Cytochrome P450-catalyzed allylic oxidation of pentalenene to 1-deoxypentalenic acid in pentalenolactone biosynthesis
Jing Li, Chengde Zhang, Shiwen Wu, Jiao Xue, Ke Chen, Zixin Deng, Dongqing Zhu

TL;DR
Scientists discovered how a specific antibiotic compound is formed using a P450 enzyme and redox proteins in a bacterial species.
Contribution
The study identifies the P450 enzyme and redox proteins responsible for a missing step in pentalenolactone biosynthesis.
Findings
Cytochrome P450 pentalenene oxygenase catalyzes the oxidation of pentalenene to 1-deoxypentalenic acid.
Ferredoxin XNR_5179 and ferredoxin reductase XNR_4478 support the activity of pentalenene oxygenase.
The identified pathway fills a gap in the biosynthesis of pentalenolactone.
Abstract
Pentalenolactone is a sesquiterpene antibiotic from Streptomyces. Its biosynthetic pathway has been elucidated, except for the oxidation of pentalen-13-al to 1-deoxypentalenic acid. In this study, we show that cytochrome P450 pentalenene oxygenase catalyzed the formation of 1-deoxypentalenic acid. Ferredoxin XNR_5179 and ferredoxin reductase XNR_4478 from S. albus are suitable redox proteins for pentalenene oxygenase. The biosynthetic pathway presented fills a gap in the biosynthetic pathway of pentalenolactone and provides an example of cytochrome P450 enzyme activity being affected by redox proteins. Image, graphical abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
