Postauricular Skin Mycobiome Profiles in Atopic Dermatitis Treated With Dupilumab or Cyclosporine A: A Descriptive Case Series
Yuta Koike, Hitomi Morisaki, Daisuke Motooka, Mai Matsumoto, Motoi Takenaka, Hiroyuki Murota

TL;DR
This study explores how two AD treatments, dupilumab and cyclosporine, affect the fungal makeup of the skin, particularly the Malassezia species.
Contribution
The study provides new insights into how systemic AD therapies alter the skin mycobiome, specifically Malassezia species.
Findings
Both treatments increased the abundance of Malassezia in the skin mycobiome.
There was a shift from M. globosa to M. restricta after treatment.
Patients with initially low Malassezia saw significant increases after treatment.
Abstract
Atopic dermatitis (AD) essentially exhibits dysbiosis of skin fungal microbiome, mycobiome, characterized by depletion of Malassezia. The effects of recent systemic therapies for AD on skin mycobiome were not understood enough. We examined changes of skin mycobiome before and after systemic treatments with anti‐IL‐4Rα antibody (dupilumab: DUP) and calcineurin inhibitor (cyclosporine, CyA). Swab samples from postauricular areas in 19 AD patients treated with dupilumab (n = 13) and cyclosporine (n = 6) were collected before and 4–8 weeks after starting each treatment. Fungal DNA was amplified from the samples and sequenced with ITS1 metagenomic analysis, and taxonomic classification was performed. Fungi belonging to total 89 genera were detected. The share of the fungus was most occupied by Malassezia (81.3%), followed by Aspergillus (3.7%), and Trametes (1.1%) before DUP and CyA…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
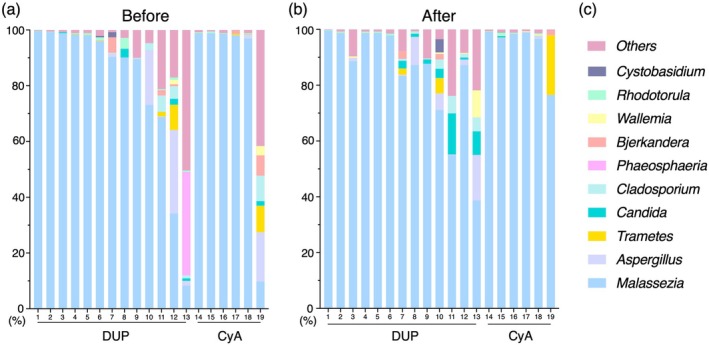 FIGURE 1
FIGURE 1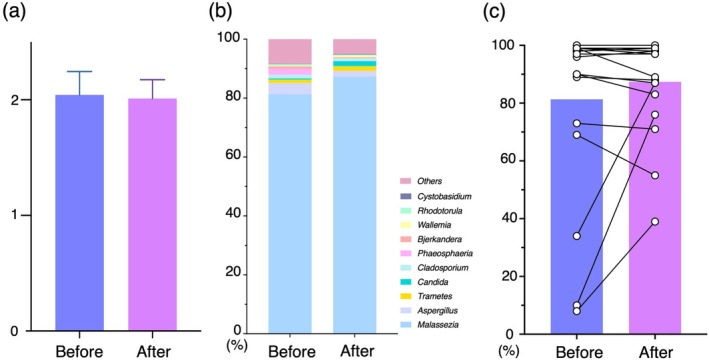 FIGURE 2
FIGURE 2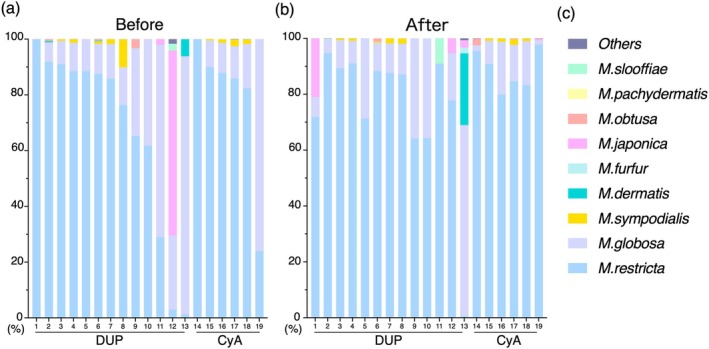 FIGURE 3
FIGURE 3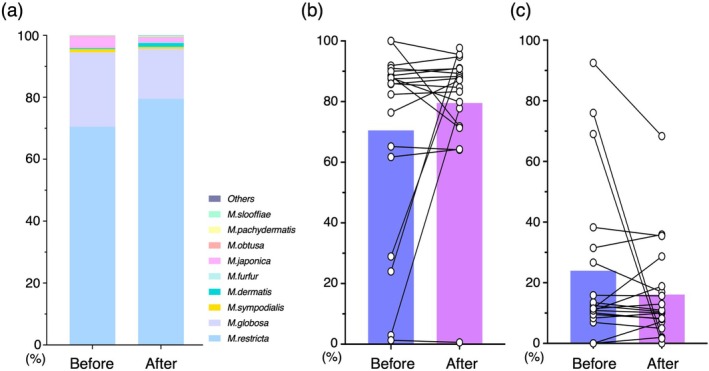 FIGURE 4
FIGURE 4| Patient numbers | Male/Female | Age | EASI | IgE | TARC |
|---|---|---|---|---|---|
| DUP ( | 10/3 | 41.8 ± 12.9 (18–68) | 27.2 ± 9.5 (16.4–45) | 10 769 ± 8435 (266–23 952) | 9039 ± 9291 (236–28 010) |
