Azole Resistance in Candida parapsilosis From Patients With Burns in Mexico: A Genomic and Phylogenetic Analysis
María de Lourdes García‐Hernández, Luis Ángel Nuñez‐García, Jossue Ortíz‐Álvarez, Marco Antonio Delaye‐Martínez, Marlen Flores‐Huacuja, María del Rocío Vazquez‐Olivares, Luis Fernando Espinosa‐Camacho, Gabriela Delgado‐Sapien, María del Rosario Morales‐Espinosa

TL;DR
This study analyzes Candida parapsilosis strains from burn patients in Mexico to understand azole resistance patterns and genetic variants.
Contribution
The study identifies specific genetic mutations linked to azole resistance in C. parapsilosis from clinical isolates in Mexico.
Findings
Most isolates were Candida parapsilosis sensu stricto, with 33% showing fluconazole resistance.
The ERG11 Y132F variant was common in resistant isolates.
Two haplotypes were identified through SNP and phylogenetic analysis.
Abstract
Candida species are common causes of nosocomial infections. Candida parapsilosis has emerged as an important pathogen. Mortality rates associated with invasive C. parapsilosis infections range from 14.5% to 47%, with azole resistance estimated between 4% and 15%. Resistance mechanisms include mutations in ERG11 that diminish azole binding, upregulation of ergosterol biosynthesis genes due to mutations in regulatory factors like UPC2 and NDT80, and increased efflux pump activity. Thirty clinical strains of the C. parapsilosis complex were isolated from bloodstream cultures, biopsies and catheter tips from hospitalised patients from a major rehabilitation centre in Mexico between June 2011 and August 2016. Minimum inhibitory concentrations for fluconazole, voriconazole, itraconazole, caspofungin, micafungin, anidulafungin and amphotericin B were determined using the broth…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
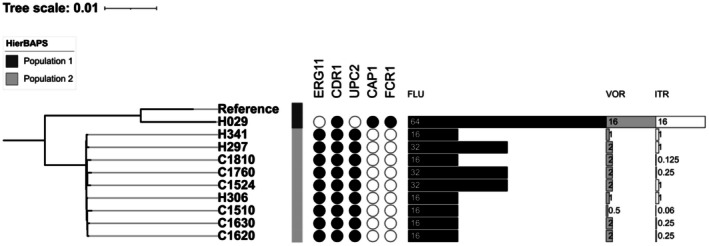 FIGURE 1
FIGURE 1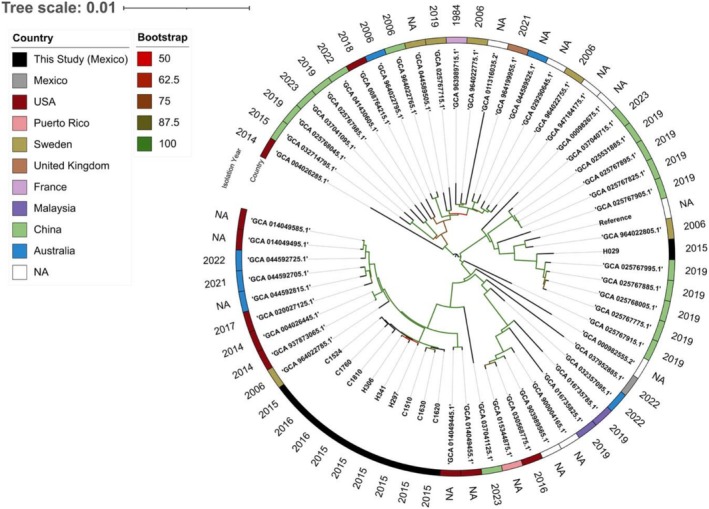 FIGURE 2
FIGURE 2| Clinical characteristics | Overall ( |
|---|---|
| Sex (male) | 23 (82) |
| Age (years) | 36 (20–54) |
| Comorbidities | |
| Diabetes mellitus | 2 (7.1) |
| Hypertension | 5 (18) |
| Obesity | 5 (18) |
| Burn injury mechanism | |
| Fire | 21 (75) |
| Scald | 1 (3.6) |
| Electric | 4 (14) |
| Mixed: fire + electricity | 2 (7.1) |
| Burn severity | |
| 2nd degree | 7 (25) |
| 3rd degree | 21 (75) |
| Burn Total Surface Body Area | 45 (30–58) |
| ABSI score | 8 (6–10) |
| Features | Overall ( |
|---|---|
| Yeast‐associated infection | |
| Invasive bloodstream infection | 12 (43%) |
| Catheter tip infection | 4 (14%) |
| Soft tissue infection | 12 (43%) |
| Anti‐fungal therapy | |
| FLU | 19 (68%) |
| VOR | 1 (3.6%) |
| AND | 3 (11%) |
| CAS | 3 (7.1%) |
| AMB | 2 (7.1%) |
| Bacterial coinfection | 15 (54%) |
| Treatment duration (days) | 14 (7–19) |
| Length of stay (LOS) (days) | 63 (34–83) |
| Overall mortality | 4 (14%) |
| Antifungal | Minimum inhibitory concentrations (MIC) (μg/mL) | Categorical susceptibility | Epidemiological definition | |||||
|---|---|---|---|---|---|---|---|---|
| Range (μg/mL) | MIC50 | MIC90 (μg/mL) | S | SDD/I | R | WT | nWT | |
| FLU | 0.03–64 | 1 | 32 | 19 (63.3) | 1 (3.3) | 10 (33.3) | — | — |
| VOR | 0.03–16 | 0.03 | 2 | 20 (66.6) | 1 (3.3) | 9 (30) | — | — |
| ITR | 0.03–16 | 0.25 | 1 | — | — | — | 25 (83.3) | 5 (16.6) |
| CAS | 0.125–2 | 1 | 2 | 30 (100) | 0 | 0 | — | — |
| MCF | 0.125–4 | 1 | 2 | 29 (96.6) | 1 (3.3) | 0 | — | — |