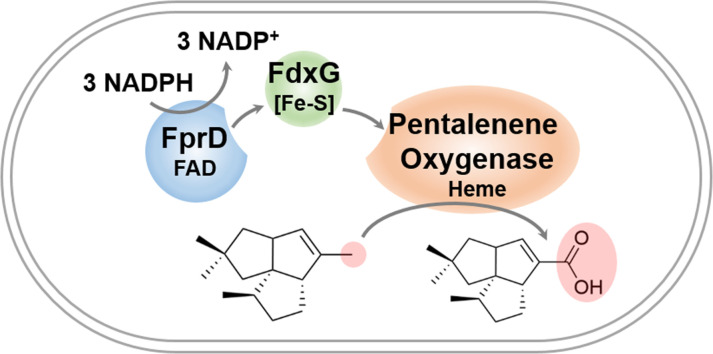 Figure 1
Figure 1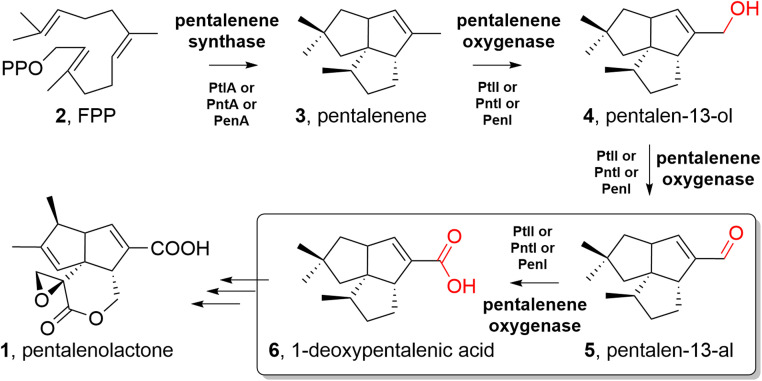 Figure 2
Figure 2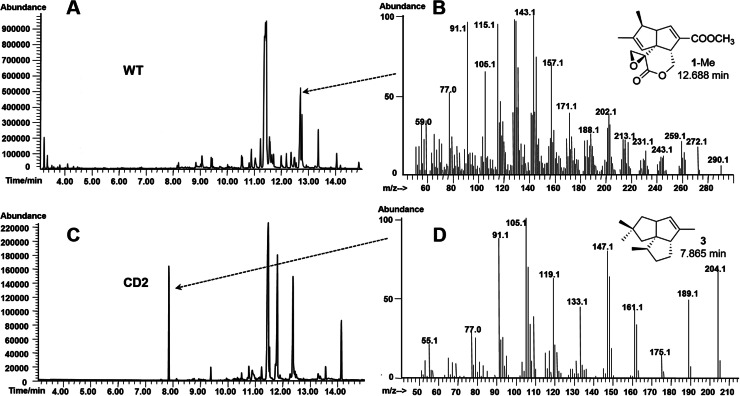 Figure 3
Figure 3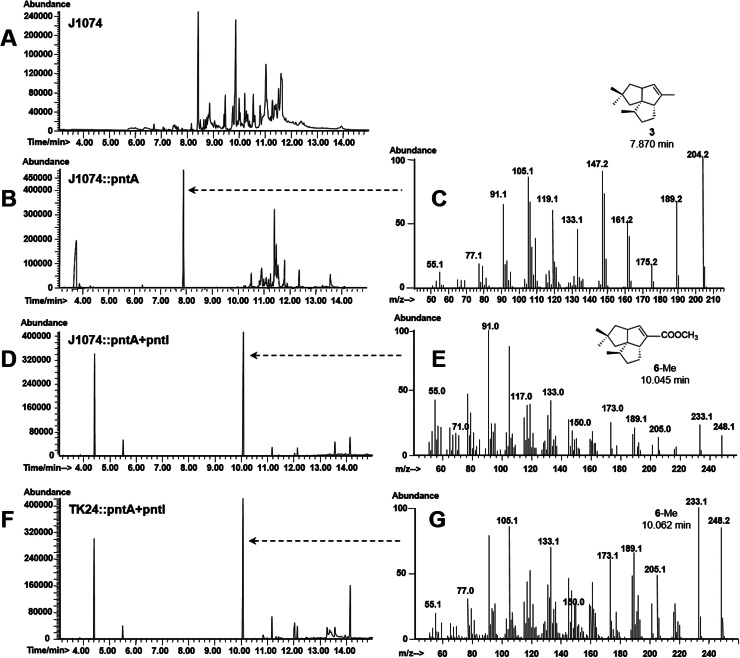 Figure 4
Figure 4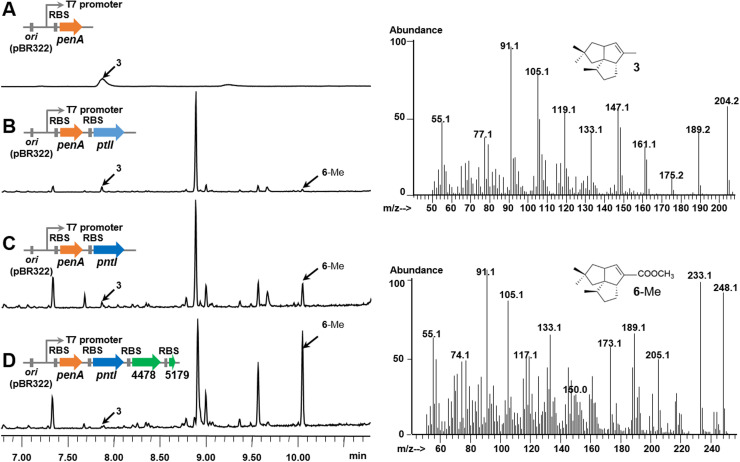 Figure 5
Figure 5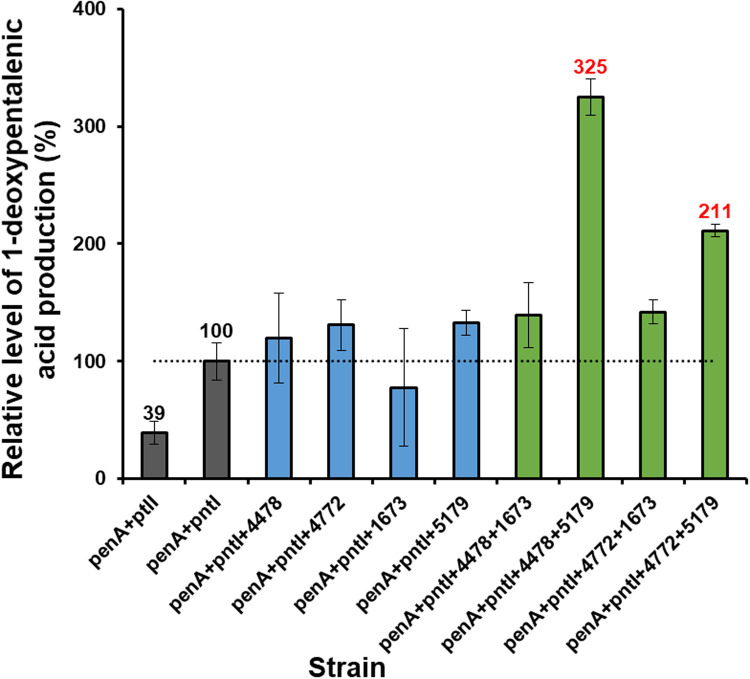 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant biochemistry and biosynthesis · Microbial Natural Products and Biosynthesis · Metal-Catalyzed Oxygenation Mechanisms
Cytochrome P450s are a family of heme-containing monooxygenases that catalyze various oxidative reactions [1]. Decades after their discovery, P450 systems have been recognized as powerful biocatalysts in synthetic biology [2,3]. Pentalenolactone (1) is a sesquiterpene derived from farnesyl diphosphate (FPP, 2) (Fig. 1) [[4], [5], [6], [7]]. Pentalenene oxygenase, encoded by the cytochrome P450 gene ptlI from the pentalenolactone (1) biosynthetic gene cluster of Streptomyces avermitilis [8], catalyzes the conversion of pentalenene (3) to pentalen-13-al (5) via stepwise oxidation of pentalen-13-ol (4) [9]. An open question remains as to how aldehyde pentalen-13-al (5) is converted into 1-deoxypentalenic acid (6). It is conceivable that another enzyme within the biosynthetic gene cluster is responsible for this conversion. Subsequently, the pentalenolactone biosynthetic pathway was identified in