| CyA ( | 6/0 | 35.7 ± 11.9 (22–53) | 27.1 ± 13.2 (17.7–36.6) | 15 841 ± 13 586 (2316–37 602) | 5091 ± 2543 (2386–8343) |
| ALL ( | 16/3 | 39.8 ± 12.6 (18–68) | 27.1 ± 9.5 (16–45) | 12 354 ± 10 126 (266–37 602) | 7805 ± 7927 (236–28 010) |
- —Leading Medical Research Core Unit, Life Science Innovation, Nagasaki University Graduate School of Biomedical Sciences
- —Japan Society for the Promotion of Science10.13039/501100001691
- —Japan Agency for Medical Research and Development10.13039/100009619
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDermatology and Skin Diseases · Nail Diseases and Treatments · Psoriasis: Treatment and Pathogenesis
Introduction
1
The skin is inhabited by a diverse community of microorganisms, including bacteria, fungi, and viruses, which maintain human health by producing antibacterial peptides, forming biofilms, and inhibiting pathogen invasion [1]. Additionally, the skin microbiome comprises a complex ecosystem in which the physiochemical conditions produced by the resident microbiome interact with host biology [2]. When the intact skin microbiome breaks down, the abnormal microbiome condition is called dysbiosis, sometimes leading to the exacerbation of host skin diseases [3].
Atopic dermatitis (AD) is a common, chronic inflammatory skin disease, in which patients' quality of life are severely disturbed by inpatient itching [4]. The etiology of AD is multifactorial and has not yet been fully elucidated. One of the contributing factors that establish AD skin is dysbiosis of the skin bacterial microbiome characterized by a reduction in microbial diversity, overgrowth of Staphylococcus aureus , and a relative reduction of commensal species [5]. In recent years, the skin fungal microbiome, known as the name of skin mycobiome, has also been highlighted in skin diseases. The human skin mycobiome shows relative monotony, in which the genus Malassezia dominates the skin mycobiome, with only minor contributions from other fungi under steady‐state conditions [6]. The AD skin mycobiome is characterized by depleted amounts of Malassezia and a higher abundance of filamentous fungi [7].
