Molecular epidemiology of rodent-borne Leptospira spp. in Sri Lanka: identification of novel sequence types and previously unrecognized reservoir animals
Nipun Rathnayake, Kyosuke Takabe, Devinda Muthusinghe, Rydhnieya Vijeyakumaran, Pavani Senarathne, Nilanthi Dissanayake, Shuzo Urata, Kumiko Yoshimatsu, Yukihiro Akeda, Chandika Gamage, Nobuo Koizumi

TL;DR
This study explores the genetic diversity of Leptospira in rodents in Sri Lanka, identifying new strains and reservoir animals linked to human infections.
Contribution
The study identifies novel Leptospira sequence types and newly recognized rodent reservoirs in Sri Lanka.
Findings
Pathogenic Leptospira DNA was detected in 12.8% of rodent kidney samples from four species.
Five Leptospira sequence types were identified, including two novel types: ST389 and ST392.
Rattus rattus and Mus booduga were newly identified as sources of human Leptospira infections.
Abstract
Introduction. Leptospirosis is an important zoonotic disease globally, which is most prevalent in tropical regions. This disease is endemic in Sri Lanka, where the complex ecology of Leptospira spp., reservoir animals and environmental and occupational factors has resulted in a public health problem. Gap Statement. Although genomic analysis of Leptospira isolates has recently revealed the diversity of Leptospira spp. in Sri Lanka, the genetic relationship between human patients and reservoir animals remains unclear. Aim. This study investigated the genetic diversity of Leptospira spp. circulating in rodent populations in three districts of Sri Lanka: Kurunegala, Anuradhapura and Badulla. Methodology. Leptospira DNA was detected from rodent kidney tissue samples by real-time PCR, from which positive samples were subjected to flaB sequencing and multilocus sequencing typing (MLST).…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
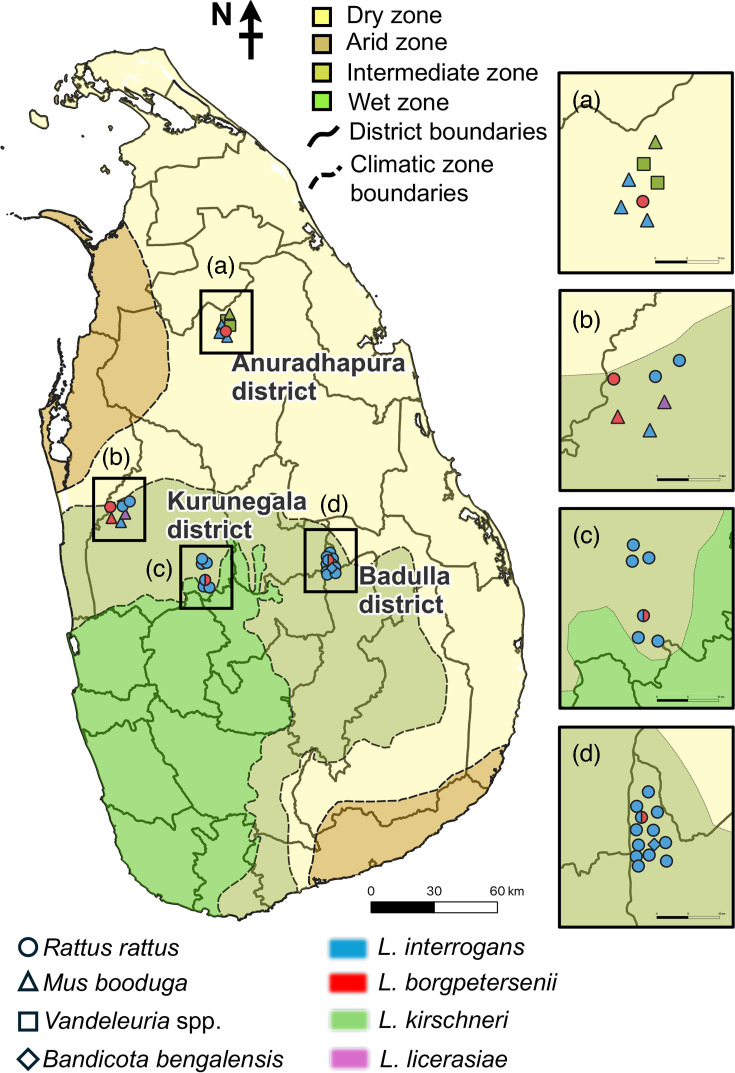 Fig. 1
Fig. 1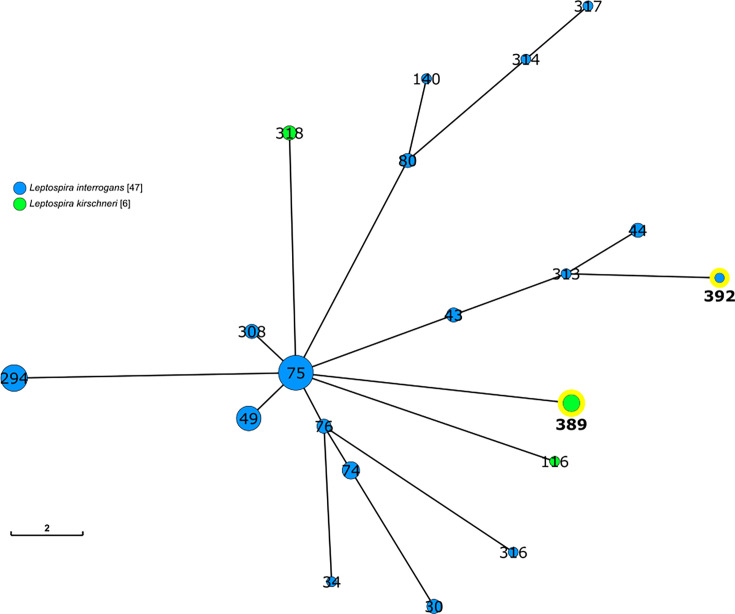 Fig. 2
Fig. 2| Sample ID | Real-time PCR | MLST | Rodent species | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
| P1 subclade | P2 subclade |
|
|
|
|
|
|
| ST | ||||
| UP-RK-026 | Kurunegala | P† | P |
| − | 5 | 1 | 1 | 1 | 3 | 2 | 7 | ST49 |
|
|
| UP-RK-038 | Kurunegala | −‡ | P | − | − | − | − | M | − | 10 | − | − |
|
| |
| UP-RK-047 | Kurunegala | − | P | − | − | M | − | 3 | − |
| − | − |
|