| AND | 0.03–2 | 1 | 1 | 30 (100) | 0 | 0 | — | — |
| AMB | 0.5–2 | 1 | 2 | — | — | — | 20 (66.6) | 10 (33.3) |
| Strain | Clinical specimen | Species | MIC (μg/mL) | Genotype | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FLU | VOR | ITR |
|
|
|
|
| |||
| H029 | Blood |
|
|
|
| — | WT/I1287V | — | WT/C478* | Q211*/Q211* |
| H297 | Blood |
|
|
|
| WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| H306 | Blood |
|
|
|
| WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| H341 | Blood |
|
|
|
| WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| C1510 | Biopsy |
|
| 0.5 | 0.06 | WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| C1524 | Biopsy |
|
|
|
| WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| C1620 | Biopsy |
|
|
| 0.25 | WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| C1630 | Biopsy |
|
|
| 0.25 | WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| C1760 | Biopsy |
|
|
| 0.25 | WT/Y132F | I1287V/I1287V | WT/N455D | — | — |
| C1810 | Biopsy |
|
|
| 0.125 | Y132F/Y132F | I1287V/I1287V | WT/N455D | — | — |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Nail Diseases and Treatments · Skin and Cellular Biology Research
Introduction
1
Candida species and yeast‐like (taxonomically reclassified into new genera and previously belonging to the genus Candida) are common causes of nosocomial infections. Candida albicans has been recognised as the most frequent species associated with infections; however, a shift towards higher frequency for non‐albicans species has been documented [1]. Candida parapsilosis has emerged as a predominant species in various settings, becoming one of the most identified non‐albicans species [2]. This yeast is a component of the human skin microbiota and can function as an opportunistic pathogen in immunocompromised individuals [1]. C. parapsilosis is part of a complex integrated by three cryptic species: C. orthopsilosis, C. metapsilosis and C. parapsilosis sensu stricto [3].
Mortality rates associated with invasive infections by C. parapsilosis range from 14.5% to 47% globally [2] and azole resistance within the complex is estimated between 4% [4] to 15% [5]. However, data on azole resistance in Mexico remain scarce. One study that included clinical isolates collected from 1999 to 2010 reported fluconazole resistance of 4.5% in C. parapsilosis sensu stricto isolates, with all C. orthopsilosis and C. metapsilosis isolates remaining susceptible [6]. In contrast, another study based on isolates collected from 2014 to 2016 documented fluconazole resistance of 54% in C. parapsilosis isolates [7].
Given the increasing concern regarding antifungal resistance and the limited availability of local surveillance data, this study aimed to identify mutations associated with azole resistance in isolates belonging to C. parapsilosis complex obtained from patients with burns treated at a referral hospital in Mexico City. Additionally, the study sought to contribute to the understanding of the molecular mechanisms underlying resistance in this clinically significant fungal species and to provide baseline information to support effective infection control and treatment strategies.
Material and Methods
2
Study Site, Culture Collection, Clinical Data and Statistical Analysis
2.1
This study was carried out at the Instituto Nacional de Rehabilitación Luis Guillermo Ibarra Ibarra (INR‐LGII), a centre specialising in rehabilitation and burn care.
