 8.4%, respectively) (Figures 2B, C). In Figure 2B, the majority of the data points—which represent the averaged inter-subtype identities for BoNT and NTNH between toxin subtype pairs—are located above the diagonal of identity. This distribution demonstrates that, in most instances, NTNH sequences exhibit significantly higher conservation than their corresponding BoNT sequences. This trend becomes even more pronounced when comparing subtypes belonging to different toxin types, further supporting our observation that NTNH remains more conserved across divergent lineages. While, Figure 2C focuses on the reliability of genomic detection regardless of sequence identity. This approach increases the detection rate of toxigenic strains that might otherwise yield negative results for the bont gene.
Furthermore, ntnh was detected in seven and twelve BoNT-negative Clostridium strains in RefSeq and GenBank genome datasets, respectively. These represented 1.8% and 3.0% increments with respect to BoNT-positive strains and involved only three RefSeq-GenBank duplicated cases. Moreover, metadata of these samples indicated a variety of isolation years, countries, isolation sources and sequencing platforms (Supplementary Table S1). Altogether, NTNH detection reinforces the sensitivity of genome-based botulism diagnosis through functional inference.
Discussion
4
The effectiveness of molecular tests in reliably detecting all known BoNT subtype variants causing botulism is constrained by the genetic diversity of BoNTs, even within toxin types (Hill et al., 2010; Peck et al., 2017). To address this constraint on accuracy, the detection of NTNH offers an alternative. The ntnh gene was exclusively found in toxin-producing clostridia and displayed a high co-presence pattern with bont gene. This confirms that the heteromeric stabilized neurotoxin is the actual virulence factor and that ntnh-based diagnostic methods are therefore causal. Despite the ntnh gene is a known “hotspot” for recombination, insertions, and mosaicism (East et al., 1996; Harris et al., 2024), it was found more conserved than BoNT variants and is thus amenable to widespread detection less multiplexing PCR.
C. botulinum has a relatively stable genome except for the bont gene cluster where ntnh gene is included (Hill et al., 2009). Most of these clusters are flanked by insertion sequence elements encoding transposases that may be capable of horizontal gene transfer between C. botulinum Groups I and II, or between species (Smith et al., 2015). Regarding the variability of ntnh and bont genes across different strains, we observed that NTNH sequences within the same BoNT subtype exhibit a high degree of redundancy. Our results show that within intra-subtype comparisons (representing short-term evolution), BoNT is marginally more conserved than NTNH (99.8 ± 0.3% vs. 97.1 ± 5.7% identity), which is expected as the BoNT variant defines the subtype itself. However, this trend reverses during mid-term evolution (different subtypes within the same type), where NTNH shows higher conservation (90.0 ± 10.6% vs. 88.3 ± 11.9%). This conservation gap is greatly enhanced in long-term evolution (different toxin types), where NTNH identity remains significantly higher than BoNT (67.2 ± 8.9% vs. 39.7 ± 8.4%).
NTNH plays a conserved core role in stabilizing and protecting BoNTs across their diverse natural environments (soil, decaying matter, etc.), forming a stable and large toxin complex (Lam and Jin, 2015) that prevents the toxin from being broken down by enzymes (like proteases) before it can infect a host. However, NTNH is also highly flexible, customizing its structure and function to adapt to either the ntnh-ha cluster or the orfX-ntnh cluster (a smaller complex). This suggests that NTNH provides an evolutionarily flexible platform for developing new BoNT variants, enabling it to fit into different environments and attack specific host organisms and tissues. The adaptability of NTNH is thus tied directly to the evolutionary progress of BoNTs (Gao and Jin, 2024; Pitel et al., 2025). Consequently, the NTNH protein acts as an “unsung hero” that not only protects the toxin but actively contributes to its evolutionary success and specialization.
While a wide spectrum of PCR assays has been developed to detect bont genes, only a limited number of studies have incorporated the ntnh gene (Raphael and Andreadis, 2007; Fach et al., 2009; Hill et al., 2010; Williamson et al., 2017). This limited implementation may be due to the fact that ntnh detection does not inherently identify the specific toxin serotype. Previous studies have integrated ntnh gene into various platforms, including real-time PCR (Raphael and Andreadis, 2007; Fach et al., 2009), quantitative PCR (Hill et al., 2010), and conventional PCR (Williamson et al., 2017). The assay designed by Williamson et al. was selected because it also provides epidemiological information regarding the neurotoxin cluster type and has been validated across C. botulinum Group I, C. sporogenes, and C. botulinum Group II (Williamson et al., 2017).