| |
| UP-RK-049 | Kurunegala | − | P | − | − | − | 3 | 3 | − | 9 | − | − |
|
| |
| UP-RK-061 | Kurunegala | P | P | − | − | 77 | 86 | M | 78 | M | 77 | 56 |
|
| |
| UP-RK-064 | Kurunegala | P | P |
| − | NT|| |
|
|
|
|
|
|
| ||
| UP-RK-068 | Kurunegala | − | P | − | − | 3 | − | − | − | − | 5 | − |
|
| |
| UP-RK-070 | Kurunegala | − | P | − |
|
|
|
|
|
|
|
|
| ||
| UP-RK-074 | Kurunegala | − | P | − | − | − | − | − | − | 9 | 5 | − |
|
| |
| UP-RK-075 | Kurunegala | P | P | − | − | 24 | 27 | 30 | 34 | 67 | 27 | 28 | ST144 |
|
|
| UP-RK-076 | Kurunegala | P | P | − | − | 24 | 27 | 30 | 34 | 67 | 27 | 28 | ST144 |
|
|
| UP-RK-077 | Kurunegala | P | − | − | − | − | − | − | − | 9 | − | − |
|
| |
| UP-RA-088 | Anuradhapura | − | P | − | − | − | − | 3 | − | 10 | 5 | − |
|
| |
| UP-RA-089 | Anuradhapura | − | P | − | − | − | − | − | − | M | − | − |
|
| |
| UP-RA-090 | Anuradhapura | P | P |
| − | 24 | 28 | 30 | 35 | 37 | 26 | 73 | ST323 |
|
|
| UP-RA-126 | Anuradhapura | P | P |
| − |
|
| 15 |
|
|
|
|
|
| |
| UP-RA-127 | Anuradhapura | P | P |
| − |
|
| 15 |
|
|
|
|
|
| |
| UP-RA-128 | Anuradhapura | P | P |
| − |
|
| 15 |
|
|
|
|
|
|
|
| UP-RA-130 | Anuradhapura | P | − | − | − | − | − | 3 | − | 9 | M | − |
|
| |
| UP-RB-156 | Badulla | P | − |
| − | − | − | − | − | − | 5 | − |
|
| |
| UP-RB-157 | Badulla | − | P | − | − | − | − | − | − | − | − | − |
| ||
| UP-RB-178 | Badulla | P | − | − | − | − | − | − | − | 68 | 2 | − |
|
| |
| UP-RB-181 | Badulla | − | P | − | − | − | 14 | − | − | 4 | 5 | − |
|
| |
| UP-RB-182 | Badulla | P | − |
| − | 8 | 14 | 2 | 17 | 10 | 5 | 6 |
|
|
|
| UP-RB-193 | Badulla | − | P | − | − | − | − | 2 | 17 | 26 | − | M |
|
| |
| UP-RB-204 | Badulla | P | P |
| − | − | − | − | 17 | − | 16 | 6 |
|
| |
| UP-RB-210 | Badulla | − | P | − | − | − | − | − | − | M | − | − |
|
| |
| UP-RB-212 | Badulla | P | P |
| − | 8 | 14 | 2 | 17 | M | M | 6 |
|
| |
| UP-RB-213 | Badulla | − | P | − | − | − | − | − | − | M | 5 | 8 |
|
| |
| UP-RB-224 | Badulla | P | P |
| − |
| − | 3 | 35 |
| 5 |
|
| ||
| UP-RB-225 | Badulla | P | P | − | − | − | − | − | − | 10 | − | − |
|
| |
| UP-RB-233 | Badulla | P | − |
| − | − | − | − | − |
| 4 | 7 |
|
| |
| UP-RB-240 | Badulla | P | P | − |
|
|
|
|
|
|
|
| |||
| Zone | District | Host species | No. of tested | No. of positive | % positive | Total positivity (no. of positive/no. of tested) |