For this study, 30 clinical strains of C. parapsilosis complex recovered from bloodstream cultures, biopsies and catheter tips from hospitalised patients from June 2011 to August 2016 were included. Isolates were identified with Vitek 2 compact (Biomérieux. Marcy‐l'Étoile, France) and were kept at −70°C in 20% glycerol Brain Heart Infusion.
For this study, clinical isolates were thawed, and identification was confirmed by Vitek MS (bioMérieux, Marcy‐l'Étoile, France). Clinical information from patients was retrieved from medical electronic records. Descriptive analysis using frequency measures and percentages was employed for categorical variables; median and interquartile range (IQR) were used for continuous variables.
This study was reviewed and approved by the local research committee under approval number 16/23.
Susceptibility Testing
2.2
Susceptibility to fluconazole (FLU), voriconazole (VOR), itraconazole (ITC), caspofungin (CAS), micafungin (MCF), anidulafungin (AND) and amphotericin B (AMB) was evaluated with the broth microdilution method and interpreted according to recommendations of CLSI M27‐A3 [8] to determine the minimum inhibitory concentration (MIC) for each antifungal; in addition, MIC_50_ and MIC_90_ were obtained, break points were considered according to CLSI M27M44S [9] and M57S [10]. For ITR and AMB, strains were defined as wild‐type (WT) and non‐wild‐type (nWT) categories according to the CLSI M57S guideline. C. parapsilosis ATCC 22019 and Pichia kudriavzevii (formerly Candida krusei ) ATCC 6528 were used as controls.
Whole Genome Sequencing
2.3
Isolates that showed resistance to at least one azole were selected for whole genome sequencing, with the aim of identifying the possible resistance mechanisms. Strains were grown on SDA and incubated at 30°C for 24 h. DNA extraction was performed using the Wizard Genomic DNA Purification Kit (Promega, Madison, Wisconsin, USA). Colonies were resuspended in 700 μL of Nucleic Lysis Solution and adjusted to a turbidity of 2 in the McFarland Scale. 10 μL of Proteinase K (10 mg/mL) and Lysozyme (10 mg/mL) was added and incubated at 37°C for 30 min; instructions from the manufacturer were followed after digestion. DNA quality was evaluated with NanoDrop One (Thermo Fisher Scientific, Waltham, Massachusetts, USA) and Qubit 4.0 fluorometer (Invitrogen, Waltham, Massachusetts, USA). Library preparation was performed with the Illumina DNA Prep Kit (Illuminam, San Diego, California, USA) and consisted of a tagging step, posttagmentation cleanup, library amplification and indexing IDT for Illumina DNA/RNA UD Indexes (Illumina, San Diego, California, USA). The libraries were then purified by a washing step with fresh 80% ethanol and then resuspended with the RSB buffer included in the kit. Subsequently, library quality assessment was performed with the 4200 TapeStation System (Agilent, Santa Clara, California, USA) and the Qubit 4.0 fluorometer (Invitrogen, Waltham, Massachusetts, USA). The libraries were normalised to 4 nM and two pools were made. The first pool was sequenced using a NextSeq High Output Flow Cell V2.5 and an NSQ 500Hi‐Output Reagment V2 cartridge (300 cycles) on the Illumina NextSeq 550 platform. The second pool was sequenced using a Flow cell V2 and a V2 cartridge (300 cycles) on the Illumina MiSeq platform, both in a paired‐end configuration. The samples were sequenced in the Sequencing Laboratory of the Centro Nacional de Referencia de Inocuidad y Bioseguridad Agroalimentaria (SENASICA).
Genome Assembly and Annotation
2.4
Raw sequencing reads were processed using Trim_Galore v0.6.10 [11] to remove low‐quality bases and adapters. De novo assembly was performed using three strategies: standalone SPAdes v4.2.0, Shovill‐SPAdes and Shovill‐SKESA [12, 13]. Metrics for assessing assembly quality were determined with QUAST [14]; the assembly with the fewest content of contigs was selected for further processing. Genome completeness and contamination were evaluated using BUSCO [15] with the Saccharomycetes dataset. Mitochondrial DNA contigs were manually removed using Seqkit [16].