S. avermitilis, S. exfoliatus and S. arenae [[10], [11], [12], [13], [14], [15], [16], [17], [18], [19], [20]], and no genes involved in the biosynthesis of 6 were found in these gene clusters.Fig. 1. Biosynthesis of pentalenolactone (1).Fig 1
Cytochrome P450s catalyze numerous oxygenation reactions including the three-step oxidation of a methyl group to a carboxylic acid. FscP in FR-008/candicidin biosynthesis [21], AmphN in amphotericin biosynthesis [22], NysN in nystatin biosynthesis [23], ScnG/PimG in pimaricin biosynthesis [24], and RosC in 20-carboxyrosamicin biosynthesis [25] are all type examples from bacteria. This type of three-step oxidation catalyzed by P450 also occurs during the biosynthesis of natural plant products, such as celastrol [26], ent‑kaurenoic acid [27,28], bile acid [29], artemisinic acid [30,31], and carlactonoic acid [32]. Therefore, we speculated that pentalenene oxygenase PtlI in S. avermitilis and its homologs PenI in S. exfoliatus and PntI in S. arenae might be responsible for the allylic oxidation of 3 to 6 under appropriate conditions (Fig. 1). In this study, we focused on the pentalenene oxygenase genes pntI from S. arenae and penI from S. exfoliatus and proved the above hypothesis using heterologous expression in Streptomyces and Escherichia coli. We also identified suitable electron transport proteins for pentalenene oxygenase that promote the catalytic efficiency of this reaction.