|---|---|---|---|---|---|---|
| Dry | Anuradhapura |
| 1 | 0 | 0.0 | 11.5 (7/61) |
|
| 45 | 4 | 8.9 | |||
|
| 10 | 1 | 10.0 | |||
|
| 3 | 0 | 0.0 | |||
| 2 | 2 | 100.0 | ||||
| Intermediate | Kurunegala |
| 28 | 0 | 0.0 | 15.2 (12/79) |
|
| 5 | 3 | 60.0 | |||
|
| 45 | 9 | 20.0 | |||
|
| 1 | 0 | 0.0 | |||
| Badulla |
| 1 | 1 | 100.0 | 12.0 (14/117) | |
|
| 2 | 0 | 0.0 | |||
|
| 113 | 13 | 11.5 | |||
| 1 | 0 | 0.0 |
- —http://dx.doi.org/10.13039/501100004398 Mitsubishi Foundation
- —http://dx.doi.org/10.13039/501100003485 Heiwa Nakajima Foundation
- —http://dx.doi.org/10.13039/100009619 Japan Agency for Medical Research and Development
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLeptospirosis research and findings · Syphilis Diagnosis and Treatment · Leprosy Research and Treatment
Introduction
Leptospirosis is one of the most prevalent zoonoses and is caused by pathogenic spirochetes of the genus Leptospira. It is estimated that approximately one million leptospirosis cases and 58,900 associated deaths occur annually worldwide, of which more than 70% occur in the tropics [1]. However, leptospirosis is underdiagnosed in many tropical regions, mainly due to its diverse, nonspecific clinical manifestations that are similar to those of many other infectious diseases such as dengue fever, and limited or absent capacity of laboratories to diagnose it [25]. Pathogenic Leptospira spp. colonize the proximal renal tubules and are excreted in the urine of maintenance host animals [67]. Humans are infected with Leptospira spp. through damaged skin or mucous membranes mainly by exposure to water or soil contaminated with the urine of maintenance hosts [68]. Although rodents are important maintenance hosts of Leptospira spp., domestic animals, such as dogs, cattle and pigs, and wildlife species can also be reservoirs of certain Leptospira strains (serovars) [68]. Leptospira spp. comprise pathogenic and saprophytic species, which are divided into four subclades: P1 and P2 for pathogenic species and S1 and S2 for saprophytic ones [9]. In the pathogenic subclade, P1 contains pathogenic species responsible for human and animal infections. Although P2 subclade species are considered to have intermediate pathogenicity and are less virulent than P1 pathogens, human infection with P2 subclade Leptospira spp. has been reported in various locations globally [1015].