In recent years, biological drugs targeting specific inflammatory cytokines have been used to improve inflammatory skin disorders, such as interleukin (IL)‐4, IL‐13, and IL‐31 for AD and tumor necrosis factor (TNF) α, IL‐17, and IL‐23 for psoriasis. These treatments not only attenuate skin inflammation activity but also influence the bacterial microbiome in humans. For example, AD treatment with an IL‐4RA inhibitor namely dupilumab (DUP) shifts the bacterial microbiome toward healthy skin flora [8]. Anti‐IL‐23p40 antibody therapy for psoriasis increased the β‐diversity of the skin bacterial microbiome to after treatment [9]. A patient with psoriasis who had been treated with an anti‐IL‐17A antibody developed AD‐like eczema in exchange for complete remission of psoriasis, which might be associated with skin bacterial dysbiosis [10]. Focusing on the skin mycobiome, a few research are carried out in psoriasis. Treatment with anti‐IL‐23p19 antibodies for psoriasis decreased the genus Malassezia on the antecubital fossa, whereas treatment with TNF‐α, IL‐17, and IL‐23p19 inhibitors did not affect the mycobiome on the postauricular skin [11, 12]. The features of skin mycobiome in Asian AD skin, which is characterized by not only Th2 but also Th17 signal involvement [13], and the effects of biological therapies for AD skin mycobiome were not sufficiently investigated.
Here, we conducted a descriptive analysis of the effects of systemic treatment with DUP and cyclosporine A (CyA) on the AD skin mycobiome. Swab samples were collected from postauricular skin before and after treatment DUP and CyA. Postauricular skin, a seborrheic and fungal‐rich skin, was selected as the sample collection site because this study aimed to elucidate the direct influence of systemic therapy on steady‐state AD skin. The effects of these treatments on skin mycobiomes were examined using ITS1 metagenomic sequencing. Finally, analysis of taxonomic diversity provided exploratory insights into the impact of systemic AD treatments on the skin mycobiome.
Methods
2
Study Subjects
2.1
Nineteen patients with AD in whom 13 treated with DUP, and six treated with CyA at the Department of Dermatology and Allergology, Nagasaki University Hospital, were enrolled. The participants were provided oral informed consent or the opportunity to opt out under protocols approved by the ethics committee of the Nagasaki University Hospital (22071112). The information of sex, current age, age at AD onset, past history, comorbidities, history of systemic treatment, blood test before treatment (counts for white blood cells and eosinophils, serum, IgE, and TARC), and the Eczema Area and Severity Index (EASI) score are retrospectively collected and are summarized in Table 1. The EASI score in a CyA‐treated patient and IgE and TARC in two DUP‐treated patients had missing data and were excluded from the Table.
Sample Collection
2.2
Dry swab samples were collected from each patient by rubbing the postauricular area over 50 times using culture swabs (BD BBD culture swab plus; BD, Japan) before treatment with DUP or CyA and after 4–8 weeks of treatment. There were no standardized conditions before sampling. The swab samples were stored at −80°C until DNA extraction.
Fungal ITS1 Deep Sequencing, Bioinformatic Analysis, and Taxonomic Assignment
2.3
Fungal ITS1 deep sequencing, bioinformatic analysis, and taxonomic assignment were performed as described previously [11]. Briefly, DNA was extracted from swab samples using the KURABO PI‐1200 DNA isolation system (Kurabo, Japan). Library preparation involved a two‐step PCR method with the primer set ITS1‐F (5′‐CTTGGTCATTTAGAGGAAGTAA‐3′) and ITS2 (5′‐GCTGCGTTCTTCATCGATGC‐3′) targeting the fungal ITS1 region, along with the Nextera XT Index Kit v2 (Illumina). Paired‐end sequencing of 301 bp amplicons was then carried out on a MiSeq instrument (Illumina). The resulting paired‐end reads were merged, filtered, and denoized using DADA2. Taxonomic assignment was conducted using the QIIME2 feature‐classifier plugin applied to the ntF‐ITS1 database, and all raw sequencing data were processed within the QIIME2 pipeline (version 2020.2) bioinformatics environment [14].