First, we deleted the pentalenene oxygenase gene penI in the pentalenolactone producer S. exfoliatus UC5319 to generate a ΔpenI mutant CD2, in which an internal 1194-bp fragment (from 100 nt to 1293 nt) in penI was replaced by a 6-bp NdeI site (Fig. S1 A; all bacterial strains and plasmids used in this work were listed in Tables S1 and S2). PCR amplification with the primer pair UCF and UCR yielded a 601-bp product from the double-crossover strain S. exfoliatus CD2 and a 1795-bp product from the wild-type strain S. exfoliatus UC5319 (Fig. S1 B), all of which are listed in Table S3. Wild-type S. exfoliatus UC5319 and the penI mutant CD2 were then cultured, acidified, and extracted. The organic extracts were dried, concentrated, methylated, and analyzed using gas chromatography-mass spectrometry (GC–MS). The GC–MS results showed that the wild-type strain UC5319 produced 1 (Fig. 2A and B). The penI mutant CD2 did not produce any detectable 1 or accumulated 3 (Fig. 2C and D), which is the cyclization product of FPP (2) catalyzed by pentalenene synthase and was used as the substrate for the reaction catalyzed by recombinant PtlI in vitro [9]. We also deleted the pntI gene in another pentalenolactone producer, S. arenae TU469, to generate a ΔpntI mutant. GC–MS analysis confirmed that the pntI mutant generated accumulated pentalenene (3) and did not produce 1, unlike the wild-type strain TU469 (not shown).Fig. 2GC–MS analysis of the S. exfoliatus penI mutant CD2. A, Gas chromatography (GC) analysis of wild-type S. exfoliatus UC5319. B, Mass spectra (MS) of 1-Me produced by the S. exfoliatus UC5319. C, GC analysis of the S. exfoliatus penI mutant CD2. D, MS of 3 produced by the S. exfoliatus penI mutant CD2.Fig 2
Next, two Streptomyces strains, S. albus and S. lividans, which do not contain the pentalenolactone biosynthetic gene cluster, were selected as hosts for the heterogeneous expression of the pentalenene oxygenase gene. The DNA fragment carrying the pentalenene synthase gene pntA and the pentalenene oxygenase gene pntI was amplified from S. arenae TU469 and inserted into the integrative vector pIB139 carrying the constitutively strong promoter ermEp* to generate the plasmid pWHU1702. Plasmid pWHU1703, which contains pntA, was used as a control. Plasmids pWHU1702 and pWHU1703 were then transformed into S. albus J1074 and S. lividans TK24, respectively. The resulting strains were cultured, acidified, and extracted. The organic extracts were dried, concentrated, methylated, and analyzed via GC–MS. GC-MS analysis of S. albus revealed that the wild-type strain J1074 did not produce any intermediates of 1 (Fig. 3A) and that J1074::pWHU1703 produced 3 (Fig. 3B and C), the product of FPP catalyzed by PntA. Further, 1-deoxypentalenic acid methyl ester (6-Me) was detected in the culture of J1074::pWHU1702 (Fig. 3D and E). GC–MS analysis of the cultures of S. lividans gave the same results as S. albus: 6-Me was found in the culture of TK24::pWHU1702 (Fig. 3F and G) but not in the cultures of the control strains TK24 and TK24::pWHU1703 (not shown). Next, 4 and 5 were not detected in the cultures of J1074::pWHU1702 and TK24::pWHU1702, revealing that PntI catalyzed 3 to 6 efficiently in Streptomyces hosts. In summary, these results indicate that pentalenene oxygenase catalyzes the allylic oxidation of 3 to 6.Fig. 3. Gas chromatography-mass spectrometry (GC–MS) analysis of S. albus and S. lividans strains harboring the pentalenene synthase gene pntA and the pentalenene oxygenase gene pntI from S. arenae. A, GC analysis of wild-type S. albus J1074; B, GC analysis of the control strain S. albus J1074::pWHU1703; C, MS spectra of 3 produced by S. albus J1074::pWHU1703; D, GC analysis of S. albus J1074::pWHU1702; E, MS spectra of 6-Me produced by S. albus J1074::pWHU1702; F, GC analysis of S. lividans TK24::pWHU1702; G, MS spectra of 6-Me produced by S. lividans TK24::pWHU1702.Fig 3
In a study by Quaderer et al., four pairs of electron transport chain proteins were tested in reactions catalyzed by recombinant PtlI [9]: (1) typical redox pairs from spinach, (2) putidaredoxin and putidaredoxin reductase from Pseudomonas putida, (3) flavodoxin (Fld, Sequence ID: WP_001018618) and flavodoxin reductase (Fdr, Sequence ID: WP_000796332.1) from E. coli, (4) and ferredoxin FdxD (SAVERM_3129) and ferredoxin reductase FprD (SAVERM_5675) from S. avermtilis. Oxidation reactions using the first or second redox pairs were unsuccessful. However, 4 and 5 were detected during incubation with the third or fourth redox pair. Therefore, 6 was not formed, probably due to mismatched redox pairs in the reactions.
