Traffic and functional polarization of macrophages in the areas of programmed interdigital cell death in the embryonic chick
G. Moran, C. Duarte-Olivenza, J. M. Hurle, C. I. Lorda-Diez, J. A. Montero

TL;DR
The study explores how macrophages contribute to digit development in embryonic chicks through distinct populations and functional roles during tissue remodeling.
Contribution
The paper identifies two distinct macrophage populations with different origins and molecular profiles during embryonic digit development.
Findings
Resident macrophages precede interdigital cell death in embryonic limb development.
Macrophages exhibit an M2/anti-inflammatory gene signature during tissue remodeling.
M1-proinflammatory genes are upregulated at the end of tissue remodeling.
Abstract
In this study, we show that during digit development, the embryonic limb contains an abundant and widespread population of “resident macrophages” that precede the onset of interdigital cell death responsible for the separation of digit primordia. The use of cultures of interdigital mesoderm at different remodeling stages, and GFP+ transgenic embryos in which the distal part of the autopodium has been surgically replaced by a fragment with identical characteristics from wild-type embryos, indicate that “transient macrophages” are also recruited at the beginning of interdigital death. The expression of the Pu.1 gene in coincidence with the onset of interdigital death suggests that primary yolk-sac macrophages are later complemented by macrophages of hematopoietic origin. Q-PCR analysis revealed a predominant M2/anti-inflammatory gene signature in the interdigits during the whole…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
