From anonymity to stardom: history of nontuberculous mycobacterial disease in humans
Surendra Kumar Sharma, Vishwanath Upadhyay, Alladi Mohan

TL;DR
This paper reviews the history and evolution of nontuberculous mycobacterial disease in humans, focusing on advances in diagnosis and treatment.
Contribution
The paper provides a comprehensive historical and contemporary analysis of NTM disease management and classification.
Findings
Over 200 NTM species have been identified due to improved diagnostic methods.
Molecular tests now enable rapid and accurate NTM diagnosis.
Treatment decisions depend on clinical relevance and species-specific regimens.
Abstract
Until recently, nontuberculous mycobacteria (NTM) were not considered as human pathogens. Their nomenclature has evolved over several years, until 1971 when finally named as NTM, which is universally accepted now. Because of continuously evolving diagnostic methods, several new species/subspecies of NTM were identified. Presently, nearly 200 NTM species have been reported. Similar to tuberculosis (TB), NTM are known to involve both pulmonary and extrapulmonary organs. Diagnosis of NTM disease is quite cumbersome and is primarily based on their growth characteristics on solid and liquid cultures. Biochemical testing was the mainstay of NTM diagnosis and species identification in the past, which was often erroneous. Recently, molecular tests like line probe assay, targeted, and whole genome sequencing have become available for rapid and accurate diagnosis. Isolation and identification of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
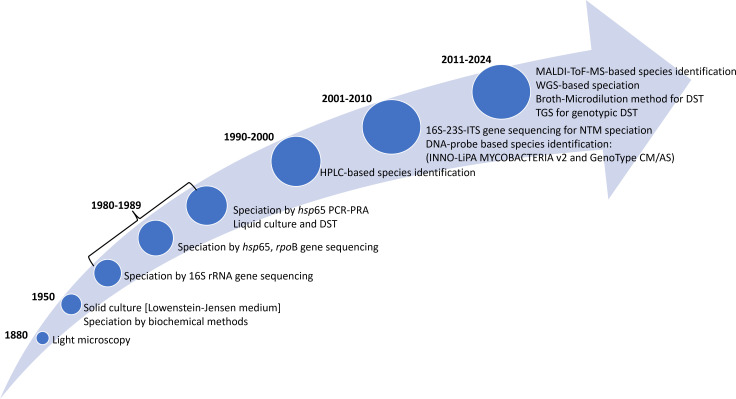 Figure 1
Figure 1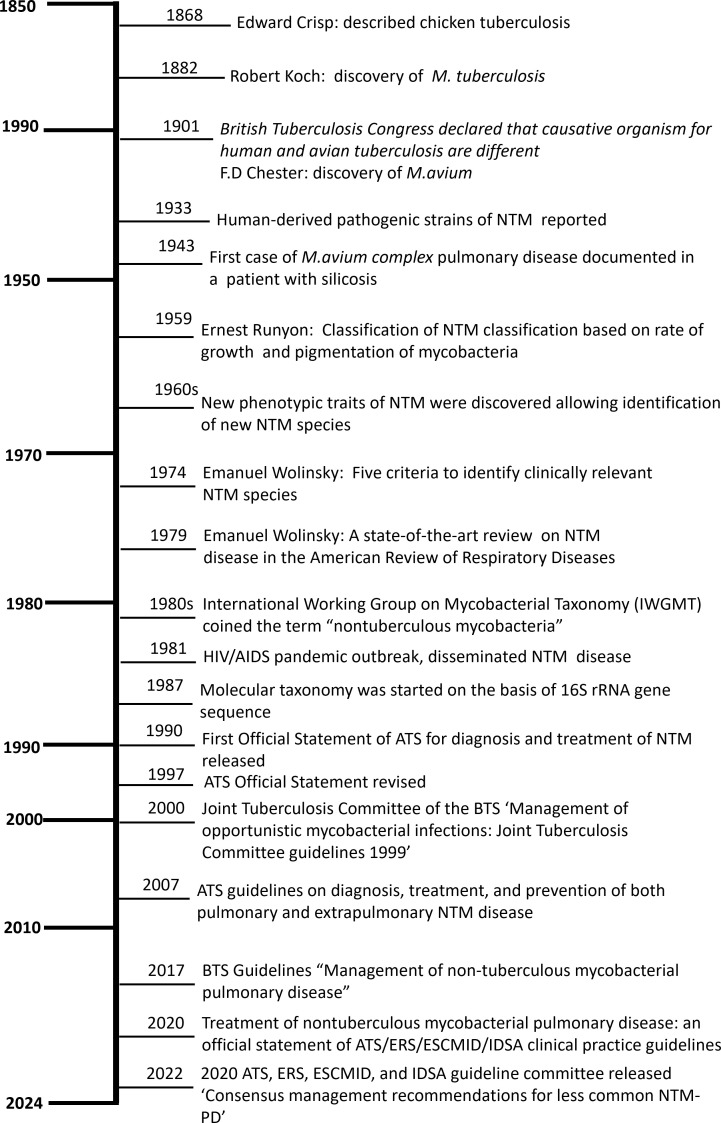 Figure 2
Figure 2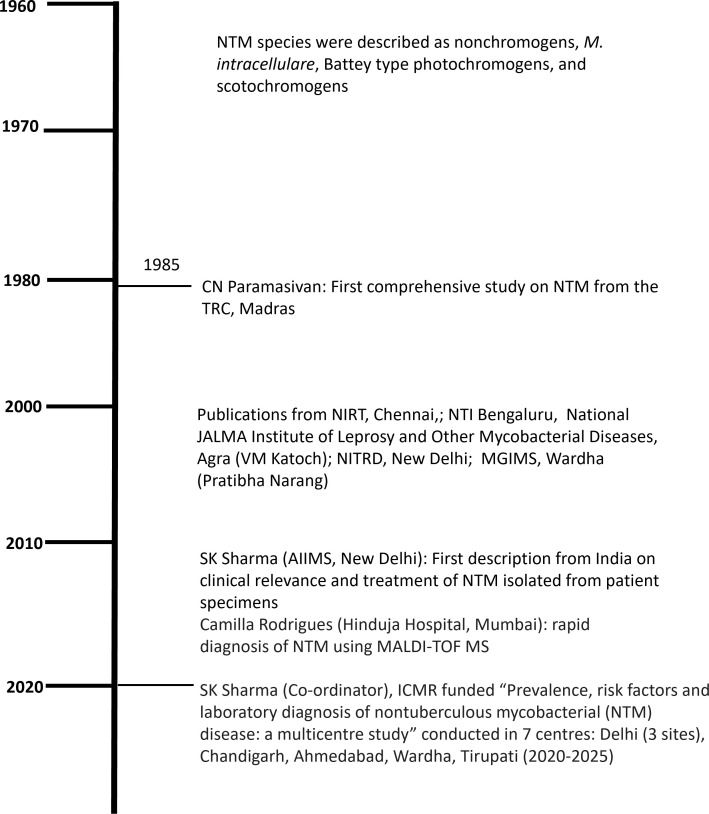 Figure 3
Figure 3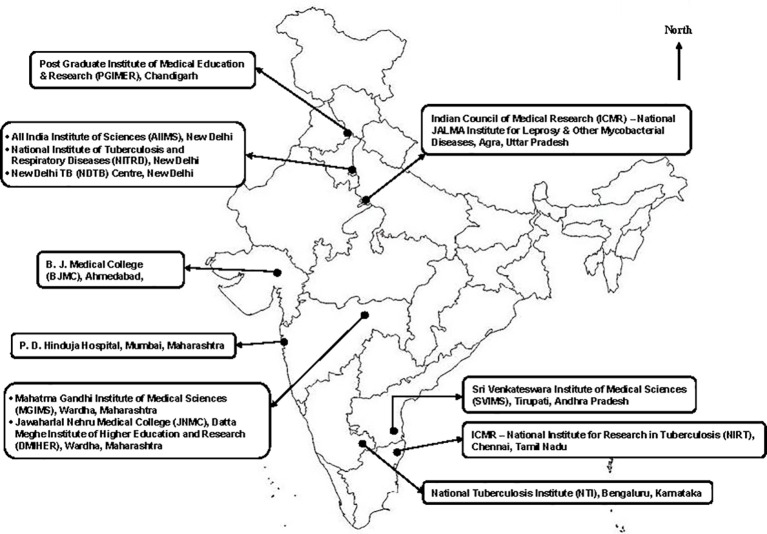 Figure 4
Figure 4| Year | Discovery | Reference |
|---|---|---|
| 1885 | Smegma bacillus | ( |
| 1900 | Report of a rapidly growing AFB (probably | ( |
| 1904 | First report of a chronic injection site abscess caused by AFB | ( |
| 1918 | AFB were noted in the chronic pustular skin lesions of an English soldier who had been wounded in battle and then torpedoed in the North Sea | ( |
| 1925 | Early efforts at classification and differentiation of mycobacteria | ( |
| 1926 | Description and naming of | ( |
| 1933 | Review of literature on NTM from human material | ( |
| 1938 | Description and naming of | ( |
| 1943 | An avian-like bacillus isolated from a man with silicotuberculosis was isolated. This organism was later shown to be | ( |
| 1948 | Description of a new mycobacterial disease of man caused by bacillus now known as | ( |
| 1951 | First reporting the AFB now known as | ( |
| 1952-1953 | An AFB of the M. | ( |
| 1953 | An account of 2 cases of “yellow bacillus” disease and the descriptions of the organism now called | ( |
| 1953 | Non-specific nature of the weak skin reactions to tuberculin PPD seen in some populations might be | ( |
| 1954 | Correlation of the known facts about the relationship between human pulmonary disease, NTM, and provided the first working classification of the organisms | ( |
| 1959 | Runyon originally proposed that the strains virulent for chickens and rabbits be called | ( |
| 1990 | Official Statement of the ATS 1990 | ( |
| 1997 | Official Statement (revised) of the ATS 1997 | ( |
| 1999 | Joint Tuberculosis Committee of the BTS guidelines 1999 | ( |
| 2007 | Official Statement of the ATS 2007 | ( |
| 2016 | The US Cystic Fibrosis Foundation and European Cystic Fibrosis Society guidelines consensus recommendations for the management of NTM in individuals with cystic fibrosis | ( |
| 2017 | BTS guidelines 2017 | ( |
| 2020 | ATS/ERS/ESCMID/IDSA guidelines | ( |
| 2022 | Consensus management recommendations for less common non-tuberculous mycobacterial pulmonary diseases | ( |
| Domain | Nontuberculous mycobacteria |
|
|---|---|---|
| Reservoir and transmission | Environmental water/soil/biofilms with exposure-driven acquisition; person-to-person spread is not dominant | Human reservoir with predominant airborne transmission and high adaptation to human hosts |
| Typical host risk profile | Structural lung disease, impaired airway clearance, elderly women with thoracic dysmorphisms, and cellular immune defects, including anti–IFN-γ autoantibodies and Mendelian susceptibility to mycobacterial disease variants | Severe disease is frequent in immunocompetent hosts; risk amplified by human immunodeficiency virus and iatrogenic immunosuppression |
| Innate recognition and activation | PRR signaling (TLR2-centered) with heterogeneous IL-12–IFN-γ coupling and variable macrophage activation by species and host | Robust PRR signaling and IL-12–IFN-γ coupling drive strong Th1 responses |
| Intracellular fate | MAC blocks phagosome-lysosome fusion; | Phagosomal arrest with canonical survival programs promotes intracellular persistence and necrotizing pathology |
| Granuloma morphology | Often chronic, fibrotic, and less caseating with prolonged host–pathogen equilibrium | Frequently, caseating granulomas with cavitation and high bacillary burden |
| Immune evasion | GPL masking of TLR ligands, phagolysosome blockade, cytokine modulation, phagosomal damage/escape, and biofilm-mediated tolerance, varying by species/morphotype | Potent inhibition of phagosome maturation and manipulation of host inflammation and cell death |
| Autophagy/xenophagy | Frequent flux impairment and non-canonical routing; TFEB activation (amiodarone, rufomycins, resveratrol analog V46), trehalose–PIKFYVE–TFEB, and AMPK activation (metformin) restrict intracellular bacilli; macrolides can alkalinize lysosomes and oppose killing in | Xenophagy is engaged, but acidification and degradation curtailed; vitamin D–CAMP, autophagy initiation, and TFEB activation reduce burden in models |
| Microbiome influence | Gut–lung axis and airway microbiota modulate immune tone and treatment responsiveness in NTM-PD | Microbiome effects are reported but less central to classical transmissibility and caseating pathology paradigms |