The genome of the pentalenolactone producer S. avermitilis contains nine ferredoxin (Fdx) genes and six ferredoxin reductase (FDR) genes, whereas the genome of S. albus, which was successfully heterologously expressed, contains only four Fdx genes and three FDR genes. To reduce this workload, we screened suitable electron transport chain proteins from S. albus. The protein sequences of Fdxs and FDRs from S. avermitilis MA-4680 (http://avermitilis.ls.kitasato-u.ac.jp/) were used to search protein databases on S. albus (GenBank: CP004370.1), S. lividans (GenBank: CP009124.1), and several Streptomyces spp. containing the biosynthetic gene cluster for pentalenolactone (1), including S. bingchenggensis BCW-1 (GenBank: CP002047.1), S. collinus TU365 (GenBank: CP006259.1), S. leeuwenhoekii C34 (NCBI Reference Sequence: NZ_LN831790.1), S. cyaneogriseus NMWT 1 (NCBI Reference Sequence: NZ_CP010849.1), S. milbemycinicus SIPI-054 (NCBI Reference Sequence: NZ_CP109672.1), and S. genisteinicus CRPJ-33 (NCBI Reference Sequence: NZ_CP060825.1). The results are summarized in Table S4. In the nine Fdx proteins identified, the identity (≥93 %) and similarity (≥96 %) of SAVERM_3129 (FdxD) to its homologs were the highest, followed by SAVERM_6676 (FdxG; identity, ≥81 %; similarity, ≥85 %). In the six predicted FDR proteins (including a predicted ferredoxin-nitrite reductase), SAVERM_5675 (FprD) had the highest levels of identity (≥80 %) and similarity (≥86 %) to its homologs, followed by SAVERM_6097 (FprE; identity, ≥72 %; similarity, ≥80 %). The best matches between two Fdx proteins (SAVERM_3129 and SAVERM_6676) and two FDR proteins (SAVERM_5675 and SAVERM_6097) in S. albus were XNR_1673, XNR_5179, XNR_4478, and XNR_4722, which were selected as candidates for further studies.