Sri Lanka is recognized as a hotspot for leptospirosis, with an estimated annual incidence of 52.1 cases per 100,000 population and ~730 deaths [16]. Leptospirosis cases have been reported in almost all regions of the country, with a high incidence in its wet zones [17]. Within the last decade, the highest number of clinically suspected cases, 9,630 cases and 203 deaths, was reported in 2023 [18], which was the highest incidence since the 2008 outbreak [1920]. This increase may be related to several factors, including an increase in agricultural activities due to the country’s economic crisis, increased rainfall, higher detection in advances in laboratory facilities and improved surveillance [21]. The prevalence of leptospirosis in Sri Lanka is influenced by environmental, weather-related, occupational and behavioural factors. Among these, agricultural exposure, particularly rice farming, is the most important [2223]. The widespread practice of rice farming involves flooded or muddy fields, especially during the rainy season, creating optimal conditions for Leptospira bacteria. Farmers working barefoot in these paddy fields are particularly vulnerable to exposure [22]. Leptospirosis is known as rat fever in this country [24], and Leptospira spp. have been detected in rodents and shrews, such as black rats (Rattus rattus), lesser bandicoot rats (Bandicota bengalensis), Indian bandicoot rats (Bandicota indica), little Indian field mice (Mus booduga) and Asian house shrews (Suncus murinus) [2526]. In addition to rodents, Leptospira spp. have been detected in companion and domestic animals such as dogs, cattle, buffaloes and elephants [25,2729].
Although many Leptospira isolates were obtained from humans, rodents, shrews and dogs in the 1960s and early 1970s [3031], recent studies have again started to successfully isolate Leptospira spp. in Sri Lanka [263233]. In 2020, multilocus sequencing typing (MLST) revealed 15 sequence types (STs), including 6 new STs from the isolates of 3 Leptospira species, Leptospira borgpetersenii, Leptospira interrogans and Leptospira kirschneri, from human patients and black rats in Sri Lanka [26] In 2022, six STs, including four new STs, from the above three species were identified in rodent populations [25]. Whole-genome sequencing (WGS) of 25 Leptospira isolates from human patients, including L. borgpetersenii, L. interrogans, L. kirschneri and Leptospira weilii, identified 15 clonal groups, including 12 groups that had not previously been reported [33]. A WGS study also showed that L. kirschneri isolates from febrile patients formed a distinct phylogenetic cluster from other L. kirschneri strains [34]. In addition to these four Leptospira species, Leptospira kmetyii and Leptospira licerasiae DNA has been detected in humans and other animals [3536]. The diversity of Leptospira spp. highlights the importance of environmental and animal reservoirs in disease transmission in Sri Lanka. However, although black rats and Asian house shrews are the source of human infection for L. borgpetersenii ST144 [263233], the reservoir animals for other Leptospira genotypes remain unknown.
In this study, we detected Leptospira DNA in rodent kidney tissue samples by real-time PCR targeting lipL32 and rrs in three districts of Sri Lanka: Kurunegala, Anuradhapura and Badulla. The real-time PCR-positive samples were subjected to MLST using seven housekeeping genes and flaB sequencing to identify Leptospira species and genotypes (STs).