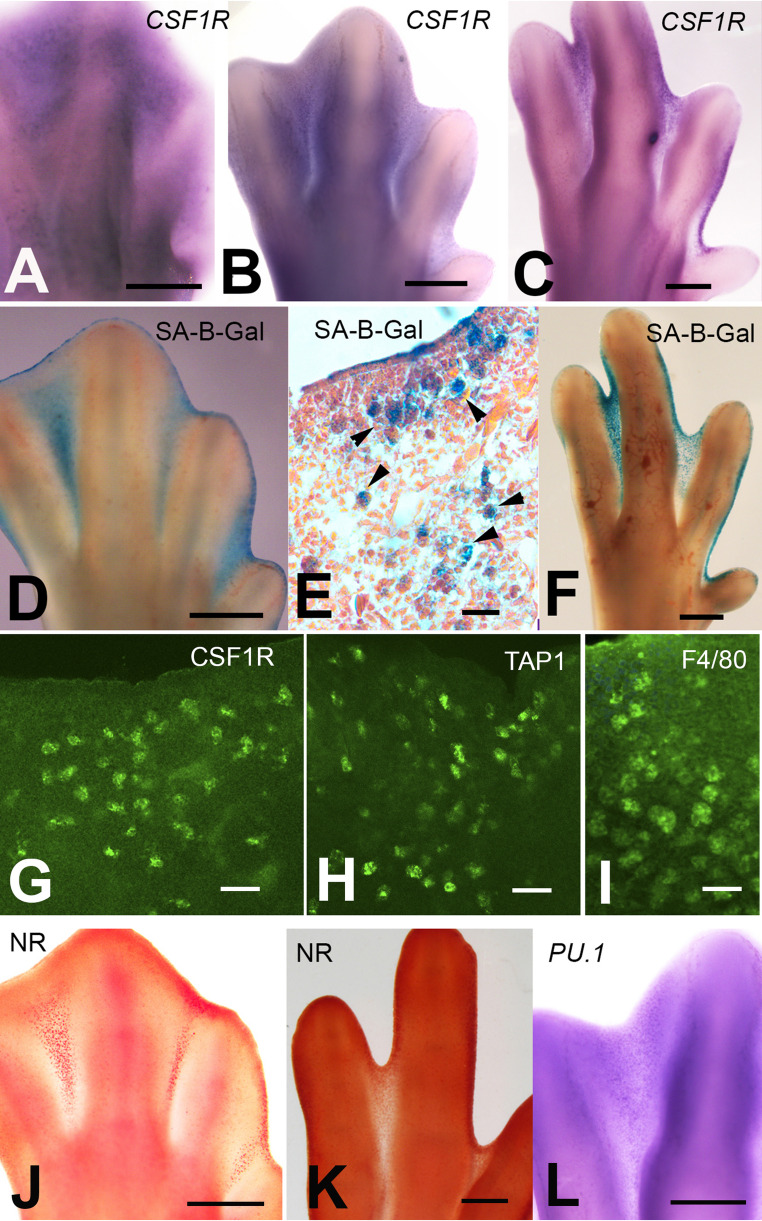 Figure 1
Figure 1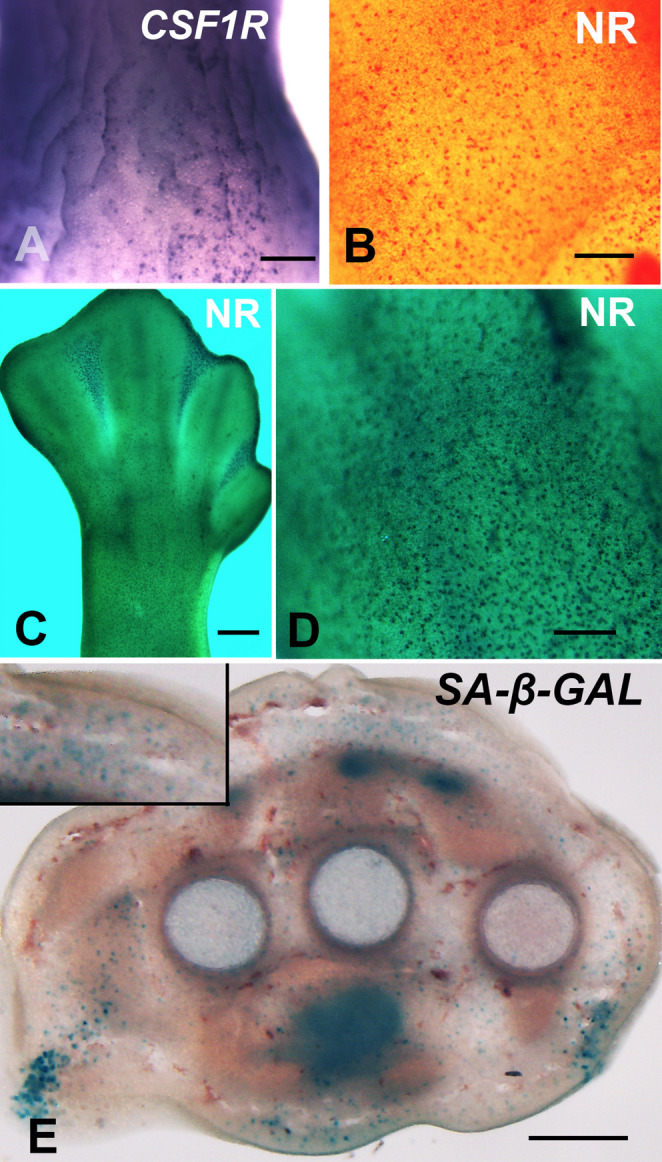 Figure 2
Figure 2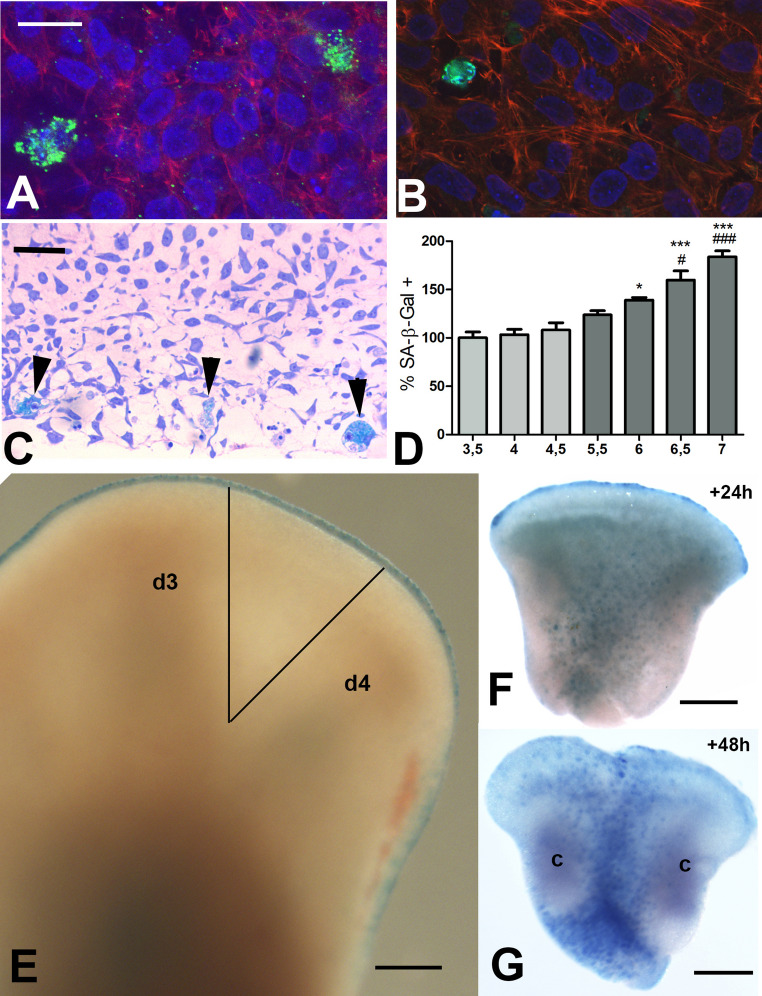 Figure 3
Figure 3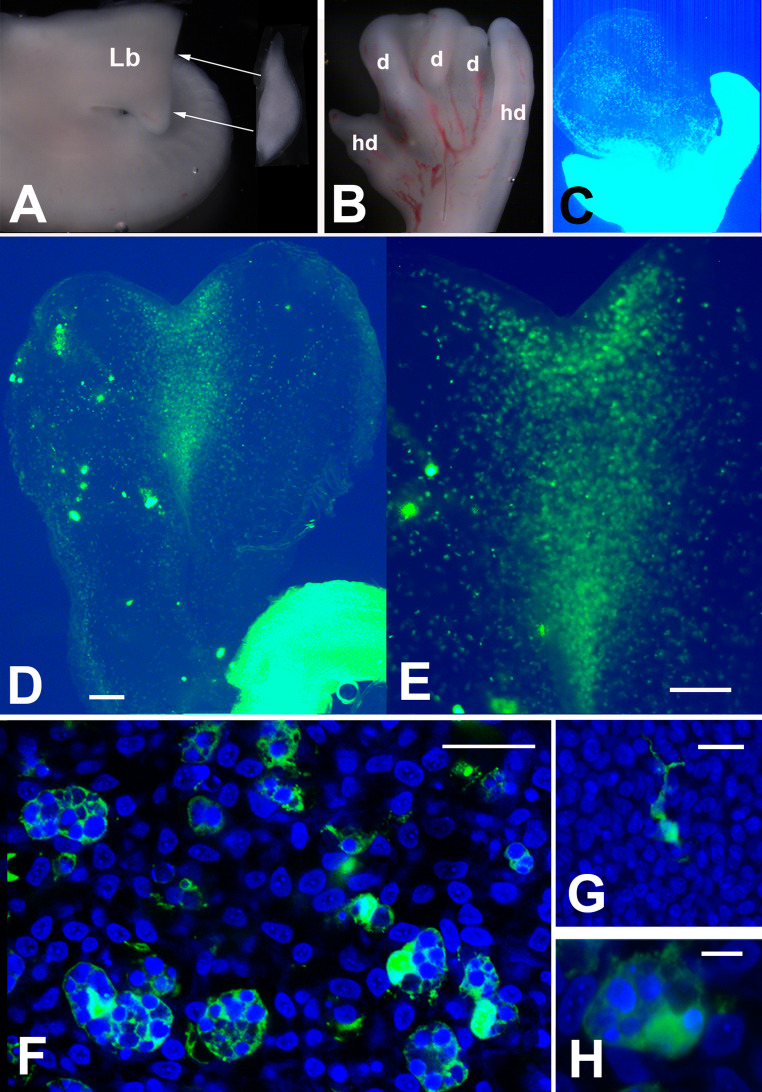 Figure 4
Figure 4- —Universidad de Cantabria
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPluripotent Stem Cells Research · Immune cells in cancer · Mesenchymal stem cell research
Introduction
Macrophages are defense cells, specialized in the ingestion and digestion of dead cells and endogenous or exogenous agents potentially dangerous for the organism as a whole, and represent a fundamental component of innate and adaptive immunity. Most, if not all, metazoans contain macrophage-like cell lineages with phagocytic functions [1, 2] Studies in adult systems revealed that the physiological functions of macrophages include many other aspects beyond phagocytosis [3, 4]. Thus, macrophages are fundamental physiological factors in the growth and postnatal maturation of the skeleton and other organs [5–7], maintain the structure and function of the cochlea in the auditory system [8], and participate in muscle contraction [9]. In addition, macrophages are involved in wound repair [10, 11] and regenerative processes [12, 13], and are able to modify the metastatic potential and response to treatments of several types of cancer [14, 15].