Antifungal Resistance Variants Screening
2.5
Trimmed reads were aligned to the C. parapsilosis CDC317 reference genome (NCBI accession GCF_000182765.1) using BWA‐MEM [17]. SAM‐formatted alignments were converted to BAM format using Samtools v1.20 [18]. Variants were identified using BCFtools v1.20: a pileup was generated, followed by variant calling with the BCFtools ‘call’ function [18]. Variant annotation was performed using SnpEff v5.2 [19]. Sequences for genes with reported mutations associated with antifungal resistance (Table S1) were extracted from the Candida genome database: CAP1, CDR1, CPH1, ERG11, FCR1, MCM1, MDR1, MRR1, NDT80, REP1, STB5, TAC1 and UPC1 [20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30]. Annotations (GFF format) for the C. parapsilosis CDC317 reference genome were obtained from NCBI [31], and gene features corresponding to resistance‐associated loci were extracted. We compared the annotated variant file to these extracted properties using BEDTools v2.31.1 [32] ‘intersect’ function to identify variants overlapping these features.
SNP‐Distance, Average Nucleotide Identity and Phylogenetic Analysis
2.6
Taxonomy was verified by calculating Average Nucleotide Identity (ANI) using ANIclustermap [33] between isolates and reference genomes of Candida species and closely related yeasts. ANI values > 95% were considered as indicative of the same species [34]. For phylogenetic analysis, assemblies were mapped to the C. parapsilosis CDC317 reference genome using Snippy [35]. The whole‐genome alignment was processed with Genealogies Unibased By recomBinations in Nucleotide Sequences (Gubbins) [36] to mask recombination sites and obtain the core‐genome SNPs alignment; SNP‐distances were calculated using snp‐dists [37]. The core alignment was used to construct a maximum‐likelihood (ML) phylogenetic tree with IQ‐TREE2 [38], the best‐fit substitution model was determined by the ModelFinder module, and branch support was estimated from 1000 ultrafast bootstrap replicates.
Comparison With Genomic Public Records
2.7
C. parapsilosis sensu stricto genome assemblies were retrieved from the NCBI database. Duplicate assemblies of the CDC317 were removed. Both the de novo assemblies and the extracted genomes were analysed using the strategy for phylogenetic analysis described above. Additionally, the final tree and SNP‐distances were used to determine population structure using rhierbaps [36].
Data Availability Statement
2.8
Raw sequencing reads and assembled genomes were deposited in the National Center for Biotechnology Information (NCBI) under project number PRJNA1287628.
Results
3
Identification and Clinical Features of Patients
3.1
Clinical strains were recovered from 28 patients with burns. Most strains were isolated from bloodstream cultures (13/30; 43.3%), followed by biopsies (12/30, 40%) and catheter tips (5/30, 16.7%). Twenty‐nine strains were identified as * Candida parapsilosis sensu stricto* (96.7%) and one as Candida orthopsilosis (3.3%).
Most patients were male (82%) with a median age of 36 years. The extent of burns involved approximately 45% (30–58) of the Total Surface Body Area (TSBA). Almost 75% (22/28) exhibited third‐degree burns (Table 1).
Bacterial coinfection was present in 54% of the patients, with Pseudomonas aeruginosa (25%; 7/28) being the most frequent, followed by Acinetobacter baumannii (21.42%; 6/28). Three patients showed coinfection with other yeast species; one with C. albicans , one with C. tropicalis and one with Pichia kudriavzevii and C. albicans.
Antimicrobial treatment was adjusted to cover both invasive candidiasis and bacterial infections. With respect to the antifungal therapy, FLU and VOR were the main prescribed antifungals, followed by echinocandins (AND, CAS) or AMB, with a median duration of 14 days (IQR 7–19) (Table 2). In 10 (33.3%) patients, azole resistance was detected. These patients received FLU (n = 6), AND (n = 3) and AMB (n = 1). Clinical evolution was favourable in all patients, with the exception of one patient, who died 6 days after invasive candidiasis was detected. This patient received FLU treatment and had a burn TSBA of 50%, with concomitant coinfection in soft tissues with P. aeruginosa and A. baumannii. On the other hand, in the azole‐susceptible C. parapsilosis group, three patients died during the treatment period. All of these patients had burn injuries between 50% and 60% of TSBA, with polymicrobial infections both in the burn injuries and bloodstream. No adverse events were reported during the treatment duration. These results are shown in Tables [Link], [Link].
Susceptibility to Azoles in Clinical Strains of
C. parapsilosis Complex Isolated From Burned Patients
3.2
Approximately 30% of isolates were resistant to FLU (MIC 16–64 μg/mL) and VOR (MIC 1–16 μg/mL); meanwhile, six strains (16.6%) were nWT to ITR, and 10/30 isolates were nWT for AMB (Table 3). All isolates were susceptible to the antifungals CAS, MCF and AND. The MIC_90_ value for the antifungals ITR and AND is 1 μg/mL; for VOR, CAS, MCF and AMB, it is 2 μg/mL; and for FLU, it is 32 μg/mL (Table 3).