Statistical Analysis
2.4
Given the exploratory nature of this study, no formal hypothesis testing, sample size calculation or power estimation was performed. Data were summarized using descriptive statistics and visualized to illustrate variability without implying excessive precision. The α diversity (Shannon's diversity index) was calculated using the bioinformatics pipeline QIIME2 (version 2020.2). Results were expressed as mean ± standard error (SE). Data processing and visualization were performed using Microsoft Excel and GraphPad Prism 8 (GraphPad, San Diego, CA, USA).
Results
3
Skin Mycobiome of Individual Samples Before and After Treatment With DUP or CyA
3.1
The mycobiome on the postauricular skin of AD patients collected before and 4–8 weeks after treatment with DUP or CyA was analyzed. From the sequence results of all swab samples, a total of 89 genera of fungi were detected, of which the genus Malassezia had the highest average individual fungal composition (84.3%, Table S1). The details of fungal genera populations per sample derived from before treatment are shown in Figure 1a, and those after treatment with DUP or CyA are shown in Figure 1b, with the color bars corresponding to the top 10 genera detected before treatment (Figure 1c). Genus Malassezia is occupied over 50% on mycobiome in 16/19 samples before treatment and 18/19 samples after treatment.
Analysis of the skin mycobiome of atopic dermatitis patients was performed using swab samples collected from postauricular skin at two time points; (a) before and (b) after treatment with DUP and CyA. Numbers under the horizontal line are individual patients, 1–13 are treated with DUP and 14–19 are treated with CyA, corresponding before and after treatments. (c) Vertical bars represent the ratio of individual fungi in the mycobiome with fungal names matched to their color. CyA, Cyclosporine A; DUP, Dupilumab.
Genus Level Analysis of Skin Mycobiome Between Before and After Treatment
3.2
We integrated and comprehensively analyzed individual skin mycobiome data to identify the features after systemic AD treatment. An index of α diversity, the Shannon index, was similar before (2.04 ± 0.87) and after treatment (2.01 ± 0.70, Figure 2a). A β diversity analysis using PCoA with weighted UniFrac distance and PCA analysis, showed no consistent separation between the groups (Figure S1). A comparison of the mean ratios of individual fungi constituting the skin mycobiome both before and after treatment is shown in Figure 2b. The average percentages of the genera in the before‐treatment samples were as follows: Malassezia (81.3%), Aspergillus (3.7%), Phaeosphaeria (1.9%), Trametes (1.1%), and Candida (0.4%), and those in after‐treatment samples were as follows: Malassezia (87.3%), Aspergillus (1.9%), Candida (1.7%), Trametes (1.5%), and Cladosporium (0.9%). There were no specific genera whose average rates were clearly changed before and after treatment, whereas the genus Malassezia showed mild increase the relative abandance from before to after treatment (81.3% ± 6.9% and 87.3%± 3.8%, respectively). Three patients whose share of genus Malassezia in skin mycobiome was under 50% before treatment, showed an increase in the relative abandance of Malassezia after systemic treatment (average: before 17.3% ± 14.4%, after 67.3% ± 25.1%, Figure 2c). When separating the mycobiome results into DUP or CyA treatment groups, the average rate of genus Malassezia similarly increased after treatment in both groups (DUP, from 80.3% to 84.0%; CyA, from 83.4% to 94.3%; Figure S2). Thus, systemic treatment for AD was associated with a tendency toward increased the genus Malassezia in the skin mycobiome, especially in patients with a low composition of the genus Malassezia on their skin, which suggests a possible movement toward Malassezia‐dominant state.
(a) Averages of an α‐diversity index, the Shannon index, of the skin mycobiome in AD showed similar results before and after systemic treatments. (b) The average percentage of individual fungi showed a mild increase in Malassezia. (c) Composition of Malassezia in the skin mycobiome before and after systemic treatment is shown with bars (average) and paired dots (individual AD patients). AD, Atopic dermatitis.