The development of macrophages in vertebrate embryos is a complex sequential process that generates variations in their functional properties associated with distinct specializations [5, 16, 17]. Pre-macrophages, and erythro-myeloid progenitors are initially formed in the yolk sac, undergoing successive migratory waves to colonize embryonic tissues and hematopoietic regions (fetal liver, the aorto-gonado-mesonephros region). From the fetal liver, a new wave of hematopoietic progenitors reaches the bone marrow that will be responsible for macrophage formation during the perinatal period and postnatal life [17–22]. During the migratory processes, yolk sac macrophages form “tissue resident macrophages” which proliferate locally and, eventually, differentiate into specialized functional subtypes [23].
The regenerative and trophic functions of macrophages are mediated by modulation of the tissue microenvironment [5, 7, 21, 24–27]. For this purpose, macrophages exhibit high secretory activity, which includes Non-Coding RNAs [28], growth factors [29], extracellular matrix remodeling molecules [30, 31]; and small signaling and immunomodulatory molecules (cytokines and chemokines). The transcriptomic and subsequent secretory profile establishes functional specializations of macrophages [32, 33] and allows the distinction of two extreme polarized states, the “pro-inflammatory” state (also called “classically activated” or “M1 macrophages”) and “anti-inflammatory/pro-regenerative” state (“alternatively activated” or “M2 macrophages”), and between them other molecular signatures associated with different functional activities [34].
Research in the last decades has shown that the expression of Csf-1R (c-fms gene) at high levels is a specific and, except for the placental trophoblast, exclusive marker of the macrophage lineage in vertebrates [26, 35, 36]. This gene reflects the developmental dependence of macrophage progenitors for the Colony Stimulating Factor 1 (CSF-1), and also Interleukin 34 (ligand of the same receptor; [37]. Several membrane receptors and transcription factors are also expressed by macrophages, but they are not as specific as CSF1R and their use as immunohistochemical markers of macrophages often requires the combination of more than one of them [4, 35, 38].
The significance and function of macrophages in the embryo remain poorly understood [3, 18, 27]. Although there are a few examples where macrophages appear to play a direct role in morphogenesis, removing transient cell components of organs in the course of morphogenesis [24, 39–42], the dynamics and local influence of macrophages in the embryo, other than removing degenerate cells, await clarification. This is particularly noticeable in processes involving programmed cell death that sculpt the shape and/or regulate the structure of many embryonic organs, where macrophages constitute a fundamental cell population [43, 44].
The objective of this study was to characterize the trafficking, phenotype, and functional polarization of macrophages in embryonic tissue-remodeling processes using the areas of interdigital apoptotic cell death of the embryonic limb (formerly termed Interdigital Necrotic Zones, INZ) as a model. Previous studies have shown that phagocytes in INZ are recruited from the yolk sac [45–47]. However, there is controversy regarding its arrival in the interdigits. It is not entirely clear whether it occurs before or after the onset of the apoptotic process [46, 48]. Most studies proposed that macrophage attraction was driven by stimuli originating from dying cells, but recent observations suggested that local macrophage arrival precedes interdigital apoptosis, proposing an active role in triggering interdigital death [48].
We identified Csf1r, SA-β-Gal, and Neutral Red positive “resident macrophages” beneath the limb ectoderm and around the skeletal primordia before apoptosis initiation. We further show that an additional population of “transitory” macrophages, expressing the hematopoietic transcription factor PU.1 (Spi1), becomes recruited coincidently with the onset of interdigital cell death. Q-PCR analysis revealed a predominant M2/anti-inflammatory gene signature in the interdigits during the whole remodeling process that correlated with the expression of a transcriptome that presumably stabilizes the macrophage M2 phenotype. However, M1-proinflammatory genes are up-regulated at the advanced stages of interdigit remodeling. Together, our findings are consistent with the proposed model of “layered macrophage system” that supports the existence of “Tissue-Resident” and “Passenger/Transitory” macrophage populations of different origin, and molecular profiles [17].
Materials and methods
Rhode Island chicken eggs (Granja Santa Isabel, Cordoba, Spain), and GFP-expressing Brown Leghorn chicken eggs [49] (Roslin Institute, Edinburgh, UK; a generous gift of Dr. MA Ros) were employed in the experiments. Fertilized eggs were incubated at 38 °C to obtain embryos ranging from 3,5 to 9 incubation days (id; stages 23HH to 35HH). Tissue samples for the study were dissected free from the leg bud following the ethical recommendations of the European Communities Council.
In Situ Hybridization (ISH)
PFA-fixed samples were treated with 10 µg/ml of proteinase K for 10–20 min at 20 °C. Hybridization with digoxigenin-labeled antisense RNA probes was performed at 67 °C. An alkaline phosphatase-conjugated anti-digoxigenin antibody (1:2000 dilution) (Roche) was used. Reactions were developed with a BCIP/NBT substrate (Roche). ISH antisense probes for chicken Csf1r (c-FMS) and Pu.1 (Spi1), were obtained by PCR employing the following primers:
Csf1r: Fwd. primer: 5´-AAGGTGGTAGAAGCCACTGC-3´ // Rev. primer: 3´-ACTGGAGCAGGTCAGAGAGG 5´.
Pu.1: Fwd primer: 5´CGGATGTGCCTACAGTACTCC-3´ // Rev. primer: 3´-CGTGCCATCTTCTGGTAGG-5´.
Mesodermal cultures
Mesodermal tissue was obtained from the distal region (progress zone) of chicken leg buds at 3.5–4.5 id or from the third interdigit of the leg buds at 5.5, 6, 6.5, and 7 id. Cells were dissociated, filtered through a 70-µm strainer (Miltenyi Biotec), suspended in DMEM-10% FBS-1% PenStrep antibiotic (Sigma), and cultured at high density (3 × 10^5^ cells/ml) as previously described in [50].
Explant cultures
The third interdigit of leg buds at 5.5 id was dissected free, explanted to culture medium (DMEN/F12 50/50 (Corning), 5% FBS; 5% Chick Serum, and 1% L-glutamine and 1% PenStrep antibiotic), and allowed to grow for 24–48 h, before processing for SA-β-Gal histochemistry or neutral red vital staining.