Phylogenetic Relationships and Antifungal Resistance‐Associated Mutations
3.3
Sequenced isolates were identified as part of the C. parapsilosis complex since all genomes lie within the 95% ANI species threshold (Figure S1). Phylogenomic analysis showed clustering of 9/10 sequenced isolates in this study, which formed a monophyletic clade (Figure 2). The closest related genomes from Genbank were isolates collected in the United States between 2014 and 2017 and in Australia during 2021 and 2022. These isolates clustered closely together, with fewer than 200 SNP differences among them (Figure S2). On the other hand, isolate H029 was phylogenetically distant from the other sequenced isolates, showing approximately 2400 SNPs differences and clustered with strains predominantly isolated in China, mostly from 2019, suggesting it represents a distinct lineage.
Strain H029 had a unique mutational profile characterised by variants in CDR1, CAP1 and FCR1 (Figure 1). Conversely, the primary clade shared mutations in ERG11, CDR1 and UPC2. The genotype 1: ERG11 ^WT/Y132F^, CDR1 ^I1287V/I1287V^ and UPC2 ^WT/N455D^, was present in 8/10 isolates. A closely related genotype 2: ERG11 ^Y132F/Y132F^, CDR1 ^I1287V/I1287V^ and UPC2 ^WT/N455D^, differed by carrying a homozygous Y132F mutation in ERG11, and occurred in isolate C1810 (Table 4). Missense variants in these genotypes have a putative moderate impact according to SnpEff annotation. The more divergent genotype 3: CDR1 ^WT/I1287V^, CAP1 ^WT/C478*^ and FCR1 ^Q211*/Q211*^ was identified in isolate H029. The nonsense mutations in CAP1 and FCR1 are predicted to have a putative high impact according to SnpEff. All the identified variants have a sequencing depth > 77, ensuring confidence in the variant calling.
Midpoint‐rooted maximum likelihood phylogenetic tree based on SNP distances of the sequenced isolates. Genes associated with antifungal‐resistance are shown on the right binary data representation. Bar‐chart values depict minimum inhibitory concentrations for fluconazole (FLU), voriconazole (VOR) and itraconazole (ITR). Numbers in bars represent values for minimum inhibitory concentrations.
All strains with genotype 1 exhibited resistance to FLU; seven were resistant and one SDD to VOR For ITR, four showed a WT phenotype. The strain with genotype 2 (C1810) showed similar MICs, resistance to FLU and VOR and WT for ITR. In contrast, strain H029 (genotype 3) was resistant to all azoles, with the highest MICs evaluated (Table 4).
Phylogenetic Relationship With Public Genomes
3.4
A total of 53 unique assemblies were obtained. A strong phylogenetic relationship was observed among 9/10 sequenced isolates in this study, which clustered together within a distinct clade (Figure 2). The closest related genomes in the NCBI Assembly database were isolates collected in the United States between 2014 and 2017 and in Australia during 2021 and 2022; all were classified as belonging to population one according to hierBAPS analysis. The remaining isolate, H029, was phylogenetically distant from the others and clustered with strains predominantly isolated in China, mostly from 2019.
Midpoint‐rooted maximum likelihood phylogenetic tree based on SNP distances. Publicly available Candida parapsilosis genomes from the NCBI RefSeq database (n = 55) and the sequence isolates were compared. ‘N/A’ indicates unavailable metadata. The outermost annotation layer represents the year of isolation.
To the best of our knowledge, this is the first study that described whole genome sequencing data from C. parapsilosis sensu stricto from strains isolated within a clinical context in Mexico.
Discussion
4
Patients with burns are highly susceptible to nosocomial infections due to the compromise of primary mucosal and skin barriers, and yeast plays a significant role, resulting in approximately 180,000 deaths each year [39]. Thus, in this study, we aimed to identify mutations associated with azole resistance in C. parapsilosis isolates obtained from burn patients treated at a specialised hospital in Mexico.
Invasive fungal infections in burned patients are predominantly caused by Candida species, particularly C. albicans and C. parapsilosis [40, 41, 42]. Within the C. parapsilosis complex, C. parapsilosis sensu stricto is the most reported species, with prevalence rates ranging from 88.5% to 95.3% [43, 44]. In contrast, C. orthopsilosis and C. metapsilosis account for 3.1%–8.4% and 1.6%–4.2% of isolates respectively [6, 45, 46]. Consistent with these reports, C. parapsilosis sensu stricto represented 96.7% of the isolates obtained from patients with burns in our study.