| Extrapulmonary pathogenesis | Direct inoculation and medical instrument-associated biofilms drive skin/soft-tissue, lymphatic, and musculoskeletal disease; hematogenous dissemination in IL-12–IFN-γ axis defects or anti–IFN-γ autoantibodies produces multi-organ disease with high organism load | Hematogenous and lymphatic spread frequently produce caseating granulomas in lymph nodes, bones, and viscera with strong necrotizing Th1 pathology, even without device biofilms |
| Drug (Year of discovery)/ Route of administration | Mechanism of action | Year of first use | Remarks in the context of NTM | Current status in NTM treatment | Reference for first use in NTM diseases | |
|---|---|---|---|---|---|---|
| TB or other infections | NTM | |||||
| Streptomycin (1943)/ | Protein synthesis inhibitor (30S ribosomal subunit) | 1943 (TB) | 1947 | First antibiotic for mycobacterial diseases | Historically approved | ( |
| PAS (1902)/ | Folate synthesis inhibitor | 1943 (TB) | 1957 | Early antimycobacterial, repurposed from TB; limited current use | Historically approved | ( |
| Isoniazid (1912)/ | Inhibits mycolic acid synthesis | 1943 (TB) | 1957 | Widely tried, effective for a few NTM species | Historically approved | ( |
| Clofazimine (1954)/ | Disrupts bacterial respiration & ion transport | 1962 (Leprosy) | 1964 | Adjunct in refractory NTM | Standard-of-care | ( |
| Ethionamide (1956)/ | Inhibits mycolic acid synthesis | 1965 (TB) | 1965 | Occasionally used in resistant cases | Historically approved | ( |
| Rifampicin (1957)/ | Inhibits DNA-dependent RNA polymerase | 1967 (TB) | 1969 | Backbone of MAC therapy | Standard-of-care | ( |
| Ethambutol (1961)/ | Inhibits arabinosyl transferase | 1969 (TB) | 1969 | Used in MAC combinations | Standard-of-care | ( |
| Doxycycline (1958)/ | Protein synthesis inhibitor (30S ribosomal subunit) | 1958 (Bacterial infections) | 1981 | Used off-label for rapidly growing mycobacteria, particularly | Standard-of-care | ( |
| Amikacin (1972)/ | Protein synthesis inhibitor (30S ribosomal subunit) | 1981 (TB) | 1981 | For severe and refractory NTM pulmonary disease. | Standard-of-care | ( |
| Cefoxitin (1972)/ | Cell wall synthesis inhibitor (β-lactam) | 1974 (Bacterial infections) | 1990 | Used in combination therapy for | Standard-of-care | ( |
| Ciprofloxacin (1983)/ | DNA gyrase inhibitor | 1986 (Broad-spectrum infections) | 1986 | Early fluoroquinolone for NTM; off-label use | Standard-of-care | ( |
| Rifabutin (1975)/ | Inhibits DNA-dependent RNA polymerase | 1990 (TB) | 1990 | Alternative rifamycin for MAC & | Standard-of-care | ( |
| Clarithromycin (1980)/ | Protein synthesis inhibitor (50S ribosomal subunit) | 1990 (Respiratory infections) | 1990 | First effective macrolide for MAC/NTM | Standard-of-care | ( |
| Azithromycin (1980)/ | Protein synthesis inhibitor (50S ribosomal subunit) | 1992 (Respiratory infections) | 1992 | Preferred macrolide for NTM treatment; better tolerance | Standard-of-care | ( |
| Imipenem (1985)/ | Cell wall synthesis inhibitor (β-lactam) | 1995 (Bacterial infections) | 1995 | Rescue therapy in drug-resistant NTM; off-label use | Standard-of-care | ( |
| Levofloxacin (1991)/ | DNA gyrase inhibitor | 1996 (Broad-spectrum infections) | 1996 | Alternative fluoroquinolone for NTM; off-label use | Standard-of-care | ( |
| Linezolid (1996)/ | Protein synthesis inhibitor (50S ribosomal subunit) | 2001 | 2001 | Salvage therapy for refractory NTM; off-label use | Standard-of-care | ( |
| Moxifloxacin (1996)/ | DNA gyrase inhibitor | 2003 | 2003 | Improved fluoroquinolone for select NTM; off-label use | Standard-of-care | ( |
| Bedaquiline (2005)/ | Inhibitor of ATP synthase | 2006 (TB) | 2012 | Experimental for NTM, especially for | Phase 2 and 3 trial | ( |
| Tigecycline (1999)/ | Protein synthesis inhibitor (30S ribosomal subunit) | 2005 (Broad-spectrum infections) | 2007 | Salvage therapy for multidrug-resistant NTM; off-label | Standard-of-care | ( |
| Tedizolid (2005)/ | Protein synthesis inhibitor (50S ribosomal subunit) | 2017 (Gram-positive infections) | 2017 | Less toxic alternative to linezolid; off-label use | Standard-of-care | ( |
| GM-CSF (1980s)/ | Immunomodulator; enhances alveolar macrophage activation | 2018 (Immunotherapy) | 2018 | Off-label, adjuvant immunotherapy in refractory NTM | Approved – Phase IV trial(off-label) | ( |
| Amikacin liposome inhalation suspension (ALIS) (2018)/ | Protein synthesis inhibitor (30S ribosomal subunit) | 2018 (NTM) | 2018 | Approved inhaled formulation for refractory MAC | Approved, Phase III/IV trial | ( |
| Bacteriophage therapy (1920s)/ | Lytic bacteriophages targeting mycobacteria | 2019 (Bacterial infections) | 2019 | Experimental; compassionate use in MDR NTM | Investigational, compassionate use/case reports | ( |
| Inhaled clofazimine (1954)/ | Disrupts bacterial respiration & ion transport | 2020 (Inhalation) | 2020 | Repurposed formulation for lung targeting in NTM | Investigational – Phase 2 trial | ( |
| Omadacycline (2014)/ | Protein synthesis inhibitor (30S ribosomal subunit) | (2021) Skin and respiratory infections | 2021 | Off-label; exploratory use against rapidly growing mycobacteria | Phase 2 trial | ( |
| Year | Name of biomarkers | Compartment | Comments | References |
|---|---|---|---|---|
| 1969-2000s (discovery and structural and functional studies) | Anti-GPL-core IgA and IgG | Blood plasma/serum |
• Rapid test • Both diagnostic and monitoring marker • Minimally invasive • Used for limited NTM species • Variable diagnostic cut-offs • Broad range of sensitivity and specificity | ( |
| 2020 | Human microRNAs: hsa-miR-484, | Total RNA (human) |
• Serum-specific miRNA signature • Host antibacterial pathway modulators • Costly and requires expertise • Requires to be validated in a large cohort for various NTM species | ( |
| 2022 | 2,3,5,8-tetramethyl-decane | Breath |
• Study done in CF patients only • Costly and requires expertise • Requires to be validated in a large cohort for various NTM species | ( |