Tissue grafts
Distal regions (200–300 μm) of the leg bud from wild-type embryos were microdissected at day 4.5 id and used to replace identical regions excised from the limb bud of GFP-expressing host embryos, see Fig. 5A, and [51] for details. The chimeric embryos were incubated for 3–4 days and processed to monitor the entry of GFP-positive cells into the grafted region. Reverse grafting experiments of the distal mesoderm of GFP-positive embryos into wild-type hosts were also performed.
Histology, β-Gal activity, Neutral Red vital staining, and immunolabeling
Histological observations were made in PFA-fixed tissue sections obtained by a vibratome or from paraffin wax-embedded or plastic-embedded material. Hematoxylin/eosin or toluidine blue staining was employed.
The β-Galactosidase activity assay [52] was performed at pH 6 in whole-mount or tissue sections (vibratome or paraffin wax-embedded) of limb autopods fixed in glutaraldehyde.
Immunolabeling for CSF1-R (Colony Stimulating Factor 1 Receptor; MCA5956GA, Bio-Rad), TIM4 (T-cell immunoglobulin mucin receptor 4; MCA6407GA, Bio-Rad), F4/80 (Adhesion G protein-coupled receptor E1; HS-397 004, HistoSure, Synaptic Systems), and TAP1 (Transporter 1, ATP binding cassette subfamily B member; Hybridoma Bank) were performed, see [53] for details. These antibodies were incubated with specific secondary antibodies (Jackson). Counterstaining was performed using rhodamine-phalloidin (Sigma) and/or DAPI (Vector Laboratories). Observations were made with a laser confocal microscope (LEICA LSM 510).
Neutral red (NR) vital staining was performed, in unfixed material, with a solution of 0.2% of NR (0.2 N) in PBS at 38 °C and controlled under the binocular microscope, as described in [54].
Real-time quantitative PCR (Q-PCR) for gene expression analysis
Total-RNA was extracted from samples, each composed of 12 interdigits, using the NucleoSpin RNA kit (Macherey-Nagel) and quantified using a spectrophotometer (Nanodrop Technologies ND-1000). cDNA was generated using random hexamers with the High-Capacity cDNA Reverse Transcription Kit (Life Technologies). The concentration was adjusted to 0.5 µg/µl. Q-PCR was performed using the CFX Connect Real-Time System (BioRad) employing SYBR Select Master Mix (Life Technologies). Rpl13 was chosen as the normalizer. Expression fold changes were evaluated relative to controls according to the 2^−(ΔΔCt)^ equation. Statistical analyses were performed using ANOVA followed by the Bonferroni test for post-hoc comparisons. Supplementary Table 1 shows Q-PCR specific primers for chick genes.
Results
“Tissue-resident macrophages” occupy the subectodermal and perichondrogenic regions of the limb and concentrate in areas of programmed cell death
Previous studies have shown that NR staining and SA-β-Gal histochemistry are markers of activated macrophages present in INZ [48, 53]. Here, we have employed in situ hybridization for the Csf1r gene (c-Fms), in combination with NR staining and SA-β-Gal histochemistry to map the distribution of “resident macrophages”. As shown in Fig. 1A-F and J-K the three procedures marked in the same fashion the areas of cell death in the remodeling interdigits. The macrophagic nature of these interdigital cells were also confirmed by immunolabeling with macrophage markers, including CSF1R, TAP1, and F4/80 (Fig. 1G-I). Fig. 1. Expression of macrophage markers in the areas of interdigital cell death. A-C In situ hybridization for Csf1r at 6.5 (A), 7.5 (B), and 8 (C) id. Note a tenuous labeling under the ectoderm at 6,5 id that becomes concentrated in the interdigits at 7.5. (B) and 8 (C) id. D-F Histochemical labeling for SA-β-Gal at 7 (D-E) and 8 (F) id showing intense labeling in the interdigits. (E) Hematoxylin-eosin-stained histological section of the third interdigit to show the distribution of SA-β-Gal labeling in macrophages (arrow heads). G-I, vibratome sections of the third interdigit at 7id, showing positive immunolabeling for CSF1R (G), TAP1 (H), and F4/80 (I) macrophage markers. , J-K Neutral Red vital staining of autopods at 7 (J) and 8 (K) id. L In situ hybridization for Pu.1 to show the weak dotted labeling of the third interdigit at 7.5 id. Scale bars in A-D and F, J, K and L = 200 μm; E, G, and H = 100 μm; I = 60 μm
Considering the molecular and functional heterogeneity and the distinct origin of macrophage progenitors in the embryo [55], we characterized the transcriptional profile of macrophage markers in INZ (see Table 1). The gene for the receptor Csf1r was selected as a general marker of macrophages [26], the receptor Cx3cr1 for its implication in the migration of macrophages to colonize the embryonic tissues [56–58], and the transcription factor Pu.1 to mark macrophages of hematopoietic origin [35]. The expression of Csf1 (macrophage colony-stimulating factor) and Il-34 were also analyzed. CSF-1 is a growth factor required for the maintenance of tissue-specific macrophage populations [59] and together with IL-34 are CSF1R ligands [37]. As shown in Table 1, the Csf-1 gene maintains a high and uniform expression in the interdigit during the remodeling process, while Il-34 is expressed at low levels but up-regulated at advanced remodeling stages. In human inflammatory diseases, Csf-1, unlike granulocyte-macrophage colony-stimulating factor (Gm-csf (Csf2); not expressed in the interdigit), has been shown to maintain the M2 phenotype of macrophages [60].Table 1. Expression and regulation of macrophage gene markers (Csf1r; Cx3cr1; Pu.1) in the interdigital tissue before (5.5 id) at the onset (6.5 id), and at the peak (7.5 id) of cell deathGeneID3 5,5 idID3 6,5 idID3 7,5 idCsf1r1,04 ± 0,321,24 ± 0,614,99 ± 2,36Cx3cr11,01 ± 0,181,98 ± 0,9611**,96 ± 6,81Pu.11,01 ± 0,155**,73 ± 2,2410,91 ± 4,33CSF1R ligandsCsf11,01 ± 0,171,03 ± 0,201,16 ± 0,12Il-341,02 ± 0,221,28 ± 0,442,09 ± 0,47Data were analyzed using ANOVA followed by the Bonferroni test for post hoc comparisons. n = 8. () p-value < 0.001
Consistent with the Csf1r expression pattern observed by in situ hybridization, Q-PCR analysis of limb mesoderm revealed the presence of Csf1r transcripts in the undifferentiated distal mesoderm before the appearance of digital condensations and in the interdigital tissue when digit rays become identified (5.5 id) but 24 h before the onset of cell death (Table 1 and Supplementary Fig. 1). At later stages, when cell death reaches its highest intensity, Csf1r expression increased up to 5-fold at 7.5 id (Table 1). Cx3cr1 shows abundant transcripts in the interdigits (Supplementary Fig. 1) and becomes up-regulated (12-fold) at advanced stages of tissue remodeling (Table 1). In contrast to these genes, expression of the hematopoietic macrophage marker Pu.1 was almost absent prior to the onset of cell death but became up-regulated from day 6.5, when cell death commences (Table 1 and Supplementary Fig. 1). In subsequent periods, expression is up-regulated up to 10-fold (7.5 id). In situ hybridization confirmed the absence of PU.1 expression domains in earlier stages of digit development, but a faint expression in the macrophages was identified by 7 id (Fig. 1L), suggesting the recruitment of hematopoietic macrophages positive for this marker once cell death is established. Csf1 expression is stable during the remodeling process, but Il-34, although expressed at low levels, is up-regulated in the advanced remodeling period (Table 1 and Supplementary Fig. 1).