Invasive candidiasis has a global incidence rate reported in 12.1 cases/100,000 population, and is associated with mortality rates between 30% and 90% [43, 44, 47].
A recent ICU‐based study from Iran (2021–2023), including 353 burn patients reported yeast bloodstream infection in 15.3% of cases. In that cohort, mortality was significantly associated with advanced age, broad‐spectrum antibiotic exposure, yeast bloodstream infection, extensive TBSA involvement (> 50%) and prolonged LOS [48]. In our study, these same risk factors were prevalent: nearly half of the patients received broad‐spectrum antibiotics, 43% developed bloodstream infection, 45% had extended TSBA involvement and the median LOS was 63 days. In the same study, resistance to fluconazole and voriconazole was reported in 28% and 18.2% of isolates respectively [48].
Although azole resistance in invasive candidiasis has been associated with higher mortality, its relation with antifungal treatment failure in the burn patient might be difficult to elucidate. In this setting, mortality is often driven by the severity of the burn injury itself and by concurrent infections, most of them with multidrug‐resistant profiles. In our study, overall mortality was 14.3%, and all deaths occurred in patients with > 50% TBSA as well as concomitant bacterial bloodstream infection. Only one fatal case involving an azole‐resistant isolate was observed, and death was difficult to attribute to the azole's resistance.
Worldwide, azole resistance in C. parapsilosis is reported to be below 15% [4, 5]. In contrast, our study detected resistance percentages of 32.3% for FLU, 29% for VOR and 16.1% for ITR. When compared to two Mexican studies, our findings show intermediate resistance levels. Treviño et al. reported a much lower fluconazole resistance rate of 4.5%, although 56.6% of their isolates were obtained from blood cultures and the clinical origin of the remaining isolates was not specified [6]. Conversely, our resistance rates were lower than those reported by Corzo et al. who found a fluconazole resistance rate of 52% in blood‐derived clinical isolates [7]. Similar to our study, both previous reports were based on single‐centre samples, overlapping with the time frame of Corzo et al. (2011–2016) [7].
Atalay et al. identified intra‐abdominal surgery, hypoalbuminaemia and previous use of FLU as risk factors for developing azole‐resistant bloodstream infection by C. parapsilosis [49]. Similarly, Semet et al. reported the presence of a central venous catheter, colostomy and age over 66 years as risk factors [50]. In our population, 43% of patients developed catheter‐associated bloodstream infection, and 68% were treated with FLU, factors that may have contributed to the emergence of azole‐resistant infections in our cohort. However, detailed data on topical antifungal or antibacterial treatments, including wound irrigation protocols, were unavailable. Consequently, potential associations between local treatments and azole resistance could not be formally assessed.
Studies such as those by Rodriguez et al. [51] have demonstrated significant genetic diversity among clinical isolates. The work is focused on the amplification of eight polymorphic microsatellite regions by PCR [51, 52]. However, other works have reported a limited variability at the DNA level. For instance, Tavanti et al. used Amplification Fragment Length Polymorphism (AFLP) analysis on 62 clinical strains from different geographic regions and found reduced genetic diversity [53]. This finding is consistent with the work of Dr. Valach, who observed low genetic variability in C. parapsilosis compared to C. metapsilosis and C. orthopsilosis based on mitochondrial genome analysis [54]. These studies support the hypothesis that C. parapsilosis may have a recent evolutionary origin. This context is relevant to our findings, in which most of the sequenced azole‐resistant strains (9/10) belonged to the same population and centre. In burn patients specifically, resistance rates to FLU and VOR were 28% and 18.2% respectively. For C. parapsilosis , resistance rates are reported at 20% for FLU and 8% for VOR [48].
The ERG11 Y132F mutation is associated with reduced susceptibility to azole antifungals across Candida species [7, 55, 56]. In this study, nine out of 10 isolates harboured at least one allele carrying this mutation. No clear correlation was observed between ERG11 Y132F zygosity and azole resistance phenotypes, as isolate C1810 carries the ERG11 ^Y132F/Y132F^ genotype but exhibits lower MICs when compared to strains heterozygous for this mutation. These findings suggest that additional factors not explored in this study, such as polymorphisms in other transcriptional factor genes, differences in virulence factor content (particularly biofilm formation) [57], epigenetic regulation or copy number variations and complex chromosomal rearrangements [26], may contribute to the observed resistance patterns.