Methods
Sample collection from the rodents
A total of 257 rodents were captured using live traps in paddy fields, agricultural areas and nearby houses from three districts of Sri Lanka, Kurunegala (n=79), Anuradhapura (n=61) and Badulla (n=117), between 2016 and 2024 (Fig. 1). The rodents were humanely euthanized using isoflurane inhalation, following the guidelines of the American Veterinary Medical Association, and kidney tissues were collected and stored at −25 °C prior to DNA extraction. Ethical approval for this study was obtained from the ethical review committee of the Faculty of Veterinary Medicine and Animal Sciences, University of Peradeniya (ethical approval no.: VERC-22-03).
A map of Sri Lanka showing the capture sites of Leptospira-positive rodents in relation to climatic zones. Geographical distribution of Leptospira-positive rodents captured in three districts of Sri Lanka plotted onto the country’s climatic zones. Colour shading indicates the climatic zones (dry, arid, intermediate and wet). District boundaries and climatic zone boundaries are shown by solid and dashed lines, respectively. Insets (a–d) present enlarged views of the specific trapping locations within each district. Symbols denote rodent species (R. rattus, M. booduga, Vandeleuria spp. and B. bengalensis), and colours indicate Leptospira species detected (L. interrogans, L. borgpetersenii, L. kirschneri and L. licerasiae).
Detection of Leptospira DNA from rodent kidney tissue
DNA was extracted from ~25 mg of rodent kidney tissue cortex using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany). The extracted DNA was then subjected to real-time PCR for lipL32 and rrs. For lipL32 detection, the sequences of the primers were the same as previously described [37], while the probe contained the ZEN quencher: 5′-FAM-AAAGCCAGG/ZEN/ACAAGCGCCG-IBFQ-3′, and the concentrations of the primers and the probe used were 0.3 and 0.2 µM, respectively. For rrs detection, 0.4 µM of the forward primer F3C (5′-TCATTGGGCGTAAAGGGTGC-3′), 0.6 µM of the reverse primer B3C (5′-TCAGTTTTAGGCCAGCAAGTC-3′) and 0.25 µM of the double-quenched probe DQ16SP (5′-FAM-AGAGGCAAG/ZEN/TGGAATTCCAGGTG-IBFQ-3′) were used. The real-time PCR was performed with Thunderbird Probe qPCR Mix (Toyobo, Osaka, Japan) on a LightCycler 96 (Roche, Basel, Switzerland): after an initial denaturation step at 95 °C for 60 s, the reaction mixture was subjected to 40 cycles of denaturation at 95 °C for 10 s and amplification at 60 °C for 60 s.
flaB-nested PCR
The real-time PCR-positive DNA samples were subjected to nested PCR targeting flaB for the P1 subclade Leptospira spp. [38] and the P2 subclade Leptospira spp. For P2 subclade Leptospira flaB amplification, the first PCR primer set L-IflaB-F1 (5′-CCGCTCTCTGAAGTTCAACGAG-3′) and L-IflaB-R1 (5′-GTTTGAGGACCGAATTCGGTTTT-3′) and the second PCR primer set L-IflaB-F2 (5′-GAGCWAGCTGTGGATAAAACC-3′) and L-IflaB-R2 (5′-CTAACATCGCCGTACCACTCTGCA-3′) were used. The reaction conditions were the same as previously described [38]. DNA sequencing of the amplicons was performed with the second primers using the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA). The nucleotide sequences of flaB detected were deposited in a public database (DDBJ accession numbers LC869567–LC869580).
MLST
MLST was applied to real-time PCR-positive DNA samples, and seven housekeeping genes, glmU, pntA, sucA, tpiA, pfkB, mreA and caiB, were amplified as previously described [3940]. DNA sequencing of the amplicons was performed using the M13 primers as described above. The nucleotide sequences of the above genes have been deposited in a public database (DDBJ; accession numbers LC869459–LC869566). New allele sequences have also been deposited in the Leptospira PubMLST database (https://pubmlst.org/organisms/leptospira-spp). A minimum spanning tree (MST) based on the allelic profiles of MLST of L. interrogans and L. kirschneri determined in this study and strains deposited in the MLST database was constructed using GrapeTree [41].