| 2023 | CRP | Serum |
• Prognostic marker • Widely available and cost-effective • Nonspecific | ( |
| 2024 | HBA1, HBA2, HBD and METTL7B genes | Genomic DNA |
• New markers • Requires expertise • Costly • Requires to be validated in a large cohort for various NTM species | ( |
| 2024 | Methionine, hypoxanthine | Urine |
• Non-invasive • Experimental stage • Costly and requires expertise • Requires to be validated in a large cohort for various NTM species | ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycobacterium research and diagnosis · Tuberculosis Research and Epidemiology · Diagnosis and treatment of tuberculosis
Introduction
1
Nontuberculous mycobacteria (NTM) are mycobacteria other than Mycobacterium tuberculosis (the causative agent of tuberculosis [TB]) and Mycobacterium leprae (the causative agent of leprosy). These mycobacteria have also been referred by several other names, including ‘paratubercle bacilli’, ‘pseudotubercle bacilli’, ‘tuberculoid bacilli’, ‘unclassified’, ‘anonymous’, ‘atypical’, ‘environmental’, ‘opportunistic’, and mycobacteria other than M. tuberculosis complex (MOTT) (Turenne, 2019). Though NTM had existed for a long time, because of similarity with Mtb and lack of clarity about their nomenclature, these were neglected. In 1981, with the advent of human immunodeficiency virus (HIV) infection, these organisms came into limelight and importance of localised and disseminated NTM disease was recognized.
Historically the first account of these ‘atypical’ mycobacteria can be traced back to the 19^th^ century, when “tuberculosis “in chickens was described in 1868 (Nontuberculous mycobacteria (overview)). By 1890, it was discovered in the laboratory that the organism was different from M. tuberculosis and later was identified to be Mycobacterium avium and it was realized by then that these organisms did not cause disease in humans (Nontuberculous mycobacteria (overview)). In 1926, M. marinum infection in salt water fish was reported (Aronson, 1926). Later in early 1930s, atypical mycobacteria were recognized to cause diseases in humans (Branch, 1933). In 1938, da Costa Cruz (1938) described M. fortuitum (da Costa Cruz, 1938). In 1943, the first case of atypical mycobacteria-associated pulmonary disease due to M. avium complex (MAC) was described in a patient with underlying silicosis (Runyon, 1959). Soon, it was realized that the atypical mycobacteria-associated lung disease did not respond to anti-TB drugs (Nontuberculous mycobacteria (overview)).
The history of NTM is marked by profound knowledge gaps, both globally and specifically in India. Historically, diagnostic limitations have been a primary obstacle; reliance on smear microscopy for TB diagnosis meant countless NTM cases were likely misidentified as “smear-positive, culture-negative TB,” making their true prevalence fundamentally unknowable.
With the exception of Buruli ulcer in specific regions of the world globally (7), NTM was never subjected to systematic international surveillance, leaving vast regions of Asia, Africa, and South America as historical blind spots. Early data often failed to differentiate between NTM species, rendering it uninterpretable. A central dogma—that NTM was only acquired from the environment—has recently been challenged, raising the critical and unanswered question of whether unrecognized human-to-human transmission has always occurred. This is compounded by a major therapeutic gap, as the development of drugs specifically for NTM has seldom been done.
The following description provides an account of the evolution of historical aspects regarding NTM.
Historical account: global perspective
2
A narrative of various historic events leading to our present knowledge regarding these atypical mycobacteria is detailed in Table 1, Figures 1, 2 (Alvarez and Tavel, 1885; Marzinowski, 1900; Ophüls, 1904; Cobbett, 1918; Wilson, 1925; Aronson, 1926; Branch, 1933; da Costa Cruz, 1938; MacCallum et al., 1948; Cuttino and McCabe, 1949; Norden and Linell, 1951; Tarshis and Frisch, 1952a; Tarshis and Frisch, 1952b; Tarshis and Frisch, 1952c; Buhler and Pollak, 1953; Gibson, 1953; Karlson and Feldman, 1953; Lominski and Harper, 1953; Moore and Frerichs, 1953; Palmer, 1953; Timpe and Runyon, 1954; Runyon, 1959; Runyon, 1959; Anonymous, 1990; Anonymous, 1997; Subcommittee of the Joint Tuberculosis Committee of the British Thoracic Society. Management of opportunist mycobacterial infections: Joint Tuberculosis Committee Guidelines 1999, 2000; Griffith et al., 2007; Haworth et al., 2017; Daley et al., 2020; Lange et al., 2022; Ravindran et al., 2025). For a long time, clinical isolates of these atypical mycobacteria from pulmonary specimens were often considered as contaminants without any clinical significance (Runyon, 1959). During the 1940s and 1950s, these acid-fast organisms were considered as saprophytes, unlike Mtb, they were not considered pathogenic in guinea pigs (Runyon, 1959) as per the dogmatic belief that acid-fast bacilli (AFB) were unable to produce disease in guinea pigs was ipso facto a saprophyte (Runyon, 1959; Cummins and Williams, 1933). Repeated isolation of AFB from sputum specimens, on culture and later on isolation from the resected lung specimen from the same patient, that were non-virulent to guinea pigs, were documented (Runyon, 1959). This observation along with the failure to find alternative etiological agents on bacteriologic and histopathologic investigations provided a strong clue that these strains produced disease in humans. In 1959 Ernst Runyon (Runyon, 1959) wrote “The guinea pig no longer sits alone on the throne of decision as to pathogenicity of acid-fast bacilli for man; there is the mouse, and also an empty chair. Mice have been found to be more susceptible than guinea pigs to some of the anonymous mycobacteria. The “empty chair” pertains to the lack of any known animal host in which certain strains can establish progressive infection, although these bacteria evidently have been involved in human disease” (Runyon, 1959).