Mutations were also detected in CDR1 and UPC2, both genes known to be associated with antifungal resistance [58]. Two studies reported the I1287V mutation in CDR1 [59, 60]; however, only one has linked this mutation to increased MICs for antifungals [59]. Different studies have analysed the relevance of mutations in UPC2 for the overexpression of ergosterol biosynthesis genes [61, 62, 63], which contributes to azole nonsusceptibility [58]. Although numerous UPC2 mutations have been described [64, 65, 66], the functional impact of the N455D substitution identified in this study has not been elucidated. Notably, this substitution is located within the zinc‐finger domain required for transcription factor activity, which may suggest a potential effect on the regulation of gene expression [67].
Mutations with a high predicted impact on protein function were identified in strain H029, which also exhibited the highest MICs across all tested antifungal agents and notably lacked the common ERG11 Y132F mutation. Loss‐of‐function mutations in FCR1, a negative regulator of the CDR1 efflux pump, might be associated with FLU resistance in C. albicans [25, 68]. Isolate H029 harboured a homozygous loss‐of‐function mutation in this gene, suggesting a constitutive upregulation of CDR1 and leading to azole resistance. In contrast, CAP1, a positive regulator of the MDR1 efflux pump [20], carried a heterozygous loss‐of‐function mutation, which could suggest reduced MDR1 expression in this isolate, partially contradicting the hypothesis of efflux‐mediated resistance. However, CAP1 is partially haplosufficient [69], at least in promoting virulence gene expression, which might suggest that the probable effect on MDR1 under expression is less pronounced.
Phylogenetic and population structure analysis suggest that nine of the 10 sequenced isolates form a distinct lineage within our dataset, a finding supported by their convergent genotypes in antifungal resistance‐associated genes. These isolates were collected between 2015 and 2016, which, along with the low pairwise SNP‐distance, suggests clonal persistence during the observation period. In contrast, strain H029 was identified as a genotypically and phylogenetically distinct lineage with more than 2400 SNPs relative to all other sequenced isolates in this study. This divergence is further supported by the geographical clustering of strain H029 with isolates originating from China, whereas the remaining isolates clustered with strains from the United States and Australia. However, the temporal gap between H029 and the closest publicly available genomes, together with the absence of patient travel or referral history, limits inferences regarding its geographic origin or transmission route.
This study has limitations. First, it was conducted at a single centre, and therefore, the results do not represent the broader epidemiological landscape in Mexico. Second, the study population consisted exclusively of patients with burns, which may limit the generalisability of the findings to other types of infections or patient groups.
Third, the interpretation of the identified variants relied on previously reported data, and no functional validation or additional analyses were performed. Furthermore, only azole‐resistant isolates were sequenced; as a consequence, the generated data do not allow us to assess epidemiological factors that drive azole resistance. Consequently, the clinical and biological significance of the genotype observed in strain H029 remains speculative.
Conclusions
5
In this study, C. parapsilosis sensu stricto was the predominant species isolated from patients with burns at a specialised rehabilitation in Mexico, with a notable proportion exhibiting resistance to FLU and VOR. The Y132F variant in the ERG11 gene was present in most sequenced isolates, which clustered phylogenetically with strains previously reported in the United States and Mexico.
Despite sharing lineage and genotypes, isolates with ERG11 mutation expressed variable resistance to the antifungals that belong to the azoles, suggesting the contribution of other molecular mechanisms not explored in this study.
One phylogenetically distinct isolate, related to strains from China, exhibited elevated azole MICs in the absence of known resistance‐associated mutations, further supporting the need for comprehensive molecular analyses to elucidate alternative pathways of antifungal resistance. The narrowing of the phylogenetic spectrum of C. parapsilosis cannot be used to exclude the possibility of clonal distribution or the occurrence of outbreaks within our institution.
Author Contributions
María de Lourdes García‐Hernández and Luis Ángel Nuñez‐García: conceptualisation, methodology, formal analysis, investigation, writing, original draft, writing, review and editing. Jossue Ortíz‐Álvarez and Marco Antonio Delaye‐Martínez: formal analysis, data curation, writing, original draft, writing, review and editing. Marlen Flores‐Huacuja, María del Rocío Vazquez‐Olivares, Luis Fernando Espinosa‐Camacho, Gabriela Delgado‐Sapien, María del Rosario Morales‐Espinosa, Tatiana Chávez‐Heres and Noé Becerra‐Lobato: formal analysis, data curation. Rafael Franco‐Cendejas and Elvira Garza‐González: formal analysis, writing, original draft, writing, review and editing. Luis Ostrosky‐Zeichner: original draft, writing, review and editing. Tatiana Chávez‐Heres: formal analysis. Luis Esaú López‐Jácome: conceptualisation, validation, supervision, project administration, investigation, writing, original draft, writing, review and editing. Claudia Adriana Colín‐Castro: conceptualisation, validation, investigation, writing, original draft, writing, review and editing.