To avoid interference from the endogenous redox proteins of Streptomyces strains, E. coli BL21(DE3) (pMH1, pFZ81) [33] was chosen as the host. The DNA fragment carrying the pentalenene synthase gene penA was inserted downstream of the T7 promoter of pET21a to generate plasmid pLJ5 (Fig. 4A). Two DNA fragments carrying the pentalenene oxygenase genes ptlI and pntI were then inserted downstream of penA in pLJ5 to generate the plasmids pLJ80 and pLJ9, respectively (Fig. 4B and C). The three plasmids were transferred into E. coli BL21(DE3) (pMH1, pFZ81) respectively, followed by fermentation, isolation, and GC–MS analysis. As expected, the heterologous expression strain of penA produced 3 (Fig. 4A). Unexpectedly, 4 and 5 were not detected in the fermentation products of the heterologous expression strains of penA and ptlI (or pntI); however, 6 was detected (Fig. 4B and C). This result differed from those of the in vitro biochemical reactions of the recombinant protein PtlI [9]. Based on our previous research, the catalytic activity of an enzyme in a cell is often higher than its activity in an in vitro biochemical reaction in the form of a recombinant protein, which may explain why PtlI can catalyze a three-step oxidation in the cell, whereas it can only perform two-step oxidation in vitro. Our results confirmed that 6 is the product of the PtlI-catalyzed reaction and that the redox pairs in E. coli are functional in the reaction.Fig. 4GC–MS analysis of E. coli BL21(DE3) strains harboring the pentalenene synthase gene penA from S. exfoliatus and pentalenene oxygenase gene pntI form S. arenae. A, BL21(DE3) (pMH1, pFZ81, pLJ5); B, BL21(DE3) (pMH1, pFZ81, pLJ80); C, BL21(DE3) (pMH1, pFZ81, pLJ9); D, BL21(DE3) (pMH1, pFZ81, pLJ62).Fig 4
It is also worth noting that the catalytic activity of PntI (0.21 mg/L) was significantly higher than that of PtlI (0.08 mg/L), increasing by about 2.6-fold (Figs. 4B and C and 5 [gray columns]). This phenomenon may be caused by differences in enzyme activity due to differences in key amino acids between PntI and PtlI. Both pntI and ptlI were inserted into a high-copy number vector and were under the control of the strong IPTG-induced promoter T7 and the same RBS so that the transcription levels of the two genes were comparable. However, subtle differences in gene and protein sequences can lead to marked differences in the expression levels and solubility of the final protein, which may be another reason for the differences in their catalytic efficiencies. The catalytic efficiency of Class I P450 enzymes is influenced by the P450: Fdx: FDR ratio. PtlI and PntI were overexpressed in E. coli hosts, but the endogenous E. coli proteins Fdx and FDR were underexpressed, which may account for the relatively low catalytic efficiency.Fig. 5. Relative level of 1-deoxypentalenic acid production of E. coli BL21(DE3) strains harboring the pentalenene synthase gene penA from S. exfoliatus, the pentalenene oxygenase gene pntI from S. arenae, and electron transport protein genes from S. albus. The relative levels of 1-deocypentalenic acid produced were investigated in the following strains: penA+ptlI, BL21(DE3) (pMH1, pFZ81, pLJ80); penA+pntI, BL21(DE3) (pMH1, pFZ81, pLJ9); penA+pntI+4478, BL21(DE3) (pMH1, pFZ81, pLJ10); penA+pntI+4772, BL21(DE3) (pMH1, pFZ81, pLJ55); penA+pntI+1673, BL21(DE3) (pMH1, pFZ81, pLJ56); penA+pntI+5179, BL21(DE3) (pMH1, pFZ81, pLJ57); penA+pntI+4478+1673, BL21(DE3) (pMH1, pFZ81, pLJ61); penA+pntI+4478+5179, BL21(DE3) (pMH1, pFZ81, pLJ62); penA+pntI+4772+1673, BL21(DE3) (pMH1, pFZ81, pLJ63); and penA+pntI+4772+5179, BL21(DE3) (pMH1, pFZ81, pLJ64). Error bars indicate the standard deviation (n = 3).Fig 5
BL21(DE3) (pMH1, pFZ81, and pLJ9), a strain containing penA and pntI, was used as the starting strain in the next work. Four Fdx and FDR genes from S. albus were inserted downstream of pntI in pLJ9 to generate the plasmids pLJ10 (carrying XNR_4478), pLJ55 (carrying XNR_4772), pLJ56 (carrying XNR_1673), and pLJ57 (carrying XNR_5179), which were then transformed into BL21(DE3) (pMH1, pFZ81). Compared to the control strain BL21(DE3) (pMH1, pFZ81, and pLJ9), no significant change in the yield of 6 was observed in the four transformants (Fig. 5, blue columns).