An additional remarkable observation was that, unlike canonical descriptions of areas of cell death, careful microscopic observation of the limbs revealed that Csf1r /NR/SA-β-Gal-positive cells are not exclusively arranged in INZ. A significant number of positive cells show a broader distribution within the limb tissues before and during the establishment of the interdigital cell death domains. As shown in Fig. 2A-D, Csf1r/NR/SA-β-Gal positive cells showed a mottled distribution beneath the ectodermal layer of the zeugopod, proximal autopod, and digit rays. In vibratome sections, these cells are also observed around the musculoskeletal primordia of the autopod (Fig. 2E).
Fig. 2“Resident Macrophages” in the developing limb labeled with Csf1r ISH A, Neutral red vital staining B,** C**,** D**, and SA-β-Gal histochemistry E. A ISH for Csf1r gene showing the widespread punctate distribution under the ectodermal surface of the zeugopod at 4.5 id. B Detailed view of the dorsal surface of the zeugopod vital stained with NR. C,** D** Micrographs, using a green filter to enhance contrast, showing the pattern of NR staining of the leg bud at 7.5 id. (C) shows a low-magnification view of the limb, and (D) a detail view of the dorsal surface of the zeugopod. Note the intense staining of the areas of interdigital death in (C), and the abundance of NR-positive cells in the subectodermal space in (D). E Transverse vibratome section of the autopod at 7 id, after SA-β-Gal histochemistry, showing the abundance of SA-β-Gal+ cells in the subectodermal space and around the metatarsal primordia. The inset shows a detailed view of the subectodemal region. Scale bars: A = 500 μm; C = 150 μm; B and D = 300 μm
Local differentiation and interdigital recruitment of macrophages
The accumulation of macrophages in the interdigits was analyzed employing micromass cultures of dissociated interdigital mesoderm. The micromass culture is an organoid-like assay that replicates the events occurring in vivo [50]. Macrophages in the culture were identified by CSF1R (Fig. 3A) and TIM4 (Fig. 3B) immunolabeling, and by histochemical detection of SA-β-Gal (Fig. 3C), which was employed for microscopic quantification (Fig. 3D). We quantified the number of SA-β-Gal-positive macrophages in micromasses obtained from the distal limb mesoderm (progress zone) prior to the formation of digit primordia (3.5, 4, and 4.5 id.) and from the third interdigit of limbs at 5.5, 6, 6.5, and 7 id. Macrophages were present in all samples, but their number increased according to the stage of the donor tissue (Fig. 3D). Considering that macrophages in micromasses are related to those present in the limb tissue at the time of obtaining the samples for culture, this observation is consistent with a progressive recruitment of macrophages to the limb autopod and/or the occurrence of local proliferation of pre-macrophages already present in the tissue. This feature was further evaluated in explant cultures of the interdigit 3 isolated surgically at 5.5 id (Fig. 3E), which precedes in 24 h the onset of cell death. In the absence of the adjacent digits, the explant, instead of degenerating, grows and forms one or two digit-like chondrogenic elongated nodules (see Omi et al., 2000). As shown in Fig. 3F-G, SA-β-Gal positive cells were very abundant in the explants after 24 (Fig. 3F) and 48 h (Fig. 3G) of culture, supporting the presence of pre-macrophages in the interdigits, capable of differentiating into active macrophages. Remarkably, macrophages are concentrated at the proximal end of the interdigit and between the two cartilage aggregates that are formed in the explant.
Fig. 3. Presence of macrophages in micromass cultures of undifferentiated leg bud mesoderm from 3.5 id embryos. A Three-day-old micromass immunostained for CSF1R (green). B, Three-day-old micromass immunostained for TIM4 (green). C Toluidine-blue-stained 1 μm thick section showing intense SA-β-Gal positivity in macrophages (arrows) in a three-day-old micromass. D Graphic representation of the relative number of SA-B-Gal+ macrophages in micromass cultures obtained from leg buds at 3.5, 4, and 4.5 id (faint gray columns) and from interdigits of 5.5, 6, 6.5 and 7 id (dark grey columns). Data was analyzed using ANOVA followed by the Bonferroni test for post hoc comparisons. N = 4. Statistical significance vs. 3.5 id micromasses: *p < 0.05; ***p < 0.001. Statistical significance vs. 5.5 id interdigits: # p < 0.05; ### p < 0.001. The number of macrophages in cultures from the limbs of 3.5 id was considered 100%. E-G SA-β-Gal histochemistry to label macrophages in cultured interdigit explants. (E) Low-magnification view of the autopod at the time of obtaining the interdigit for explant culture. Note the absence of labeling in the third interdigit located between digits 3 (d3) and 4 (d4), and a slight positivity of autophagic vacuoles in the periderm layer of the distal ectoderm bordering the limb bud (termed the AER). The lines mark the microdissected interdigital region for the explants. (F) Interdigit after 24 h of culture. Note the absence of labeled macrophages in the distal region of the interdigit subjacent to the AER, in contrast to the abundance of SA-β-Gal+ cells in the proximal tip and in the central zone of the explant. (G) Interdigit explant after 48 h of culture. Note the higher number of macrophages compared to F, and their predominant arrangement between the cartilage-forming zones (c) and at the proximal tip of the explant. Scale bars: A and B = 15 μm; C = 50 μm; E to G = 60 μm
Fig. 4GFP+/GFP- chimeric limb experiments. A Illustration of the surgical replacement of the distal undifferentiated mesoderm from a wild-type autopod in a GFP+ host embryo of the same stage. Arrows indicated the implantation of the wild-type autopod fragment into a host GFP + leg bud (Lb). B Autopod morphology 3.5 days after tissue graft illustrated in (A). The digits at the autopod margins (hd) are host –derived, while the central digits (d) are graft-derived. C Histological section of the graft illustrated in B, under fluorescent microscopy. Note the intense fluorescence in the digits at the autopod margins, in contrast to the graft-derived central digits, and the absence of massive labeling in the grafted tissue. D Low-magnification view of a section through two adjacent graft-derived digits. Note that positive GFP labeling is concentrated in the interdigit region. E Section of chimeric limb showing the selective distribution of GFP+ cells in the interdigits. F Detailed view of the interdigit of an experimental chimeric limb stained with DAPI showing selective GFP positivity of large macrophages. G-H Detailed view of GFP+ host cells located outside (G) and in the interdigits (H). Note the protrusive morphology of the migrating host cells outside the areas of cell death, compared to the characteristic macrophage morphology of the cells that migrated to the interdigits. Scale bars in D and E = 100 μm; F = 50 μm; G= 25 μm; H= 10 μm