Identification of the rodent species
Rodent species were identified morphologically, and those from which leptospiral DNA was detected were further confirmed by sequencing of the mitochondrial cytochrome b (Cytb) gene. The Cytb gene was amplified using the primer set L14115 and H655A [42], and the amplicons were sequenced with the same primers. The resulting sequences were compared with those in public databases using the blast algorithm (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Statistical analysis
The associations between the Leptospira carriage rate in rodents and the districts where rodents were captured were tested using a 2×3 chi-square test. A P value less than 0.05 was considered statistically significant.
Results
Detection of Leptospira DNA from rodent kidney tissues
Leptospiral DNA was detected in 20 and 27 of the 257 kidney tissue samples by real-time PCR for lipL32 and rrs, respectively (Table 1). In total, 33 samples were positive by at least 1 assay. Both genes were detected in 14 samples, whereas lipL32 alone or rrs alone was detected in 6 and 13 samples, respectively. These results indicate that the rrs assay detected a greater number of positives than the lipL32 assay, suggesting higher analytical sensitivity in our rodent samples. Conversely, the six samples positive only by lipL32 indicate that each assay may miss a subset of infections. The rodent species from which leptospiral DNA was detected were R. rattus (23 animals), M. booduga [7], Vandeleuria sp*.* [2] and B. bengalensis [1] (Tables 1 and 2). There was no significant difference in the carriage rate of Leptospira spp. among the districts where rodents were captured: Kurunegala (15.2%, 12 out of 79), Anuradhapura (11.5%, 7 out of 61) and Badulla (11.9%, 14 out of 117) [χ^2^ (2, N=257)=0.57, P=0.75] (Table 2).
Species identification and MLST for Leptospira DNA detected in rodent kidney tissues
flaB of the P1 and P2 subclades was detected in 13 and 1 of the 33 real-time PCR-positive samples, respectively (Table 1). Of the 33 samples positive by real-time PCR, 3 (2 with mixed profiles and 1 identified as a P2 species by flaB sequencing) were not subjected to MLST. Among the remaining 30 samples, at least 1 MLST gene was amplified in 29. The highest positivity was observed for pfkB (26 out of 30), while pntA showed the lowest (12 out of 30) (Table 1). There were 7 alleles in glmU, 7 in pntA, 5 in sucA, 6 in tpiA, 12 in pfkB, 8 in mreA and 8 in caiB. One new allele was found in pntA, tpiA and mreA, two in glmU and caiB and three in pfkB (Table 1). Combined with the sequences of flaB and seven housekeeping genes, four Leptospira spp. were identified in Sri Lankan rodents. Specifically, L. interrogans, L. borgpetersenii, L. kirschneri and L. licerasiae were identified in 24, 4, 3 and 1 rodents, respectively. Mixed infections, defined by either (i) the presence of double peaks in sequencing chromatograms or (ii) the detection of alleles originating from different Leptospira species (e.g. L. interrogans and L. borgpetersenii) across the 7 MLST loci or flaB, were observed in 12 of the 33 rodents (36.4%).
All seven loci were successfully determined in eight animals, including L. borgpetersenii ST144 in R. rattus and M. booduga, L. borgpetersenii ST323 in R. rattus, L. interrogans ST49 in R. rattus, a novel ST (ST392) of L. interrogans in R. rattus and a novel ST (ST389) of L. kirschneri in Vandeleuria sp. and M. booduga (Table 1). The novel L. interrogans ST392 consisted of the allele combination glmU: 8, pntA: 14, sucA: 2, tpiA: 17, pfkB: 10, mreA: 5 and caiB: 6. Five of these alleles were identical to those of ST57. The novel L. kirschneri ST389 consisted of six novel alleles (glmU: 93, pntA: 102, tpiA: 94, pfkB: 130, mreA: 89 and caiB: 85) and one previously known allele (sucA: 15) (Table 1). The MST indicated that L. interrogans ST392 and L. kirschneri ST389 were genetically distinct from the other Sri Lankan isolates (Figs 2, S1 and S2, available in the online Supplementary Material). The flaB sequence of L. licerasiae UP-RK-070 was identical to that detected in human blood in Sri Lanka (accession number LC752688 [36].