The Fdx and FDR genes were then combined, and four plasmids were constructed: pLJ61 (containing XNR_4478 and XNR_1673), pLJ62 (containing XNR_4478 and XNR_5179), pLJ63 (containing XNR_4772 and XNR_1673), and pLJ64 (containing XNR_4772 and XNR_5179). Compared to the control strain BL21(DE3) (pMH1, pFZ81, pLJ9), the yield of 6 did not change in the two strains harboring Fdx genes XNR_1673, BL21(DE3) (pMH1, pFZ81, pLJ61), and BL21(DE3) (pMH1, pFZ81, pLJ63). In contrast, the yield of 6 increased 2-fold (0.68 mg/L) and 1-fold (0.44 mg/L) in the two strains carrying the Fdx gene XNR_5179, including BL21(DE3) (pMH1, pFZ81, and pLJ62), and BL21(DE3) (pMH1, pFZ81, and pLJ64). (Figs. 4D and 5 [green columns]). These results showed that XNR_5179 and XNR_4478 are the most suitable electron transport proteins for PntI and suggested that Fdxs have a greater effect on the catalytic activity of P450 proteins than FDRs.
To obtain purified proteins for in vitro biochemical reactions, we constructed protein expression plasmids for the genes pntI, XNR_5179, and XNR_4478, and expressed the recombinant proteins using an E. coli protein expression system; however, no soluble proteins were observed. Despite the lack of data on in vitro biochemical reactions, we believe that XNR_5179 and XNR_4478 efficiently assist PntI in catalyzing the formation of 6. Based on the high degree of identity and similarity of their protein amino acid sequences (Table S4), we believe that FdxG (SAVERM_6676) and FprD (SAVERM_5671) in S. avermitilis are appropriate natural redox proteins of the pentalenene oxygenase PtlI. In a study by Quaderer [9], 6 was not detected in the pentalenene oxygenase PtlI assay using electron transport proteins from S. avermitilis because of the incorrect use of ferredoxins (SAVERM_3129).
The P450 family is diverse and is used to catalyze key steps in drugs such as steroids and antibiotics. In synthetic biology, the combination of P450 with artificial or heterologous redox pairs can extend their substrate range and catalytic capacity. The catalytic process of P450 involves complex electron transfer steps, and its catalytic activity is highly dependent on the efficiency of the electron transfer chain. Studying the fitness of the electron transport chain can help elucidate how electrons are transferred from NAD(P)H to the P450 active site. By optimizing the interactions of relevant proteins in the electron transport chain with P450, the efficiency of electron transfer can be improved, thereby enhancing the catalytic capacity of the enzyme and providing a theoretical basis for protein and metabolic engineering.
Class I cytochrome P450 proteins in bacteria do not fuse with their electron transport chain proteins to form a single protein. In most cases, their genes are not clustered on the chromosome, making the identification of the natural electron transport chain proteins of cytochrome P450 proteins very difficult [[34], [35], [36]]. Early studies on prokaryotic cytochrome P450s used heterologous electron transport chain proteins. However, an increasing number of studies have shown that electron transport chain proteins affect not only enzyme activity but also the chemo-, regio-, and stereoselectivity of the catalytic reaction [37,38]. Therefore, it is necessary to identify the natural intracellular electron transport chain proteins of cytochrome P450 to uncover the true biosynthetic pathways of natural products in a cell.