Timeline of various laboratory techniques for NTM isolation and identification of its species. rRNA, ribosomal ribonucleic acid; DNA, deoxyribonucleic acid; DST, drug-susceptibility testing; HPLC, high performance liquid chromatography; PCR-PRA, polymerase chain reaction restriction fragment length polymorphism; CM/AS, common mycobacteria/additional species; MALDI TOF MS, matrix assisted laser desorption time of flight mass spectrometry; ITS, internal transcribed spacer; WGS, whole genome sequencing; TGS, targeted gene sequencing.
History of nontuberculous mycobacteria: global perspective. NTM, nontuberculous mycobacteria; IWGMT, International Working Group on Mycobacterial Taxonomy; HIV, human immunodeficiency virus; AIDS, acquired immunodeficiency syndrome; rRNA, ribosomal ribonucleic acid; ATS, American Thoracic Society; BTS, British Thoracic Society; ERS, European respiratory Society; ESCMID, European Society of Clinical Microbiology and Infectious Diseases; IDSA, Infectious Diseases Society of America.
Thorough preparations for this survey were made. Particularly, the efforts by William H. Feldman circulated outlines for the study of these bacteria in 1953 are noteworthy. The Veterans Administration (VA) - National Tuberculosis Association (NTA) Survey of Mycobacterial Infections initiated in 1954 by a subcommittee of the VA Committee on Bacteriology with W.E. Dye as Chairman provided a great impetus to the understanding of the clinical significance of various mycobacteria (Timpe and Runyon, 1954; Runyon, 1955; Runyon, 1959). Ernst Runyon, the Chairman of VA subcommittee, had obtained the collaboration of the American Trudeau Society of the NTA in the distribution of a questionnaire and request for cultures (Runyon, 1955). In this survey (Runyon, 1955) of the assembled cultures, established Groups I (photochromogens), II (scotochromogens), III (nonphotochromogens) and IV (rapid growers) as useful features for differentiating the anonymous strains from M. tuberculosis. Aforementioned grouping of NTM was used for a long time and now is of historical significance. This grouping has been replaced by molecular methods for speciation and sub-speciation. In light of this survey, it became evident that clinical, radiographic and histopathologic studies are insufficient to differentiate pulmonary diseases due to various distinct types of anonymous (“atypical”) acid-fast organisms from each other and from TB; bacteriologic identification is essential. NTM from Groups I and III were noted to be the most important pathogens; geographical variations were evident. In the 1960s, studies from Mexico (Bojalil et al., 1962), USA (Wayne, 1966), and Japan (Tsukamura, 1967) reported new phenotypic traits allowing the identification of novel mycobacterial species, as well as improved understanding of epidemiology and pathogenicity of these organisms.
Wolinsky (1981) had addressed the issue of colonization versus infection in the context of these atypical organisms. He stated that while M. tuberculosis, a definitive pathogen, is rarely recovered from culture except in association with disease; NTM that are plentiful in the environment are often encountered as colonizers, or agents producing infection but not recognizable disease. Further, their propensity to cause opportunistic infection in previously damaged lung tissue makes it difficult to distinguish new disease from old. Wolinsky (1981) stated that the decision whether to treat disease caused by these atypical organisms or not rests with the physician and mentions following helpful facts to decide treatment. These include (i) moderate to heavy growth of the mycobacteria from sputum specimens (light growth may occur from sterile body fluids or tissues); (ii) repeated isolation of the same organism (in pulmonary disease); (iii) site of origin of the positive specimen; (iv) species/subspecies of mycobacteria identified; and (v) host risk factors/predisposing conditions. Wolinsky (1981) suggested that even when disease is determined it may be prudent to withhold treatment unless the disease progresses rapidly. This is still relevant presently as after isolation a greater emphasis is given to clinical relevance of isolated NTM species/subspecies (Daley et al., 2020).
In a 1979, in a state-of-the-art review (Wolinsky, 1979) titled, ‘Nontuberculous mycobacteria and associated diseases’ in the American Review of Respiratory Disease (an official publication of the American Thoracic Society [ATS], now named American Journal of Respiratory and Critical Care Medicine) Emanuel Wolinsky changed the perception about NTM and their potential to produce human diseases. By this time, 40 NTM species had been identified with the help of phenotypic and biochemical characteristics. This review article (Wolinsky, 1979) described virulence potential of these mycobacteria, and since then, this topic had been explored extensively by several researchers across the globe with a phenomenal momentum. The incidence of disseminated MAC rose dramatically in the pre-cART era, particularly in patients with CD4+ T-lymphocyte counts <50 cells/mm³ but declined substantially following the introduction of effective combination antiretroviral therapy and routine primary prophylaxis. In high-income settings, disseminated MAC is now concentrated in patients with advanced, untreated HIV infection and in those with poor treatment adherence. In contrast, pulmonary NTM disease has emerged as a major problem in older, structurally abnormal lungs, even in HIV-negative populations. Consistent with these trends, current guidelines recommend macrolide or rifabutin prophylaxis for patients with advanced immunosuppression (e.g. per NACO) alongside early ART and careful evaluation of any mycobacterial isolate to distinguish disseminated MAC from TB (Panel on opportunistic infections in adults and adolescents with HIV. Guidelines for the prevention and treatment of opportunistic infections in adults and adolescents with HIV: recommendations from the centers for disease control and prevention, the national institutes of health, and the HIV medicine association of the infectious diseases society of America). In the late 1980s, the International Working Group on Mycobacterial Taxonomy (IWGMT) coined the term “nontuberculous mycobacteria (NTM)” which has been widely accepted and used to date by various microbiologists and clinicians worldwide (Wayne, 1988). Several researchers are working either singly or in collaboration mode, nationally or at the international level. The speed and accuracy of mycobacterial species identification showed a significant improvement with the application of high-performance liquid chromatography (HPLC) for documenting the chromatographic profile of mycolic acid extracted from the bacterial cell wall and subsequent introduction of various molecular methods such as DNA probes and gene sequencing. After the advent of whole genome sequencing in 1987 the International Committee for Systematic Bacteriology defined that a species includes strains with ~70% or more deoxyribonucleic acid (DNA)-DNA homology and with 5 °C or less difference in melting temperature of the homologous genomic DNA and hybrid DNA (ΔTm) providing the precise difference between genetically closed NTM species (Wayne, 1988). Conventionally NTM are divided into slowly growing mycobacteria (SGM; ≥7 days) and rapidly growing mycobacteria (RGM; <7 days) (Runyon, 1959). In molecular taxonomy, this has been attributed to the differences of 2–4 nucleotides in helix 18 of the hypervariable region B of the 16S ribosomal ribonucleic acid (rRNA) gene (Pereira et al., 2020). With the availability of gene sequencing for NTM speciation and a wide range of databases of mycobacterial species, in the last three decades the number of NTM species expanded more than three times (Turenne, 2019). Although nearly 200 NTM species have been described, only a few of these are pathogenic to causes diseases in humans (Turenne, 2019).
Friedmann’s discovery of M. chelonae (“chelonae”, Latin for “of a turtle”) in the lungs of two sea turtles in the early 20^th^ century marked the beginning of the history of the major pathogenic species of rapidly growing mycobacteria (RGM) (Cobbett, 1918; Brown-Elliott and Philley, 2017). The closely related M. abscessus was initially identified as the cause of a human skin and soft tissue infection in a patient who had several lower extremities soft tissue abscesses nearly 50 years later (Moore and Frerichs, 1953). Another RGM species, M. fortuitum (formerly M. ranae), was originally recovered from frogs in 1905. However, in 1938, da Costa Cruz (1938) named M. fortuitum to an isolate considering it a new mycobacterial species isolated from a patient with a skin abscess from local vitamin injections site. Subsequently, both organisms were proven to be the same, with the illegitimate name M. fortuitum retained as the species name. Early taxonomic studies based on phenotypic analysis concluded that the two species M. fortuitum and M. chelonae were composed of several “subspecies” (i.e., M. chelonae subsp. chelonae and M. chelonae subsp. abscessus) or biovars (M. fortuitum bv. fortuitum, M. fortuitum bv. peregrinum, and M. fortuitum third biovariant complex). In early 1980s, DNA-DNA hybridisation and 16S rRNA gene analysis revealed that M. chelonae and M. fortuitum biovars and subspecies are in fact different species. Until 1992, M. abscessus was considered as a subspecies of M. chelonae, however, after extensive genomic analysis in previous decade, M. chelonae and M. abscessus were designated as a different species (Kusunoki and Ezaki, 1992).