To analyze the recruitment of macrophages from proximal limb regions, we generated chimeric limbs by grafting a distal limb fragment from wild-type embryos into transgenic GFP+ host embryos at 4.5 id. For this purpose, a strip of tissue (200–300 μm-thick) was removed from the leg bud of GFP+ host embryos at 4.5 id and replaced with an identical fragment dissected from wild control embryos at the same stage (Fig. 4A). After 3 to 3.5 days of additional incubation, the chimeric limb was removed (Fig. 4B), fixed, and sectioned for subsequent observation (Fig. 4C). As shown in Fig. 4D-E, an intense population of GFP-positive cells was detected in the grafted tissue, occupying the interdigital areas, the peridigital mesoderm and the capillaries. High magnification confocal observation (Fig. 4F and H) identified GFP+ interdigital cells as macrophages showing numerous cytoplasmic vacuoles containing GFP-negative cell debris including DAPI positive picnotic nuclei. (Fig. 4G), shows the protrusive morphology of host GFP+ cells that did not reach the interdigit region of the grafted tissue. Mirror experiments implanting GFP+ grafts in wild-type embryos discarded the occurrence of distal-proximal cell migration. The suture edge between the host and the graft was well established, and GFP+ cells were almost absent from the host tissue (not shown).
Interdigital macrophages share M1 and M2 phenotypic features
Macrophages show considerable genetic diversity associated with their development and distribution in the body [19, 61]. Additionally, in the different locations, macrophages display dynamic transcriptomic profiles that reflect a variable and sometimes opposite functional activity, dependent on the context [62]. Here, we characterized the transcriptional signature of the interdigital tissue and the changes that occur during the remodeling process. Depending on local signaling cues, undifferentiated macrophages undergo polarization between two extreme functional states of active macrophages, M1 and M2, with several intermediate subtypes [63–66].
Our Q-PCR transcriptional study showed a complex pattern of expression of M1 and M2 macrophage markers with significant changes in the course of interdigit remodeling (Tables 2 and 3; and Supplementary Fig. 1). As shown in Table 2 and Supplementary Fig. 1, M1 markers showed variable patterns of expression. Prior to the onset of cell death, Tnfα is expressed at very low levels but from 6.5 id undergoes intense and progressive up-regulation, achieving the highest values at 7.5 id. Tlr4, Mhc-II, and Interleukin-16 (Il-16) follow a similar pattern of expression to Tnfα but maintain higher expression levels. Inos,* Notch1*, and Foxo1 maintain high levels of expression during periods of remodeling, but they were not significantly regulated. Other M1 markers, such as the inflammatory cytokines Interleukin-1β (Il-1β), Interleukin-6 (Il-6), Ifn-β, and Ifn-γ (not included in Table 2), are not expressed or expressed at low levels. Distinctly, the M1 transcription factor Stat1 is expressed at a high level, but its expression becomes moderately down-regulated at advanced stages of tissue remodeling (Table 2).
Table 2. Expression and regulation of M1-macrophage gene markers in the interdigital tissue before (5.5 id), at the onset (6.5 id), and at the peak (7.5 id) of cell death. Note the intense up-regulation of Tnfα and Tlr4GeneID3 5,5 idID3 6,5 idID3 7,5 id Tnfα 1,02 ± 0,212,50 ± 1,416,07 ± 2,89** Il-8 (L1) 1,013 ± 0,193,035 ± 1,1231,52 ± 23,49* Il-6 1,004 ± 0,110,705 ± 0,251,698 ± 0,63 Il-1β 1,005 ± 0,120,346 ± 0,100,734 ± 0,25 Il-16 1,018 ± 0,221,061 ± 0,382,775 ± 1,77 Tlr4 1,042 ± 0,352,072 ± 0,7617,18 ± 8,61*** Mhc-II 1,021 ± 0,242,098 ± 0,482,464 ± 1,13 Inos 1,012 ± 0,170,766 ± 0,290,820 ± 0,43 Stat1 1,07 ± 0,460,587 ± 0,170,516 ± 0,11 Ifnβ 1,002 ± 0,070,478 ± 0,240,881 ± 0,28 Foxo1 1,01 ± 0,151,11 ± 0,461,72 ± 0,63 Notch1 1,01 ± 0,110,59 ± 0,190,65 ± 0,37Data were analyzed using ANOVA followed by a Bonferroni test for post hoc comparisons, n = 6. () p-value < 0.05, () p-value < 0.01, () p-value < 0.001 Table 3. Expression and regulation of M2-macrophage gene markers in the interdigital tissue before (5.5 id) the onset (6.5 id) and at the peak (7.5 id) of cell deathGeneID3 5,5 idID3 6,5 idID3 7,5 id Mrc1 (Cd206) 1,081 ± 0,480,69 ± 0,280,66 ± 0,21 Il-10 1,033 ± 0,300,43 ± 0,270,34 ± 0,25 Il-4 1,022 ± 0.220,80 ± 0,301,15 ± 0.36 Il-4r 1,032 ± 0,301,72 ± 0,671,15 ± 0,25 Stat6 1,06 ± 0,391,12 ± 0,701,41 ± 0,56 Stat3 1,01 ± 0,141,92 ± 1,061,91 ± 0,92 Klf4 1,01 ± 0,152,48 ± 0,875,61 ± 3,35** Irf4 1,04 ± 0,302,00 ± 1,561,43 ± 1,01Data were analyzed using ANOVA followed by a Bonferroni test for post hoc comparisons, n = 6 (**) p-value < 0.01
A notable disparity in the pattern of gene expression for M2 markers in relation to observations in adult organisms was also observed (Table 3 and Supplementary Fig. 1). The characteristic M2 marker, mannose receptor Mrc1 (Cd206), is expressed at a considerable level before the onset of cell death but becomes moderately down-regulated as degeneration progresses. The Krüppel-like factor 4 (Klf4), Il-4r, and the transcription factors Stat6 and Stat3 were expressed at high levels through the remodeling process, with Klf4 showing a significant up-regulation at advanced stages of remodeling. Irf4 maintains moderate expression levels through the remodeling period. Il-4 maintains a low expression level without significant changes in the course of remodeling. Il-10, a further anti-inflammatory cytokine, is also expressed at very low levels, showing decreased expression in interdigits of 7 and 7.5 id. In contrast to those M2 markers, Il-13, which, together with Il-4 and glucocorticoids, is a canonical inducer of M2 polarization, is not expressed in the regressing interdigit.