MST of L. interrogans and L. kirschneri strains in Sri Lanka. Each circle represents an ST, and the numbers indicate ST designations. Circle sizes are proportional to the number of strains assigned to each ST. Blue and green circles represent L. interrogans and L. kirschneri, respectively, and yellow-highlighted circles indicate the new STs identified in this study. The MST was constructed based on allelic profiles from the seven housekeeping genes of MLST scheme 1.
Discussion
This study revealed that the kidney tissues of 12.8% (33 out of 257) of rodents in the Kurunegala, Anuradhapura and Badulla districts of Sri Lanka contained Leptospira spp. with high genetic diversity, including four Leptospira species and at least 12 genotypes, including 2 novel STs. Here, Leptospira spp. were detected in the black rat, R. rattus; the lesser bandicoot rat, B. bengalensis; and the little Indian field mouse, M. booduga, as reported previously [2526]. However, to the best of our knowledge, this is the first study to identify the long-tailed mouse, Vandeleuria sp., as a carrier of Leptospira. The carriage rate in rodents was almost the same among the three regions. The areas where rodents were trapped in the Badulla and Kurunegala districts belong to the intermediate zone, while the Anuradhapura district belongs to the dry zone (Fig. 1, Table 2). Although the prevalence of rodents carrying L. interrogans in Southeast Asia is known to be higher in humid habitats than in dry ones [43], another study in Sri Lanka reported no difference in the carriage rate between rodents captured in the dry and intermediate zones [2526], which may be due to the different genotypes of L. interrogans circulating in these areas.
MLST of Leptospira DNA in kidney tissue samples identified reservoir animals that are the source of infection for humans. L. borgpetersenii ST144 has been isolated from human patients, as well as black rats and shrews in Sri Lanka [263233], and this study identified the little Indian field mouse as the reservoir of this genotype in addition to black rat. L. interrogans ST49 has been isolated from humans in Sri Lanka (https://pubmlst.org/organisms/leptospira-spp), and this study demonstrated that black rats are the source of infection for this genotype in humans. In addition to the existing STs, this study also identified two novel STs: L. interrogans ST392 and L. kirschneri ST389 (Table 1, Figs 2, S1 and S2), although these STs have not been detected in human patients. The novel L. interrogans ST392 shared five of seven alleles with ST57, suggesting a close genetic relationship, whereas the new L. kirschneri ST389 comprised mostly novel alleles, representing a distinct lineage from previously identified Sri Lankan isolates (Table 1, Figs 2, S1 and S2). This suggests the presence of unique genetic variations in local L. kirschneri populations. Therefore, further studies are needed to investigate these novel genetic variations, particularly their role in zoonotic spread.
Human and animal infection with the P2 subclade of Leptospira spp. has been reported in various locations worldwide [1015,44]. In Sri Lanka, L. licerasiae DNA has been detected in humans [35], dogs [3536] and pigs (DDBJ accession number LC830693), and this study showed that the little Indian field mouse, M. booduga, is a potential source of L. licerasiae infection in humans. A wide variety of animals such as rats (Rattus spp. and Proechimys spp.), opossums (Metachirus nudicaudatus) and bats (Uroderma magnirostrum) are known to be reservoirs for L. licerasiae in the Peruvian Amazon [12], suggesting that other animals may be reservoirs for other genotypes of L. licerasiae in Sri Lanka. In addition, this study demonstrated the utility of real-time PCR for rrs, resulting in the identification of the mouse as a reservoir. It has been reported that some assays targeting rrs can detect DNA from other bacteria, especially when they are applied to urine samples [4547]. The assay used in this study showed high sensitivity and specificity that are comparable with those of the assay targeting lipL32 (Table 1), although no other genes were detected in 1 rrs-positive sample (Cq 36.57, the highest value in this study). Notably, the rrs assay identified more positive samples than lipL32, suggesting higher analytical sensitivity under our conditions. However, six samples were detected only by lipL32, indicating that both assays have inherent limitations. These discrepancies may be attributable to low DNA concentrations, stochastic amplification effects or sequence variation at primer and/or probe binding sites.