In view of increasing isolation rates and the establishment of various NTM as pathogens for humans, the first Official Statement of the ATS for diagnosis and treatment of NTM was released in 1990 (Anonymous, 1990) which was further revised in 1997 (Anonymous, 1997). The Joint Tuberculosis Committee of the British Thoracic Society (BTS) came up with an evidence-based document namely ‘Management of opportunist mycobacterial infections: Joint Tuberculosis Committee guidelines 1999’, which was published in the year 2000 (Subcommittee of the Joint Tuberculosis Committee of the British Thoracic Society. Management of opportunist mycobacterial infections: Joint Tuberculosis Committee Guidelines 1999, 2000). In 2007, the ATS issued for the first time landmark guidelines that addressed the diagnosis, treatment, and prevention of both NTM pulmonary disease (NTM-PD) and extra-pulmonary NTM disease (EP-NTM) (Griffith et al., 2007). Thereafter, in 2016, the consensus recommendations for the screening, investigation, diagnosis and management of NTM-PD in individuals with CF were issued by the US Cystic Fibrosis Foundation and European Cystic Fibrosis Society (ECFS) (Floto et al., 2016). Following that, two new guidelines, BTS 2017 (Haworth et al., 2017) and the ATS/European Respiratory Society (ERS)/European Society of Clinical Microbiology and Infectious Diseases (ESCMID)/Infectious Diseases Society of America (IDSA) 2020 (Daley et al., 2020), were released. The last two guidelines (Haworth et al., 2017; Daley et al., 2020) have exclusively focused on NTM-PD in people without HIV/AIDS or cystic fibrosis. Both guidelines endorsed the clinical, radiographic and microbiological criteria defined in the ATS (2007) guideline (Griffith et al., 2007) for diagnosing NTM-PD. Subsequently, the panel members of the 2020 ATS, ERS, ESCMID, and IDSA Guideline Committee also released ‘Consensus management recommendations for less common NTM-PD’ in 2022 (Lange et al., 2022).
Nontuberculous mycobacterial diseases in India
3
In India, there has been no coherent historical time-line of NTM prevalence due to a lack of surveillance data and a near-total absence of research mapping environmental reservoirs in water or soil. A significant policy gap meant NTM remained invisible to the public health system. Unlike TB, India has historically had no national program or country-specific guidelines for NTM diagnosis and treatment, forcing clinicians to rely on international standards that may be ill-suited for the local context of drug availability and cost. The historical interaction between human, animal, and environmental NTM remains completely unexplored. The “One Health” perspective—linking human, animal, and environmental health—is a recent development (Hegde et al., 2024) The historical interaction and potential transmission between animals and humans in the Indian context is completely unknown.
A few studies (Narain et al., 1968; Rao and Kotian, 1976; Mustafa and Talwar, 1978; Choudhri et al., 1979; Kotian et al., 1981; Ramakrishnan, 1981) on NTM were published between 1961 and 1981 from India in which most NTM species were described as nonchromogens, M. intracellulare, Battey type photochromogens, and scotochromogens. A timeline of key landmarks from India in the field of NTM is provided in Figure 3. In 1981, C.N. Paramasivan, a medical microbiologist and pioneer in the field of NTM research in India, published the first comprehensive study from the Tuberculosis Research Centre, Madras (now known as National Institute for Research in Tuberculosis [NIRT], Chennai) (Paramasivan et al., 1985). Though occasional original articles/research articles on NTM had been published thereafter (Shanker et al., 1989; Karak et al., 1996), work on NTM in India peaked in the 2000s. Work reported from several national TB diagnostic laboratories (Figure 4) (Jain et al., 1991; Gupta et al., 2002; Narang et al., 2009; Parashar et al., 2009; Garima et al., 2012; Mishra et al., 2018; Sebastian et al., 2018a; Sebastian et al., 2018b; Sharma and Upadhyay, 2020; Shrivastava et al., 2020) contributed to spreading awareness about the various methods of NTM isolation and speciation among microbiologists in India. In that era, biochemical assays that were cumbersome and time-consuming had been used for speciation and sub-speciation. However, most of the data reported from these centers had emanated from laboratories, often based on a single time isolation, without establishing the clinical relevance of NTM isolates (Wolinsky, 1981; Narang et al., 2009; Mishra et al., 2018; Sharma and Upadhyay, 2020). No serious efforts were made to record and report their clinical details, or to trace these patients for their treatment. Though some studies (Jesudason and Gladstone, 2005; Maurya et al., 2015; Radha Bai Prabhu et al., 2019; Arvind et al., 2020; Gupta et al., 2020; Singh et al., 2020; Wani et al., 2020; Sali et al., 2021; Thangavelu et al., 2021; Das et al., 2022; Dadheech et al., 2023) were published subsequently, these have not systematically documented clinical and radiographic involvement in NTM patients.
History of nontuberculous mycobacteria: India. TRC, Madras, Tuberculosis Research Centre (now called National Institute for Research in Tuberculosis, Chennai); NTI, National Tuberculosis Institute; NITRD, National Institute of Tuberculosis and Respiratory Diseases; AIIMS, New Delhi, All India Institute of Medical Sciences; MGIMS, Mahatma Gandhi Institute of Medical Sciences, Wardha (Maharashtra); MALDI TOF MS, matrix assisted laser desorption time of flight mass spectrometry; ICMR, Indian Council of Medical Research.
Locations of major Indian laboratories and hospitals reporting NTM studies.
Researchers from the All India Institute of Medical Sciences (AIIMS), New Delhi (Singh et al., 2007; Khatter et al., 2008; Jain et al., 2014; Sharma et al., 2019; Wazahat et al., 2023) had contributed to the understanding of NTM disease from India. During the last decade, the research group led by Sharma (Sharma et al., 2019) was the first to publish data from India describing the “clinical relevance and treatment outcomes” of NTM isolated from patient specimens at AIIMS, New Delhi (Sharma et al., 2019). “Pseudo outbreaks” of health-care associated NTM-PD due to use of tap water to clean wounds, instruments is common; tap water is the major reservoir for MAC, M. kansasii, M. gordonae, M. xenopi, M. abscessus, M. fortuitum, M. chelonae, M. scrofulaceum, M. kansasii (Satta Y, et al., 2020; Sharma and Upadhyay, 2020). They had also advocated the role of molecular diagnosis of NTM using line probe assay and targeted gene sequencing (Sharma et al., 2019). The research group led by Camilla Rodrigues, Mumbai had also contributed to NTM research and worked on rapid diagnosis of NTM using matrix-assisted laser desorption/ionization-time of flight mass spectrophotometry (MALDI-TOF MS) (Shenai et al., 2009; Shenai et al., 2010; Jani et al., 2011; Rodrigues, 2015). There have also been occasional publications from the north-eastern states of India, namely, Arunachal Pradesh (Mudliar et al., 2022), Assam (Kalita et al., 2005), Meghalaya (Vise et al., 2016), Mizoram (De Mandal et al., 2015), and Tripura (Bhattacharya et al., 2021).
The first-of-its-kind, multicenter research project (Sharma et al., 2025) of national importance on NTM disease funded by the Indian Council of Medical Research (ICMR), Ministry of Health & Family Welfare, Government of India revealed that NTM were isolated from 226/71,143 (0.32%) TB suspects. In NTM-pulmonary disease (PD) (n=187), most common species isolated were M. avium complex (MAC) (27.8%), followed by M. kansasii (24.1%), M. abscessus (21.4%); in EP-NTM disease (n=39), M. abscessus (41%), M. fortuitum (41%), and MAC (15.4%) were the most common species isolated. This study is expected to provide a detailed account of prevalence, risk factors, clinical presentation, imaging features, laboratory diagnosis, and treatment outcomes of NTM-PD and EP-NTM (Sharma et al., 2025).