Malassezia Species in Individual Samples Before and After Treatment With DUP and CyA
3.3
Next, we focused on the details of each Malassezia species in the genus Malassezia. The individual populations before and after treatment are shown in Figure 3a and Figure 3b, respectively, with the names of detected nine species of Malassezia (Figure 3c). Among samples before treatment, M. restricta is the most dominant in 15/19 samples (78.9%), followed by M. globosa in 3/19 samples, and M. japonica in 1/19 sample. Likewise, among the samples after systemic treatment, M. restricta was the most dominant in 18/19 samples (94.7%), and M. globosa was the most in only one sample. These results suggest that Malassezia species on the postauricular skin in AD patients are dominated by M. restricta, in which several samples especially before treatment disrupts the domination.
Analysis of Malassezia species in the postauricular area of atopic dermatitis patients is performed with samples taken (a) before and (b) after treatment with DUP and CyA. Numbers under the horizontal line are individual patients; 1–13 are treated with DUP and 14–19 are treated with CyA, corresponding before and after treatments. (c) Vertical bars represent the ratio of individual Malassezia species in the whole of Malassezia with species names matched to their color. CyA, Cyclosporine A; DUP, Dupilumab.
Treatments With DUP and CyA Modulate the Composition Balance Between M. Restricta and
M. globosa
3.4
The average ratios of the Malassezia species before and after treatment were shown in Figure 4a. M. restricta was the most dominant in samples before treatment (70.5%), followed by M. globosa (23.9%), M. japonica (3.5%), M. sympodialis (1.0%), and M. dermatis (0.3%). Similarly, M. restricta showed the most dominant after treatment (79.5%), followed by M. globosa (16.1%), M. japonica (1.5%), M. dermatis (1.3%), and M. sympodialis (0.5%). Focusing on the individual ratio of fungi, M. restricta increased its ratio (from 70.5% ± 31.8% to 79.5% ± 21.5%) and M. globosa decreased its ratio (from 23.9% ± 26.6% to 16.1% ± 16.3%) after treatment. When separating the treatment groups into DUP and CyA treatments, we obtained similar results on each treatment (Figure S3). Four patients whose ratio of M. restricta in total Malassezia species was under 50% tended to increase that of M. restricta after treatment (before 14.2% ± 7.1%, after 66.7% ± 22.4%, Figure 4b). Similarly, three patients whose ratio of M. globosa in Malassezia species was over 50% in the mycobiome tended to have decreased M. globosa after treatment (before 79.1% ± 6.9%, after 23.2% ± 22.5%, Figure 4c). Thus, DUP and CyA treatment for AD may be associated with modulation of the balance of Malassezia species characterized by an increase in M. restricta and a decrease in M. globosa , especially in patients with disrupted M. restricta domination to M. globosa on their skin.
(a) Average percentage of individual Malassezia species in the total Malassezia showed a mild increase in M. restricta and a mild decrease of M. globosa . Composition of (b) M. restricta and (c) M. globosa in the total Malassezia before and after systemic treatment is shown with bars (average) and paired dots (individual atopic dermatitis patients).
Discussion
4
In this study, we evaluated the skin mycobiome of AD patients systemically treated with DUP and CyA. Paired skin swab samples before treatment and 4–8 weeks after starting treatment were obtained and analyzed to describe the prevalence of different genera within the mycobiome. The results indicated that systemic treatments for AD were associated with tendency toward increased Malassezia in the skin mycobiome, especially in cases where the ratio of Malassezia was less than 50% before treatment. Focusing on Malassezia species, the ratio of M. restricta tended to increase after systemic treatment, in contrast to that of M. globosa which tended to decrease. No consistent patterns distinguishing DUP and CyA in the skin mycobiome were observed. Thus, systemic treatments for AD may be associated with the skin mycobiome characterized by a tendency toward increased Malassezia, especially M. restricta, indicating possible modulation of the skin mycobiome in AD skin.