To further characterize the molecular signature of macrophages, we also analyzed the expression and regulation of non-specific M1 or M2 markers. Cd45, Pparγ, C/ebpβ, Runx1, Pd-l1, and Ccr2 were all expressed at intermediate levels, becoming up-regulated at advanced stages of interdigit remodeling (Table 4). In contrast, other markers, including Tap1, Cd11b, and c-Myb, were also expressed at intermediate levels but were not up-regulated during interdigit remodeling.Table 4. Expression and regulation of non-specific M1/M2 macrophage markersNo M1/ M2 specific macrophage markersGene5,5 id6,5 id7,5 idTap11,06 ± 0,401,38 ± 0,431,76 ± 0,93Cd-451,04 ± 0,330,89 ± 0,232,11 ± 0,56Cd11b1,02 ± 0,191,27 ± 0,390,94 ± 0,46c-Myb1,00 ± 0,080,89 ± 0,421,01 ± 0,52Pparγ1,02 ± 0,092,83 ± 0,495**,32 ± 2,90C/ebpβ1,00 ± 0,071,49 ± 0,633**,39 ± 2,76*Runx11,05 ± 0,343,84 ± 0,886,98 ± 2,47Pd-l11,01 ± 0,151,08 ± 0,502**,74 ± 1,08*Ccr21,05 ± 0,371,30 ± 0,804,82 ± 2,38Data were analyzed using ANOVA followed by a Bonferroni test for post hoc comparisons, n = 6. () p-value < 0.05, () p-value < 0.01, (*) p-value < 0.001
Molecular signature of the regressing interdigital tissue
The interdigits are transient structures composed of skeletal progenitors [67], capillaries [68], the tissue-resident macrophages described here, and a cover of ectoderm. The fate of these components in species with separated digits, such as the chicken, is their complete elimination within a short developmental period (60–70 h in the chicken embryo). In a previous study, we observed that, despite its transient nature, the interdigits contain abundant signaling molecules that were pointed out as evidence of a senescence-associated phenotype (SASP) [69]. However, it is now believed that senescent cells, previously considered as such, actually represent tissue-resident macrophages [48, 53], and many SASP signaling factors are shared by immune defense processes [70]. Considering that in adult systems the local microenvironment plays an active role in macrophage activation and polarization [11] we explored the interdigital expression of factors potentially relevant to modulating the macrophage phenotype. We detected significant expression levels (Supplementary Fig. 2) of several growth factors and receptors capable of regulating the polarization of macrophages in adult systems, including members previously considered SASP components. As shown in Table 5, the panel of regulatory factors analyzed included Igf1, Igfbp5, Tgfβ4 (Tgfβ1 in mammals), P75^NTR^/ Tnfrsf16, Hgf, and Vegfa. Except for Hgf and Vegfa, all of them were up-regulated in the course of interdigit remodeling.Table 5. Expression and regulation of growth factors expressed in the interdigits in the course of tissue remodelingGeneID3 5,5 idID3 6,5 idID3 7,5 idTgfβ41,01 ± 0,161,12 ± 0,401,94 ± 0,85*Igf11,08 ± 0,485,30 ± 1,537,40 ± 4,43Igfbp51,00 ± 0,043**,18 ± 0,509,62 ± 0,61P75^NTR^1,01 ± 0,161,88 ± 0,694**,01 ± 0,76Vegfa1,04 ± 0,360,79 ± 0,400,45 ± 0,25Hgf1,02 ± 0,211,59 ± 0,891,24 ± 0,37Data were analyzed using ANOVA followed by a Bonferroni test for post hoc comparisons, n = 6. () p-value < 0.05, (*) p-value < 0.001
TGFβ4 (TGFβ1 in mammals) is a member of the TGFβ family produced by macrophages after phagocytosis of apoptotic cells. It is expressed throughout the entire remodeling process. Studies in different systems showed a function for this cytokine promoting M2 anti-inflammatory macrophage phenotype [71].
IGF1 is a macrophage autocrine promoter of M2 polarization that functions together with its binding protein IGFBP5 in a balanced and antagonistic manner [72, 73]. Both factors are expressed in the interdigits, reaching their highest levels at most advanced stages of tissue remodeling.
P75^NTR^ (TNFRSF16) has long been characterized as a Tumor Necrosis Factor superfamily member that plays important neuronal functions, including apoptosis or synaptic remodeling. However, recent studies have shown that this receptor is also implicated in inflammatory processes via binding immunoglobulin superfamily member B7-1 [74]. In injured dorsal neural ganglia, P75^NTR^ signaling promotes M2 macrophage polarization [75].
VEGFA is an angiogenic growth factor that mediates the influence of M2 macrophages in the blood vessels of several cancers, contributing to tumor growth and metastasis [76]. In the interdigits, VEGFA maintains high levels of expression, with a moderate decline at 7.5 days, when the interdigital blood vessels begin to collapse [68].
HGF promotes cell proliferation and cell motility in regenerative processes. In the interdigits it is expressed at considerable levels during the remodeling process. As the abovementioned growth factors, it is produced by mesenchymal cells and tumor-associated macrophages [77], and their exogenous administration promotes an anti-inflammatory M2 macrophage phenotype [78].
Taken together, this pattern of expression of interdigital secreted signals is consistent with their involvement in maintaining the predominant M2 phenotype of interdigital macrophages.