The first case of HIV in India was reported from Chennai in 1987 (Simoes et al., 1987). Subsequently, recognizing the importance of the deadly interaction between TB and HIV, the National (policy) Framework for Joint TB/HIV Collaborative Activities was jointly developed by the Revised National TB Control Programme (RNTCP) (now called National TB Elimination Programme [NTEP]) and the National AIDS Control Programme (NACP), bidirectional screening for HIV and TB was initiated (Department of AIDS Control et al., [[NoYear]]). In high TB burden countries including India, where TB is much more common, it is often difficult to differentiate pulmonary TB from the NTM-PD. The picture gets further complicated when sputum smear microscopy was extensively used for the diagnosis and treatment of TB in National TB Control Programmes, because of non-availability of mycobacterial culture facilities in high TB burden countries. These sputum smear-positive patients were often started on anti-TB drugs on the basis of sputum smear microscopy results. In the absence of mycobacterial culture, it is possible that several of these sputum smear-positive patients treated for TB would in fact have been patients with NTM-PD that were missed. The important differences between TB and NTM disease are shown in Supplementary Table 1 (Sharma and Upadhyay, 2020).
Immunopathogenesis of NTM disease
4
Immunopathogenesis of NTM disease is not completely understood because earlier literature lacked species and subspecies−level identification, surveillance, and environmental mapping, obscuring links between particular NTM, host factors, and characteristic immune phenotypes relative to tuberculosis (TB) (CruzAguilar et al., 2021; Lyu, 2024; Chancharoenthana et al., 2025). Modern liquid culture, probe−based and PCR diagnostics, and species−specific epidemiology revealed heterogeneity in virulence attributes such as biofilm formation, glycopeptidolipid (GPL) masking, and phagosome−level evasion, while cohort immunophenotyping uncovered variable Th1 tone, innate receptor signaling, and exhaustion signatures across clinical phenotypes (CruzAguilar et al., 2021; Lyu, 2024; Chancharoenthana et al., 2025). These advances explain why unified models are elusive and why species, morphotype, host structure, and immunogenetics must be integrated to describe NTM pathogenesis, in contrast to the more stereotyped, necrotizing Th1 pathology of human−adapted M. tuberculosis (Table 2) (Wu and Holland, 2015; Lyu, 2024; Chancharoenthana et al., 2025). Key immunopathogenic contrasts between nontuberculous mycobacteria and M. tuberculosis are summarized in Table 2.
Innate and adaptive immunity
Inhaled NTM adhere to mucus and compromised epithelium and are phagocytosed by alveolar macrophages that recognize PAMPs via PRRs, prominently TLR2, initiating IL−12–IFN−γ signaling, TNF production, and chemokine programs that recruit neutrophils, monocytes, NK cells, and lymphocytes to nascent lesions (Lyu, 2024; Chancharoenthana et al., 2025). In many patients with NTM−D, TLR2 expression and downstream IL−12 and TNF production are diminished in response to MAC stimulation, reflecting subtle innate deficits that bias toward persistence rather than sterilization at the macrophage–granuloma interface (CruzAguilar et al., 2021; Lyu, 2024; Chancharoenthana et al., 2025).
In contrast to TB’s frequent caseating necrosis, NTM granulomas show chronic inflammation and fibrosis with comparatively less caseation, consistent with heterogeneous Th1 drive, prolonged host–pathogen equilibrium in structurally abnormal lungs, and species−specific blockade of phagolysosomal killing (Table 3) (Lyu, 2024; Chancharoenthana et al., 2025). Adaptive control hinges on Th1 polarization driven by IL−12–IFN−γ that activates macrophage microbicidal programs and sustains organized granulomas, while Th17 pathways aid mucosal defense and neutrophil recruitment but can also amplify airway injury in bronchiectasis and cystic fibrosis when dysregulated (Lyu, 2024; Chancharoenthana et al., 2025). Disseminated NTM often unmasks inborn or acquired defects along the IL−12–IFN−γ axis, and even in pulmonary disease without overt inborn errors, signatures of T−cell exhaustion and altered cytokines point to broader immune dysregulation underlying chronic infection trajectories. Clinically, MAC typically impedes phagosome–lysosome fusion and neutralizes antimicrobial effectors to persist within vacuoles, whereas M. abscessus smooth variants use GPLs to dampen TLR2 sensing and rough (more virulent) variants damage phagosomes, induce type I IFN, and spread cell−to−cell, mapping distinct evasion modes to nodular−bronchiectatic versus rapidly destructive phenotypes.
Autophagy and xenophagy in NTM infection
4.1
Macroautophagy and xenophagy tag intracellular bacilli with ubiquitin and recruit cargo receptors (p62, OPTN, NBR1, NDP52) to LC3−decorated membranes for autophagosome formation and lysosomal degradation, yet NTM frequently impair fusion, acidification, or flux to persist in macrophages. In MAC, the dominant phenotype is vacuolar residence with blockade of phagosome–lysosome fusion and neutralization of ROS/RNS, whereas M. abscessus shows morphotype−dependent autophagy biology: smooth (GPL−rich) variants attenuate early PRR signaling while rough variants trigger abundant autophagosomes yet damage phagosomal membranes, induce apoptosis and type I IFN, and spread cell−to−cell. Additional diversity arises in M. marinum, where ubiquitinated bacilli can be routed to LAMP1−positive compartments via non−canonical routes and ESCRT−mediated vacuolar repair while ESX−1 upregulates autophagy genes but suppresses cargo degradation. These mechanistic bottlenecks create host−directed therapy opportunities: TFEB–lysosomal activators (amiodarone, rufomycins, resveratrol analog V46), AMPK activation (metformin), and trehalose–PIKFYVE–TFEB signaling restore degradative capacity and reduce intracellular MAC and M. abscessus in models, whereas macrolide−induced lysosomal alkalinization can impair autophagy−dependent killing of M. abscessus, emphasizing the need to align host−directed therapy with species and regimen context (Table 2).
In TB, xenophagy is also engaged but acidification and flux are curtailed by M. tuberculosis, and augmentation through vitamin D–CAMP, autophagy initiation, and TFEB activation restricts bacillary burden in experimental systems, highlighting convergences and divergences from NTM autophagy biology (Table 3).
Gut–lung axis and NTM disease
4.2
Human cohorts reveal reduced gut microbial diversity and taxa shifts in NTM−PD correlating with diminished TLR2−linked signaling, immune exhaustion markers, and early treatment responsiveness differences, while experimental restoration of commensals such as Prevotella copri augments TLR signaling and reduces pulmonary susceptibility.These data suggest that immunometabolic–microbiome interactions contribute to heterogeneous inflammatory phenotypes, reinfection risk, and outcome variability in NTM−PD beyond pathogen−intrinsic virulence and host structure alone.
Species mechanisms beyond MAC and M. abscessus complex
4.3
Beyond MAC and M. abscessus, M. kansasii in immunocompetent hosts often mimics TB with cavitation and stronger Th1−associated pathology, reflecting robust PRR signaling and intracellular persistence with more pronounced necrosis than MAC in some cohorts (Lyu, 2024; Chancharoenthana et al., 2025). M. xenopi favors vacuolar persistence with impaired phagolysosome fusion, producing indolent granulomatous inflammation and fibrosis in structurally abnormal lungs, and M. malmoense produces chronic fibrotic granulomas and low−grade inflammation analogous to MAC. Rapid growers such as M. fortuitum and M. chelonae form biofilms on airway surfaces and devices, provoking neutrophil−predominant inflammation and extracellular tolerance where source control is essential alongside antimicrobials (Lyu, 2024; Chancharoenthana et al., 2025). M. marinum causes cutaneous granulomatous disease via ESX−1–mediated phagosomal damage and ESCRT−linked routing, while autophagic cargo degradation is suppressed in vivo. M. ulcerans secretes mycolactone, driving local immunosuppression through mTOR activation and autophagy suppression with extensive necrosis, immunologically distinct from NTM−PD. M. haemophilum and pediatric lymphadenitis species such as M. scrofulaceum show cooler temperature preference and iron dependence with nodal tropism or strong localized granulomatous responses emphasizing regionally constrained Th1 control.
Extrapulmonary NTM disease pathogenesis
4.4
Extrapulmonary disease follows two routes: direct inoculation from procedures/trauma/injections causing skin/soft−tissue infection and lymphadenitis, or hematogenous spread in hosts with compromised cell−mediated immunity, with device−associated biofilms sustaining chronic infection and necessitating source control plus prolonged, species−directed therapy (Table 3). Disseminated disease is classically associated with IL−12–IFN−γ axis defects and anti–IFN−γ autoantibodies, while musculoskeletal and prosthetic infections reflect extracellular biofilms and neutrophilic inflammation distinct from caseating granulomas of extrapulmonary TB (Table 2).