Previous studies have suggested fungal dysbiosis in the skin of AD patients. The most characteristic fungal community of AD skin is the decrease in the genus Malassezia [7]. Since the genus Malassezia is essentially a dominant fungus for healthy skin, its depletion is an irregular condition for the skin that may allow the proliferation of pathogenic microorganisms and then prompt skin inflammation. Our results suggest that AD patients, whose genus Malassezia on the skin was depleted before treatment, can expand the genus Malassezia on their skin after treatment with DUP and CyA, which may be a modulation from the fungal dysbiosis of the AD skin. Genus Candida, that is usually live in the mucosal area on human, are found in several samples both before and after treatment. Candida is more frequently identified on AD skin than on healthy skin [15]. Our previous investigation of the skin mycobiome, in which we collected postauricular swab samples from 19 psoriasis patients using the same method of the current research, revealed that the genus Candida DNA was not detected at all even after treatment with anti‐TNFα antibody or anti‐IL‐17A antibody [12]. The presence of Candida in the postauricular skin might be a feature of AD skin that is not acquired even with immunosuppression by TNF or IL‐17.
The major component of Malassezia species on human skin is M. restricta, followed by M. globosa . These Malassezia species are commensal and usually not harmful to human skin, although overgrowth of these species sometimes causes skin diseases, such as seborrheic dermatitis and pityriasis versicolor [16]. The ratio of the predominant Malassezia species associates severity of AD lesions. Ratio of M. restricta is low and that of M. globosa is high in severe AD skin in comparison with healthy skin and mild AD skin [17, 18]. The inflammation mechanisms of Malassezia is thought to have several ways Malassezia can penetrate the defects in the skin barrier subsequently be recognized by pattern recognition receptors and initiate skin inflammation [19, 20]. In addition, the genus Malassezia has been suggested to provoke an allergic reaction and exacerbate the inflammatory response in AD [21]. Especially, M. globosa produces a protein namely MGL_1304 and dissolve it into human sweat. Recombinant MGL_1304 induces histamine release from basophils from patients with AD [22], which can explain the reverse phenomenon of the ratio of M. restricta and M. globosa in severe AD skin. Our results showed trends in which some swab samples from the AD skin showed a low ratio of M. restricta and a high ratio of M. globosa , and the ratios appeared to move toward a normal balance after systemic treatment for AD, suggesting possible modulation of Malassezia composition. These exploratory observations may imply a potential reduction of type 1 allergic responses related to proteins produced by M. globosa .
The variety of Malassezia species is also characteristic on AD. The relative abundances of M. dermatis and M. sympodialis were increased in AD lesions, and some species of Malassezia such as Malassezia slooffiae, Malassezia obtusa and Malassezia yamatoensis, were detected exclusively in the AD samples [23, 24]. Our results showed nine Malassezia species on the postauricular skin where is seborrheic and is relatively steady skin, while only four species were detected in psoriasis patients [12]. The multifarious inhabitant Malassezia species might be a feature of dysbiosis in the AD skin. However, our results did not show clear changes in the diversity of Malassezia species.
We observed no consistent differences in the skin mycobiome between the DUP and CyA treatments. A recent report on the skin bacterial microbiome revealed that DUP, but not CyA treatment, improved dysbiosis in patients with moderate‐to‐severe AD, in which S. aureus was reduced toward healthy control levels and α diversity was increased only in the DUP treatment group [8]. One of the reasons why DUP could improve the skin bacterial microbiome might be that DUP is an immunomodulator that ameliorates type 2 allergies in contrast to CyA as a broad immunosuppressant. The response of the skin microbiome to systemic treatment of AD might depend on the type of microorganisms, bacteria or fungi.