Discussion
The formation of separated digits in vertebrates is mediated by massive cell death of the interdigital mesoderm of the embryonic limb. As in other areas of programmed cell death, macrophages accumulated massively in the remodeling interdigits to eliminate the remnants originating from the degenerative events [46–48, 79, 80]. Previous studies have shown that interdigital macrophages originate from the yolk sac at very early stages of development [18, 45, 47]. However, the time and manner in which macrophages reach the interdigits are less understood. At the functional level, besides its role in the elimination of cell remnants, it has been suggested that macrophages may trigger apoptosis in INZ [48]. Our study reveals a wide distribution of tissue resident macrophages in the whole limb primordia, preceding and independently of cell death. We show that the activated tissue resident macrophages are positive for Csf1r /NR staining/ and SA-β-Gal (pH6) histochemical labeling (Csf1r -NR- SA-β-Gal +), as well as other macrophage markers such as TAP1, F4/80 and TIM4. Furthermore, consistent with the routes between germ layers followed by migrating macrophages in early amphibian embryos [81], macrophages in the embryonic limb show a preferential, but not exclusive, subectodermal localization. The appearance of macrophages in micromass cultures obtained from healthy interdigital mesoderm confirms the presence of macrophages within the undifferentiated limb tissues preceding the dying process [48]. However, it is unlikely that macrophages were themselves the inducers of cell death, as occurs in the regression of hyaloid vessels in the developing mouse eye [40], because macrophages are widely distributed within limb tissues before, during, and after interdigit remodeling without necessarily being associated with death processes. Indeed, the attraction exerted by the interdigital mesoderm to recruit macrophages was strongly supported in our GFP+/GFP- chimeric experiments.
Q-PCR analysis and the in situ hybridization observations were consistent with the involvement of at least two waves of macrophages arriving at the interdigit during the degenerative process. Indeed, in contrast with the sustained expression of Csf1r, and Cx3cr1 genes prior to and throughout the degenerative events, Pu.1, a specific marker of hematopoietic macrophages, is almost unexpressed until 6.5 id, which corresponds to the onset of apoptosis. The up-regulation of this marker in the course of tissue remodeling supports that signals emanating from the dying process attract macrophages from hematopoietic regions of the embryo (i.e. aorto-gonadal-mesonephric regions or the fetal liver) that complement a primary population of resident macrophages recruited from the yolk sac.
Analysis of macrophage function via silencing Pu.1 [82] did not find modifications in digit development. This finding was claimed to support the idea that professional macrophages are not required for interdigit remodeling because skeletal progenitors can replace macrophages in the mutant mouse. We cannot discard this possibility, but the demonstrated diversity in the embryonic origin of adult macrophages [17] is suggestive of innate immune plasticity to replace missing macrophages with others from different embryonic locations [35, 83, 84]. However, this aspect awaits further clarification because a digit phenotype was neither reported in mice deficient for the Csf1r gene [7, 26].
In adult in vitro and in vivo conditions, macrophages exert dual contradictory effects on damaged tissues (reviewed by [64, 65]). First, macrophages, termed M1 macrophages (“Pro-inflammatory”, or “Classically activated”) release pro-inflammatory cytokines and chemokines to neutralize harmful agents. At a later stage, macrophages, termed M2 (“Anti-inflammatory”, or “Alternatively activated”), are functionally polarized to deliver reparative factors that promote trophic effects [3, 85–88]. There is consensus in considering the M1/M2 classification as an oversimplification with intermediate molecular macrophage profiles [64, 89] that can be actively modified in the course of the tissue damage by signals from the affected tissue. Alternative classifications (Classically Activated/Wound-Healing/ Regulatory macrophages) considering the functional diversity of M2 macrophages have been proposed [66]. The molecular signature we observed in the interdigits does not exactly match the M1/M2 division of macrophages. This can be due to a singular molecular signature of embryonic interdigital macrophages or, most likely, by the coincidence of distinct sub-populations of macrophages, as suggested by the expression pattern of Pu.1 discussed above. In general, M2 markers (Stat6, Stat3, Mrc1, Il4r) are expressed at higher levels than M1 markers, and some of them, such as Klf4, a zinc-finger transcription factor of major importance as an M2 promoter [90], are intensely up-regulated with the advance of tissue remodeling. However, the expression of some M1 markers (Tnfα, Il-16, Tlr4) intensifies in the late periods of interdigit remodeling. In the developing limb, interdigit remodeling begins with a massive apoptotic process that disintegrates the tissue, but, as occurs during wound healing processes, tissue must be restored almost simultaneously to confer the free or webbed digit morphology observed in different species. Together, our observations suggest that M2-like macrophages ensure the restoration of the interdigit undergoing degeneration. In turn, the increase in M1 markers could reflect the arrival of populations of M1-type macrophages attracted by debris secondary to necrotic degeneration of initially apoptotic cells [91].
Our transcriptional analysis of the remodeling interdigits confirms the expression of members of the so-called “Senescence-Associated Secretome Phenotype” (SASP), that has been considered as an indication of cellular senescence in embryonic systems [69, 92]. However, distinguishing between cell senescence and macrophage activation is often a difficult task in adult tissues. Indeed, whether macrophages share senescence features [70] or are senescent [93] is a controversial question. Our findings provide evidence against the interpretation of interdigital macrophages as senescent cells. First, because interdigital macrophages display intense phagocytic activity that contrasts with the decreased phagocytic capacity reported for senescent macrophages [94, 95]. Secondly, the SASP components detected here are shared by chronic inflammatory processes in adult organisms [96], playing a fundamental role in the regulation of macrophage polarization. Indeed, most of the factors overexpressed in the regressing interdigits have been described as promoters of M2 macrophage polarization in adult systems, including Csf-1 [60]; Tgfβ1 [71]; Igf1/Igfbp5 [73]; P75^NTR^ [75]; and Hgf [78].
From a developmental biology point of view, the complex origin [47] and widespread distribution of resident macrophages in embryonic tissue may reflect a dual strategy of development, with cells that construct embryonic organs according to precise growth and differentiation patterns, and cells characterized by high functional plasticity that eliminate the remnants derived from such a complex anabolic process at the same time favoring the remodeling and healing of degenerated tissues.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1. Supplementary table 1: Q-PCR specific primers for chick genes (DOCX 21 KB)
Supplementary material 2. Supplementary Figure 1: Graphical representation of ranges for Q-PCR CT values of macrophage markers. CT median value for Rpl13 in our experiments was 20,64±0,97. The dotted line indicates the CT values for genes that we consider to be unexpressed or expressed below levels of functional significance (DOCX 427 KB)
Supplementary material 3. Supplementary Figure 2: Graphical representation of the CT value ranges of qPCR transcripts of growth factor expression in interdigital samples at id 5.5, 6.5 and 7.5. The value for 7.5 is indicated with an asterisk. The dotted line indicates the CT values for genes that we consider to be unexpressed or expressed below levels of functional significance (DOCX 241 KB)