In botulism diagnostics, discrepancies between the detection of BoNT (the protein) and its corresponding genes (bont) are a well-documented event (Centurioni et al., 2022). Here, the application of ntnh detection in diagnosis proved to be a useful complementary target for rapidly confirming positive results from the SMB or bont PCR (26 cases, 35.62%). High correlation was found between the presence of NTNH and BoNT (96.5% of the analyzed genomes).
A potential limitation of ntnh detection is non-expressed toxin genes (Raphael and Andreadis, 2007), although this event does not occur in our study, with the detection of ntnh-ha and ntnh-orfX clusters in a BoNT/A1(B5) case. Furthermore, the ntnh assay provides only indirect evidence for BoNT presence and relies on the functional assumption that BoNT and NTNH are mutually required. While the current criteria for laboratory confirmation of botulism requires explicit positive results from the SMB and/or bont gene identification, there are no established guidelines for interpreting a positive ntnh gene detection when both the SMB and bont tests are negative. This gap in guidelines is highlighted by its occurrence in two patients within BoNT- diagnostic group who were later clinically diagnosed with food-borne botulism.
In conclusion, the detection of ntnh represents a valuable addition to our diagnostic arsenal of botulism. Our findings suggest the need for establishing guidelines on how to interpret a positive ntnh result in the absence of direct toxin and bont gene confirmation to maximize the potential of this target.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Centers for Disease Control and Prevention . (1998) Botulism in the United States, 1899–1996. handbook for epidemiologists, clinicians, and laboratory workers (Atlanta, GA: CDC).
- 2Centurioni D. A. Egan C. T. Perry M. J. (2022). Current developments in diagnostic assays for laboratory confirmation and investigation of botulism. J. Clin. Microbiol. 60, e 0013920. doi: 10.1128/JCM.00139-20, PMID: 34586891 PMC 9020338 · doi ↗ · pubmed ↗
- 3East A. K. Bhandari M. Stacey J. M. Campbell K. D. Collins M. D. (1996). Organization and phylogenetic interrelationships of genes encoding components of the botulinum toxin complex in proteolytic Clostridium botulinum types A, B, and F: evidence of chimeric sequences in the gene encoding the nontoxic nonhemagglutinin component. Int. J. Syst. Bacteriol 46, 1105–1112. doi: 10.1099/00207713-46-4-1105, PMID: 8863443 · doi ↗ · pubmed ↗
- 4Fach P. Micheau P. Mazuet C. Perelle S. Popoff M. (2009). Development of real-time PCR tests for detecting botulinum neurotoxins A, B, E, F producing Clostridium botulinum, Clostridium baratii and Clostridium butyricum. J. Appl. Microbiol. 107, 465–473. doi: 10.1111/j.1365-2672.2009.04215.x, PMID: 19291235 · doi ↗ · pubmed ↗
- 5Gao L. Jin R. (2024). NTNH protein: more than a bodyguard for botulinum neurotoxins. FEBS J. 291, 672–675. doi: 10.1111/febs.17004, PMID: 38009421 PMC 10922118 · doi ↗ · pubmed ↗
- 6Harris R. A. Flint A. Blondin Brosseau M. Weedmark K. Austin J. W. (2024). Complete genomes of Clostridium botulinum type B(F) isolates associated with a 1995 foodborne botulism outbreak from commercial pâté reveals a recombination event disrupting the ntnh gene. Microb. Genom 10, 1169. doi: 10.1099/mgen.0.001169, PMID: 38175697 PMC 10868621 · doi ↗ · pubmed ↗
- 7Hill B. J. Skerry J. C. Smith T. J. Arnon S. S. Douek D. C. (2010). Universal and specific quantitative detection of botulinum neurotoxin genes. BMC Microbiol. 10, 267. doi: 10.1186/1471-2180-10-267, PMID: 20961439 PMC 2973968 · doi ↗ · pubmed ↗
- 8Hill K. K. Smith T. J. Helma C. H. Ticknor L. O. Foley B. T. Svensson R. T. . (2007). Genetic diversity among Botulinum Neurotoxin-producing clostridial strains. J. Bacteriol. 189, 818–832. doi: 10.1128/JB.01180-06, PMID: 17114256 PMC 1797315 · doi ↗ · pubmed ↗