Mixed infections with the same or different species were observed in 36.4% (12 out of 33) of the animals from which Leptospira sequences were obtained. Mixed infections with L. borgpetersenii and L. interrogans in a single animal have been observed in different small mammals from around the world [4849]. This common phenomenon poses a problem for PCR-based MLST: in the sample UP-RB-224, the sequences of sucA and mreA were derived from L. interrogans, while that of tpiA was from L. borgpetersenii (Table 1), indicating that each locus can be amplified from different strains infecting a single animal and that it is not possible to determine whether the sequences are from the same or different strains when multiple strains of the same species are infecting a single animal. Therefore, the new ST of L. interrogans identified in this study needs to be confirmed in an isolate. Although sample-to-sample contamination cannot be entirely excluded, this is unlikely because double peaks were not consistently observed across all loci but rather appeared only in specific genes, and because negative controls consistently remained negative. Moreover, the presence of alleles corresponding to different species in the same specimen strongly suggests genuine co-infection rather than laboratory contamination.
There are several limitations to this study. First, we focused exclusively on rodents and did not include other potential reservoir animals that may contribute to human infections. Second, the novel STs of L. interrogans and L. kirschneri identified in this study have not yet been detected in human patients, and therefore, their zoonotic potential and transmission pathways remain to be determined. Finally, our conclusions regarding the reservoir role of rodents are based solely on the detection of leptospiral DNA in kidney tissues, and isolation of viable organisms from urine would be valuable to confirm active shedding, although such attempts are technically challenging because obtaining uncontaminated urine from small wild rodents is difficult.
In conclusion, this study demonstrates the genetic diversity of Leptospira spp. in rodent reservoirs in Sri Lanka, highlighting the complexity of Leptospira ecology and the disease transmission to humans. In addition, the high frequency of mixed infections highlights the diagnostic and clinical challenges associated with coinfections. There are still several genotypes detected only in human patients, such as ST323 in this and previous studies [2526], and continued surveillance of rodents and other reservoir animals and genetic analysis of Leptospira isolates are needed to better understand transmission pathways, develop effective strategies for controlling leptospirosis and improve public health interventions in Sri Lanka.
Supplementary material
10.1099/jmm.0.002133Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Costa F Hagan JE Calcagno J Kane M Torgerson P et al Global morbidity and mortality of leptospirosis: a systematic review P Lo S Negl Trop Dis 20159 e 000389810.1371/journal.pntd.000389826379143 PMC 4574773 · doi ↗ · pubmed ↗
- 2Guernier V Goarant C Benschop J Lau CL A systematic review of human and animal leptospirosis in the Pacific Islands reveals pathogen and reservoir diversity P Lo S Negl Trop Dis 201812 e 000650310.1371/journal.pntd.000650329758037 PMC 5967813 · doi ↗ · pubmed ↗
- 3Berlioz-Arthaud A Kiedrzynski T Singh N Yvon JF Roualen G et al Multicentre survey of incidence and public health impact of leptospirosis in the Western pacific Trans R Soc Trop Med Hyg 200710171472110.1016/j.trstmh.2007.03.01817442353 · doi ↗ · pubmed ↗
- 4Victoriano AFB Smythe LD Gloriani-Barzaga N Cavinta LL Kasai T et al Leptospirosis in the Asia Pacific region BMC Infect Dis 2009914710.1186/1471-2334-9-14719732423 PMC 2749047 · doi ↗ · pubmed ↗
- 5Musso D Roche C Marfel M Bel M Nilles EJ et al Improvement of leptospirosis surveillance in remote Pacific Islands using serum spotted on filter paper Int J Infect Dis 201420747610.1016/j.ijid.2013.11.00724384412 · doi ↗ · pubmed ↗
- 6Bharti AR Nally JE Ricaldi JN Matthias MA Diaz MM et al Leptospirosis: a zoonotic disease of global importance Lancet Infect Dis 2003375777110.1016/s 1473-3099(03)00830-214652202 · doi ↗ · pubmed ↗
- 7Faine S Adler B Bolin C Perolat P. Leptospira and Leptospirosis, 2nd ed Med Sci Press 1999
- 8Levett PN Leptospirosis: a forgotten zoonosis?Clinc Appl Immunol rev 2004443544810.1016/j.cair.2004.08.001 · doi ↗