Disseminated NTM disease
4.5
Disseminated NTM is strongly linked to inborn errors in the IL−12–IFN−γ pathway, including autosomal mutations in IL12B, IL12RB1, ISG15, IFNGR1, IFNGR2, STAT1, and IRF8, X−linked defects in IKBKG (NEMO) and CYBB (gp91phox), plus GATA2 deficiency and anti–IFN−γ neutralizing autoantibodies, spanning childhood to adult susceptibility across localized and disseminated syndromes.
Evolution of treatment
5
High TB burden countries have prioritized healthcare systems towards TB elimination because of the importance of person-to-person transmission in pulmonary TB, which forms ~80% of TB cases in immunocompetent adults, while NTM is not transmitted from person-to-person, except M. abscessus in cystic fibrosis cases, which is rare in high TB burden countries. Further, unlike in the natural history of TB, where TB infection (TBI, previously called latent TB infection) and TB disease have been described, latent NTM infection does not occur (Sharma and Upadhyay, 2020; Sharma and Upadhyay, 2021). The treatment of NTM-PD and EP-NTM varies according to the isolated species and, in some patients, subspecies, disease severity, underlying comorbidities. Unlike in TB, in vitro drug-susceptibility testing (DST), often does not predict in vitro clinical response and is, therefore, not considered essential for management of NTM diseases. DST should follow the Clinical and Laboratory Standards Institute (CLSI) guidelines (Clinical and Laboratory Standards Institute, 2018). Like TB, treatment of NTM disease involves administration of multiple drug combinations. Treatment regimens comprise various antimycobacterial drugs that need to be administered daily if the NTM-PD is severe for long periods of time. Most antimicrobial agents used for NTM diseases have been repurposed from TB or other bacterial infections. Over time, their roles have evolved based on emerging evidence, clinical experience, and drug resistance patterns, shaping the current therapeutic landscape (Table 3) (Maclean and Smith, 1947; Crow et al., 1957; Wolinsky et al., 1957; Lunn and Rees, 1964; Carruthers and Edwards, 1965; Jones, 1969; Rothstein, 1969; Becker et al., 1980; Dalovisio et al., 1981; Young et al., 1986; Snider et al., 1987; Woods and Washington, 1987; Bessesen et al., 1990; Young et al., 1991; Rastogi et al., 1996; Brown-Elliott et al., 2001; Griffith et al., 2003; Rolfe et al., 2007; Philley et al., 2015; Yuste et al., 2017; Griffith et al., 2018; Scott et al., 2018; Dedrick et al., 2019; Kim et al., 2020; Mingora et al., 2023). Because of the 200 NTM species/subspecies, it is not possible to carry out randomized controlled clinical trials. As a result of this, there are no evidence-based guidelines for the treatment of NTM diseases. In high TB burden countries like India, patients with NTM-PD have extensive unilateral or bilateral lung involvement due to past history of TB and, therefore, surgery is not possible. In EP-NTM, surgical debridement and multidrug treatment are possible (Sharma et al., 2021).
Since the emergence of the 1990 ATS guidelines (Anonymous, 1990), guideline-based treatment (GBT) often involved a multi-drug regimen to be continued for 12 months even after culture conversion, whereas watchful waiting is recommended in specific cases where clinical relevance of isolated NTM is not clear. In 2017, a phase 2 clinical trial (Olivier et al., 2017) had shown that addition of amikacin liposome inhalation suspension (ALIS) that delivers high concentrations to the site of infection, limiting systemic exposure was found to increase culture conversion rates in patients with refractory NTM lung disease compared to GBT alone. Following encouraging results noted in this trial (Mingora et al., 2023) and another phase 3 trial published in 2018 (Griffith et al., 2018), addition of ALIS to GBT in patients who fail to convert after 6 months of treatment has been included in the ATS/ERS/ESCMID/IDSA clinical practice guideline (Daley et al., 2020).
It became known that organism is naturally resistant to pyrazinamide (a prodrug) due to reduced pyrazinamidase activity, preventing conversion of the drug into pyrazinoic acid (active bactericidal compound) (Sun and Zhang, 1999), pyrazinamide is not recommended for M. kansasii pulmonary disease. The total duration of treatment of M. kansasii and M. szulgai disease is 12 months, inclusive of culture conversion time period. In all other NTM species, treatment duration is for 12 months after culture conversion. The importance of an active erythromycin ribosomal methylase [erm] 41 gene in rapidly growing mycobacteria like M. abscessus in inducible resistance to macrolides was recognized in 1996 (Meier et al., 1996) and this has led to checking for erm (Wolinsky, 1979) gene status in M. abscessus in the later ensuing guidelines (Wallace et al., 1996).
Consequently, NTM-PD is associated with a huge socioeconomic burden on the patient. As a result, multidisciplinary teams are believed to be most suited for helping patients, providing adjunctive therapy in addition to GBT as needed. However, studies have indicated that in general compliance with GBT during the therapy of NTM-PD is very low. Surgery constitutes a first-line treatment option in NTM-PD if the patient has localised disease, good nutritional and cardiopulmonary functional status. The treatment of EP-NTM involves combining antimycobacterial therapy with possible surgery to achieve the best outcome. Adjunctive surgery is described as a possibility for selected individuals who have failed antibiotic treatment due to drug-resistant NTM isolates and drug-induced toxicity. Similar to the diagnosis, there are currently no published guidelines for the treatment of EP-NTM. Recently dated June 14, 2024, the NTEP, in the website “Knowledge Base for the National TB Elimination Program – NTEP”, under “Treatment regimen for Non-Mycobacterium tuberculosis (NTM) (ID 1791)” (https://ntep.in/node/1792/CP-treatment-regimen-ntm) provided “suggested treatment regimen covering maximum Non-Mycobacterium Tuberculosis (NTM) mainly MAC”. Overall, a lack of awareness among healthcare workers, inadequate laboratory infrastructure, and non-existent programmatic management constitutes obstacles in ensuring rational scientific diagnosis and treatment of NTM disease.
Biomarkers associated with NTM disease risk, treatment response, and prognosis
6
Culture-independent markers may be biological components of NTM or the host immune response to the organism (Table 4) (Schaefer, 1965; Szulga et al., 1966; Birn et al., 1967; Marks et al., 1969; Jenkins et al., 1971; Yoder and Schaefer, 1971; Jenkins and Marks, 1973; Brennan et al., 1978; Torrelles et al., 2002; Schorey and Sweet, 2008; Watanabe et al., 2011; Kitada et al., 2017; Han et al., 2020; Matsuda et al., 2020; Nithichanon et al., 2020; Steffen et al., 2020; Kawasaki et al., 2022; Mani-Varnosfaderani et al., 2022; Manbenmad et al., 2023; Park et al., 2023; Anh et al., 2024; Choi et al., 2024; Jia et al., 2024). To present, the most promising diagnostic and disease monitoring biomarkers include serum anti-glycopeptidolipid (GPL)-core antibodies [immunoglobulin A (IgA) and immunoglobulin G (IgG)]. Schaefer, Marks, and Jenkins’ work in the 1960s and 1970s laid the groundwork for identifying and characterizing GPLs (Rothstein, 1969) Later, it was demonstrated that thin-layer chromatography could be used to describe various membrane lipids of NTM and supplement the use of seroagglutination in NTM categorization (Schaefer, 1965; Szulga et al., 1966; Birn et al., 1967; Marks et al., 1969; Jenkins et al., 1971; Yoder and Schaefer, 1971; Jenkins and Marks, 1973). Patrick Brennan, a renowned microbiologist from the USA, and his colleagues discovered the mycobacterial glycopeptidolipid (GPL) structure in 1978–79 and continued to study its various aspects in different NTM species until early 2000 (Brennan et al., 1978; Torrelles et al., 2002). The GPLs are a class of glycolipids present on the outermost surfaces of several NTM, including MAC, M. smegmatis, M. abscessus, and M. fortuitum. GPL core structure consists of a 3-hydroxy or 3-methoxy C26-C33 fatty acyl chain N connected to a tripeptide-amino-alcohol composed of D-phenylalanine-D-allo-threonine-D-alanine-L-alaninol (Schorey and Sweet, 2008). The lipopeptide core is glycosylated with 6-deoxytalose (linked to allo-threonine) and α-L-rhamnose (linked to L-alaninol) (Schorey and Sweet, 2008). The resultant oligosaccharide residues are methylated to generate non-serovar-specific (apolar) GPLs (nsGPLs), which are present in all GPL-producing mycobacteria (Schorey and Sweet, 2008). However, MAC species also synthesize polar GPLs, in which some oligosaccharides residues are attached to the 6-deoxytalose-producing serovar-specific GPLs (ssGPLs) (Schorey and Sweet, 2008). Some RGM (M. smegmatis, M. abscessus, M. fortuitum, and M. chelonae) species also produce polar GPLs different from MAC due to the presence of 3,4-di-O-methyl rhamnose attached to the alaninol-linked 3,4-di-O-methyl rhamnose. By, contrast, these GPLs are not present in Mtb or M. kansasii (Schorey and Sweet, 2008). The levels of serum anti-GPL-core IgA estimated by enzyme-linked immunosorbent assay (ELISA) have been used for the diagnosis of MAC-PD in clinical practice in Japan since 2011 (Watanabe et al., 2011). Since then, a handful of studies from different countries have been reported on anti-GPL-core IgA-based MAC-PD diagnosis. However, variable sensitivity (48%-85%) and specificity (52% -99%) of anti-core GPL-IgA-based MAC-PD diagnosis were reported in different studies (Kitada et al., 2017; Matsuda et al., 2020; Kawasaki et al., 2022; Choi et al., 2024). It has been noted in a few studies that using a lower diagnostic cut-off value than the manufacturer’s cut-off of anti-core GPL-IgA ELISA kits increases sensitivity and specificity to diagnose MAC-PD (Watanabe et al., 2011; Choi et al., 2024). Overall, varying sensitivities and specificities may be attributable to different diagnostic cut-offs for anti-core GPL-IgA levels, as well as differences in geographic, ethnic, and research population characteristics (Choi et al., 2024). Moreover, factors associated with lower anti-core GPL-IgA levels include immunocompromised states such as HIV/AIDS (patients on prophylactic macrolide monotherapy), cancer, etc. resulting in the possibility of higher false-negative rates. Therefore, different optimal cut-off points of anti-core GPL-IgA levels must be set in these populations (Choi et al., 2024).