Funding
The authors have nothing to report.
Conflicts of Interest
Pertinent to this work, LO has received research grants and/or consulting honoraria from the following companies: Scynexis, Melinta, GSK, Pulmocide, F2G, Basilea, Pfizer, Gilead, T2 Biosystems, Octapharma, Meiji, Stendhal, Knight and Eurofins Viracor. He is partially funded by the National Center for Advancing Translational Sciences (NCATS), National Institutes of Health, through UTHealth‐CCTS Grant Number UL1TR003167 and contract U01CK000692, Centers for Disease Control and Prevention. The contents of this publication are solely the responsibility of the authors and do not necessarily represent the official views of the NIH or CDC.
Supporting information
**Table S1:**Genes with reported mutations associated with antifungal resistance.
**Table S2:**Sociodemographic characteristics of the cohort.
**Table S3:**Treatment characteristics and clinical outcomes between study groups.
**Figure S1:**Average Nucleotide Identity between the sequenced isolates and representative yeast species. Numbers in the boxes represent de ANI values (%).
**Figure S2:**Recombination‐masked SNP‐distance matrix of the sequenced isolates.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1I. Franconi , C. Rizzato , A. Tavanti , M. Falcone , and A. Lupetti , “Paradigm Shift: Candida Parapsilosis Sensu Stricto as the Most Prevalent Candida Species Isolated From Bloodstream Infections With Increasing Azole‐Non‐Susceptibility Rates: Trends From 2015–2022 Survey,” Journal of Fungi 9, no. 10 (2023): 1012, 10.3390/jof 9101012.37888268 PMC 10608153 · doi ↗ · pubmed ↗
- 2M. Asogan , H. Y. Kim , S. Kidd , et al., “ Candida parapsilosis : A Systematic Review to Inform the World Health Organization Fungal Priority Pathogens List,” Medical Mycology 62, no. 6 (2024): 131, 10.1093/mmy/myad 131.PMC 1121061638935912 · doi ↗ · pubmed ↗
- 3F. Daneshnia , J. N. de Almeida Júnior , M. Ilkit , et al., “Worldwide Emergence of Fluconazole‐Resistant Candida parapsilosis : Current Framework and Future Research Roadmap,” Lancet Microbe 4, no. 6 (2023): e 470–e 480, 10.1016/S 2666-5247(23)00067-8.37121240 PMC 10634418 · doi ↗ · pubmed ↗
- 4S. G. Whaley , E. L. Berkow , J. M. Rybak , A. T. Nishimoto , K. S. Barker , and P. D. Rogers , “Azole Antifungal Resistance in Candida albicans and Emerging Non‐albicans Candida Species,” Frontiers in Microbiology 7 (2017): 2173, 10.3389/fmicb.2016.02173.28127295 PMC 5226953 · doi ↗ · pubmed ↗
- 5D. Yamin , M. H. Akanmu , A. Al Mutair , S. Alhumaid , A. A. Rabaan , and K. Hajissa , “Global Prevalence of Antifungal‐Resistant Candida parapsilosis: A Systematic Review and Meta‐Analysis,” Tropical Medicine and Infectious Disease 7, no. 8 (2022): 188, 10.3390/tropicalmed 7080188.36006280 PMC 9416642 · doi ↗ · pubmed ↗
- 6R. d. J. Treviño‐Rangel , E. Garza‐González , J. G. González , V. Bocanegra‐García , J. M. Llaca , and G. M. González , “Molecular Characterization and Antifungal Susceptibility of the Candida parapsilosis Species Complex of Clinical Isolates From Monterrey, Mexico,” Medical Mycology 50, no. 7 (2012): 781–784, 10.3109/13693786.2012.675526.22493945 · doi ↗ · pubmed ↗
- 7D. E. Corzo‐Leon , M. Peacock , P. Rodriguez‐Zulueta , G. J. Salazar‐Tamayo , and D. M. Mac Callum , “General Hospital Outbreak of Invasive Candidiasis due to Azole‐Resistant Candida parapsilosis Associated With an Erg 11 Y 132F Mutation,” Medical Mycology 59, no. 7 (2021): 664–671, 10.1093/mmy/myaa 098.33305313 PMC 8257413 · doi ↗ · pubmed ↗
- 8Clinical and Laboratory Standards Institute , Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed. (CLSI, 2017), 1–46.