Our study provided the descriptive analysis regarding alterations in the skin mycobiome in patients with AD due on systemic therapies. However, this study had several limitations. This study did not include a healthy control group and AD group without systemic treatments. Generally, the skin mycobiome generally remains relatively stable in natural variations [25]. The relatively small cohort size may have limited the ability to detect subtle changes in the mycobiome such as the differences between DUP and CyA treatments. The sample collection timing after systemic treatments may affect the results of skin microbiome. A study of the skin bacterial microbiome revealed that systemic IL‐23 inhibition therapy for psoriasis initiated a gradual increase in the β‐diversity of the bacterial microbiome from before treatment to after 4 and 28 weeks of treatment [9]. In our current method, samples were collected 4–8 weeks after treatment which might be insufficient to alter the skin mycobiome. Regarding sample collection, washing of the sampling site might have affected the number of microorganisms [26]. Similarily sex and age distribution, disease severity, topical treatments, personal hygiene practices, and cleansing/face‐washing routines should have been standardized or adjusted. These factors can materially influence mycobiome profiles. Despite these limitations, our study provides descriptive insights into alterations of the mycobiome of AD skin after systemic treatment, serving as hypothesis generating evidence that warrants validation in larger, controlled cohorts.
Funding
This work was supported by the Leading Medical Research Core Unit, Life Science Innovation, Nagasaki University Graduate School of Biomedical Sciences. Japan Society for the Promotion of Science (25K11567). Japan Agency for Medical Research and Development (JP256f0137009).
Ethics Statement
Protocol was approved by the ethics committee of the Nagasaki University Hospital (22071112).
Conflicts of Interest
Yuta Koike, Motoi Takenaka, and Hiroyuki Murota received honoraria as a speaker or chair of lectures from Sanofi and Novartis.
Supporting information
Figure S1: β diversity analysis using PCoA with weighted UniFrac distance and PCA analysis.
Figure S2: Average proportion of fungal genera in DUP or CyA treatment groups.
Figure S3: Average proportion of Malassezia species in DUP or CyA treatment groups.
Table S1: List of detected fungal genera from skin swab samples.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Y. Belkaid and J. A. Segre , “Dialogue Between Skin Microbiota and Immunity,” Science 346, no. 6212 (2014): 954–959, 10.1126/science.1260144.25414304 · doi ↗ · pubmed ↗
- 2E. A. Grice and J. A. Segre , “The Skin Microbiome,” Nature Reviews. Microbiology 9, no. 4 (2011): 244–253, 10.1038/nrmicro 2537.21407241 PMC 3535073 · doi ↗ · pubmed ↗
- 3L. F. Koh , R. Y. Ong , and J. E. Common , “Skin Microbiome of Atopic Dermatitis,” Allergology International 71, no. 1 (2022): 31–39, 10.1016/j.alit.2021.11.001.34838450 · doi ↗ · pubmed ↗
- 4H. Murota , Y. Koike , H. Morisaki , M. Matsumoto , and M. Takenaka , “Exacerbating Factors and Disease Burden in Patients With Atopic Dermatitis,” Allergology International 71, no. 1 (2022): 25–30, 10.1016/j.alit.2021.10.002.34764038 · doi ↗ · pubmed ↗
- 5A. S. Paller , H. H. Kong , P. Seed , et al., “The Microbiome in Patients With Atopic Dermatitis,” Journal of Allergy and Clinical Immunology 143, no. 1 (2019): 26–35, 10.1016/j.jaci.2018.11.015.30476499 PMC 7163929 · doi ↗ · pubmed ↗
- 6K. Findley , J. Oh , J. Yang , et al., “Topographic Diversity of Fungal and Bacterial Communities in Human Skin,” Nature 498, no. 7454 (2013): 367–370, 10.1038/nature 12171.23698366 PMC 3711185 · doi ↗ · pubmed ↗
- 7R. Tao , R. Li , and R. Wang , “Dysbiosis of Skin Mycobiome in Atopic Dermatitis,” Mycoses 65, no. 3 (2022): 285–293, 10.1111/myc.13402.34817898 · doi ↗ · pubmed ↗
- 8J. Hartmann , L. Moitinho‐Silva , N. Sander , et al., “Dupilumab but Not Cyclosporine Treatment Shifts the Microbiome Toward a Healthy Skin Flora in Patients With Moderate‐To‐Severe Atopic Dermatitis,” Allergy 78, no. 8 (2023): 2290–2300, 10.1111/all.15742.37032440 · doi ↗ · pubmed ↗