FR-008/Candicidin, amphotericin, nystatin, and pimaricin are polyene macrolide antibiotics with similar structures. Therefore, FscP, AmphN, NysN, and ScnG/PimG are homologous proteins with a high degree of sequence identity. To investigate the structure-activity relationship of various drugs, the corresponding P450 genes were knocked out, and decarboxylated compounds were derived from the resulting mutant strains to assess alterations in their activity. However, this study did not extend the examination of electron transport chain proteins associated with these P450s. Rosamicin is a macrolide antibiotic, and its carboxylation is achieved by overexpressing the P450 gene rosC and electron transport chain protein genes from P. putida in an E. coli host. However, researchers have not focused on the natural electron transport chain proteins of RosC. Based on the available experimental data, we hypothesized that the concurrent overexpression of E. coli electron transport chain proteins in strain BL21(DE3) (pMH1, pFZ81, and pLJ9) (Fig. 4C) could increase the enzymatic activity of P450 pentalenene oxygenase. However, this enhancement was likely to be less than the effect exerted by XNR_5179 and XNR_4478 on enzyme activity.
The heterologous expression of genes pntI, Fdx, and FDR in E. coli showed specific recognition between P450 and its auxiliary proteins and that only suitable auxiliary proteins can increase the efficiency of P450-catalyzed reactions. This also confirmed that P450 has stringent compatibility requirements for Fdx, with which it interacts directly, whereas the compatibility requirements between Fdx and FDR are relatively less stringent. PntI, XNR_5179, and XNR_4478 from Streptomyces were not soluble in E. coli; therefore, we believe that it is possible to improve the catalytic activity of PntI by increasing protein solubility through gene codon optimization. Adjusting the organization of genes and replacing the promoter and RBS with those of different strengths will change the ratio of the three proteins in the cell, which may further improve the catalytic activity of PntI.
In conclusion, as shown via gene knockout and heterologous expression in Streptomyces spp. and E. coli, pentalenene oxygenase can catalyze the production of 6 from 3, filling the only remaining gap in the biosynthetic pathway of 1. Hence, FdxG and FprD are considered suitable electron transport chain proteins for pentalenene oxygenases.
Data Availability Statement
All data generated or analysed during this study are included in this published article and its supplementary information files or are available upon request.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sono M.Roach M.P.Coulter E.D.Dawson J.H.Heme-containing oxygenases Chem. Rev.9619962841288710.1021/Cr 950050011848843 · doi ↗ · pubmed ↗
- 2Bernhardt R.Cytochromes P 450 as versatile biocatalysts J. Biotechnol.124200612814510.1016/j.jbiotec.2006.01.02616516322 · doi ↗ · pubmed ↗
- 3Urlacher V.B.Girhard M.Cytochrome P 450 monooxygenases in biotechnology and synthetic biology Trend. Biotechnol.37201988289710.1016/j.tibtech.2019.01.00130739814 · doi ↗ · pubmed ↗
- 4Cane D.E.Rossi T.Pachlatko J.P.Biosynthesis of pentalenolactone Tetrahedron. Lett.197936393642
- 5Pfeiffer F.Mecke D.Purification of pentalenolactone-insensitive glyceraldehyde-3-phosphate dehydrogenase from the producer strain streptomyces arenae H-S Z. Physiol. Chem.3611980314
- 6Cane D.E.Harrison P.H.M.Lamberson C.R.Oliver J.S.Sohng J.K.Biosynthesis of pentalenolactone - isolation of the pentalenene synthase gene Abstr. Pap. Am. Chem. S.2031992390 ORGN
- 7Chen L.Y.Deng Q.Ma T.T.Gu J.Y.Yang J.Zhang X.Zou Y.Q.Deng Z.X.Chen L.Zhao C.M.Substrate-dependent mechanism switch in the desaturation reactions of the mononuclear nonheme iron enzyme Ptl DACS Catal.1420247389740110.1021/acscatal.4c 00757 · doi ↗
- 8Tetzlaff C.N.You Z.Cane D.E.Takamatsu S.Omura S.Ikeda H.A gene cluster for biosynthesis of the sesquiterpenoid antibiotic pentalenolactone in Streptomyces avermitilis Biochem.-Us 4520066179618610.1021/bi 060419 n PMC 251862316681390 · doi ↗ · pubmed ↗