Some studies also reported that anti-core GPL-IgA levels are useful in detecting M. abscessus infection (Manbenmad et al., 2023). It was also observed that the IgA2 subclass of IgA antibodies is predominantly elevated in patients with NTM disease (Steffen et al., 2020). Few studies from Thailand also showed the utility of anti-core GPL-IgG which has a greater specificity than anti-core GPL-IgA in diagnosing NTM-PD. Further, it has been demonstrated that plasma anti-GPL-core IgG levels in patients with SGM were higher than in patients with RGM. Patients with disseminated NTM also exhibit higher elevated anti-core GPL-IgA (Nithichanon et al., 2020).
Recently, high throughput gene sequencing revealed some genetic markers that can differentiate between NTM, TB infection (TBI), and active TB (Schorey and Sweet, 2008). The differential expressions of four hub genes: hemoglobin-α 1 and 2 (HBA1 and HBA2), hemoglobin subunit delta (HBD), and methyltransferase-like 7B (METTL7B) have been shown useful in identifying NTM, LTBI, and active TB patients (Jia et al., 2024). Upregulated HBA1 and 2 in LTBI patients distinguish individuals having NTM disease. Upregulated HBD genes in active TB patients can distinguish them from NTM patients. Moreover, the downregulated METTL7B gene has been associated with LTBI and may distinguish LTBI from active TB (Jia et al., 2024). A South Korean study demonstrated that homo sapiens microRNAs (has-miR) such as hsa-miR-484, hsa-miR-584-5p, hsa-miR-625-3p, and hsa-miR-4732-5p are differentially expressed in individuals with NTM-PD and may serve as possible biomarkers for the disease (Han et al., 2020). A Canadian study on cystic fibrosis discovered that breath samples from patients with NTM-PD had differential levels of volatile molecules. When compared to healthy controls, NTM-PD patients had significantly higher levels of these compounds, including 2,3,5,8-tetramethyl-decane, 2,2,4,6,6-pentamethyl-heptane, 2,3-dimethyl-3-heptene, and tridecane, but ethanol levels were significantly lower (Mani-Varnosfaderani et al., 2022). A South Korean study revealed urine biomarkers that can differentiate between NTM and TB disease (Anh et al., 2024). These biomarkers included methionine, hypoxanthine, and acetylserotonin, which were all 1.8-fold elevated in NTM patients (Anh et al., 2024). In contrast, the level of 4-guanidinobutanoate decreased nearly threefold, while adenosine, cystine, and acetylcholine decreased approximately 1.7fold in NTM patients (Anh et al., 2024).
There are very few studies on prognostic markers for NTM disease. A South Korean study (Park et al., 2023) showed that elevated serum C-reactive protein (CRP) levels at diagnosis predicted a worse prognosis in individuals with non-tuberculous mycobacterial pulmonary disease (NTM-PD). Approximately one-fourth of NTM-PD patients had elevated CRP levels, which increased their risk of death (Park et al., 2023). Data from Korea (Koh, et al., 2012) suggest that M. intracellulare are more virulent and are associated with a worse prognosis. NTM disease with M. avium and M. intracellulare behave similarly clinically and therapeutic response wise. M. avium often presents with disseminated disease (Koh, et al., 2012).
Conclusion
7
NTM has undertaken a rather very long journey to reach its current status, from anonymity to intense research efforts are on to unravel several areas, such as rapid diagnosis, independent of culture, development of new biomarkers for early diagnosis, and monitoring treatment response. Equal importance is being given to drug discovery for the development of new drugs for an effective NTM treatment. In spite of a heightened degree of awareness about NTM diseases among microbiologists and clinicians in India more efforts are required to determine the exact species/subspecies of isolated NTM and other high TB burden countries disease prevalence including geographical variations and treatment outcomes. The measures should be taken by various stakeholders to formulate India-centric guidelines for diagnosis and treatment of NTM disease and eventually inclusion in the national healthcare programmes of India. Efforts should be made to establish a National Registry of NTM in India.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alvarez G. Tavel R. (1885). Recherches sur le bacille de Lustgarten. Arch. Physiol. Norm Pathol. 3, 303–306.
- 2Anh N. K. Phat N. K. Thu N. Q. Tien N. T.N. Eunsu C. Kim H. S. . (2024). Discovery of urinary biosignatures for tuberculosis and nontuberculous mycobacteria classification using metabolomics and machine learning. Sci. Rep. 14, 15312. doi: 10.1038/s 41598-024-66113-x, PMID: 38961191 PMC 11222504 · doi ↗ · pubmed ↗
- 3Anonymous (1990). Diagnosis and treatment of disease caused by nontuberculous mycobacteria. Am. Rev. Respir. Dis. 142, 940–953. doi: 10.1164/ajrccm/142.4.940, PMID: 2282111 · doi ↗ · pubmed ↗
- 4Anonymous (1997). Diagnosis and treatment of disease caused by nontuberculous mycobacteria. This official statement of the American Thoracic Society was approved by the Board of Directors, March 1997. Medical Section of the American Lung Association. Am. J. Respir. Crit. Care Med. 156, S 1–25. doi: 10.1164/ajrccm.156.2.atsstatement, PMID: 9279284 · doi ↗ · pubmed ↗
- 5Aronson J. D. (1926). Spontaneous tuberculosis in salt water fish. J. Infect. Dis. 39, 315–320. doi: 10.1093/infdis/39.4.315 · doi ↗
- 6Arvind B. Medigeshi G. R. Kapil A. Xess I. Singh U. Lodha R. . (2020). Aetiological agents for pulmonary exacerbations in children with cystic fibrosis: An observational study from a tertiary care centre in northern India. Indian J. Med. Res. 151, 65–70. doi: 10.4103/ijmr.IJMR_1275_18, PMID: 32134016 PMC 7055172 · doi ↗ · pubmed ↗
- 7Becker G. J. Walker R. G. Dziukas L. J. Harvey K. J. Valentine R. Kincaid-Smith P . (1980). Renal infection with Mycobacterium chelonei. Aust. N Z J. Med. 10, 44–47. doi: 10.1111/j.1445-5994.1980.tb 03417.x, PMID: 6929675 · doi ↗ · pubmed ↗
- 8Bessesen M. T. Berry C. D. Iohnson M. A. Klaus B. Blaser M. J. Ellison R. T . (1990). “ Site of origin of disseminated MAC infectionin AIDS [abstract 1268],” in Program and abstracts of the 30th interscience conference on antimicrobial agents and chemotherapy (Atlanta) ( American Society for Microbiology, Washington, DC).