Unveiling the hidden diversity of neotropical Steccherinum and allied genera (Steccherinaceae, Basidiomycota)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
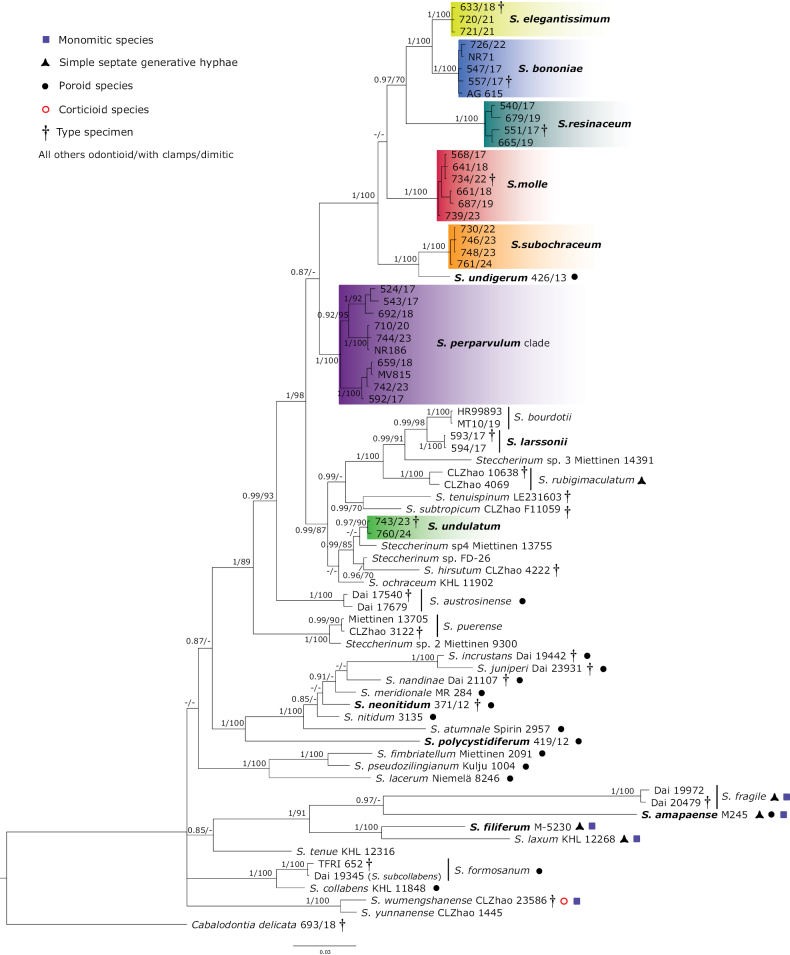 Figure 1
Figure 1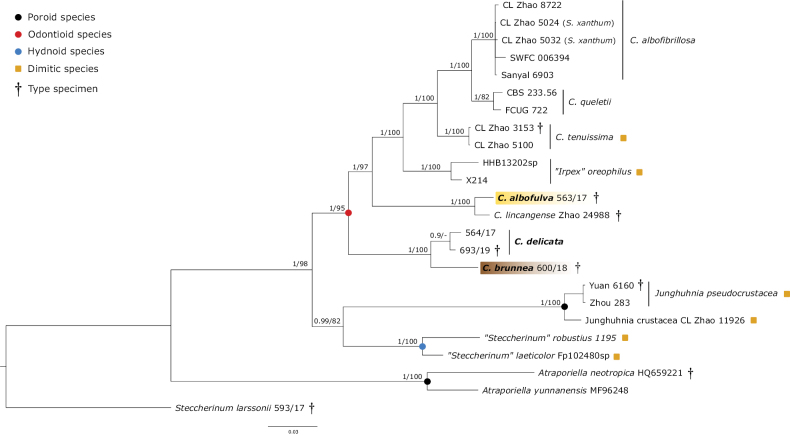 Figure 2
Figure 2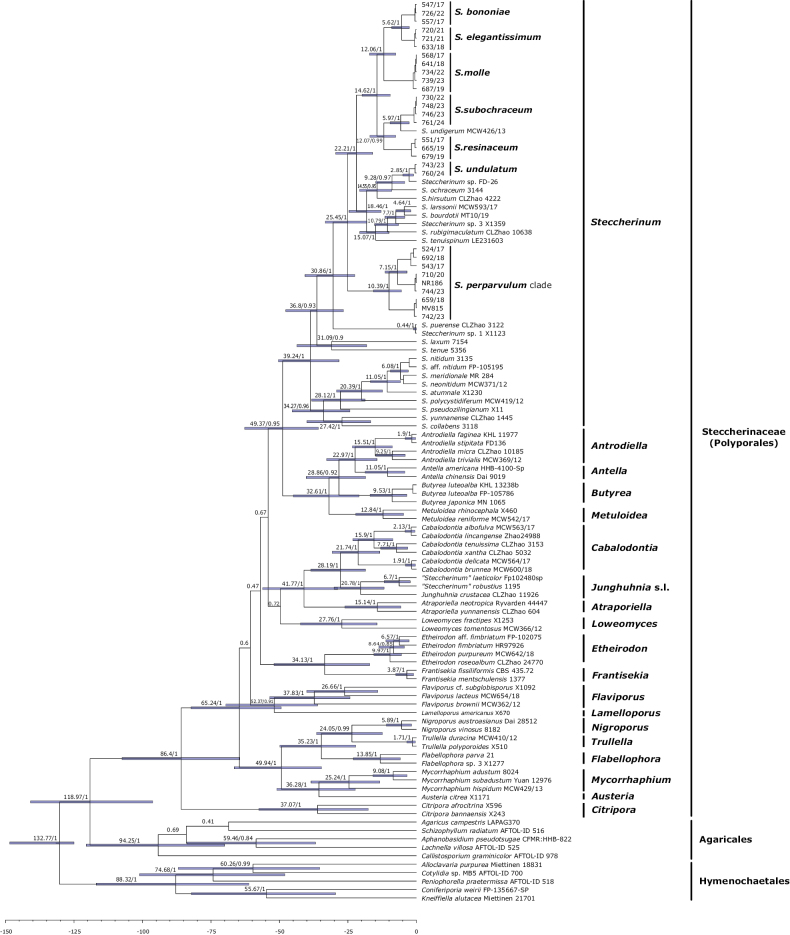 Figure 3
Figure 3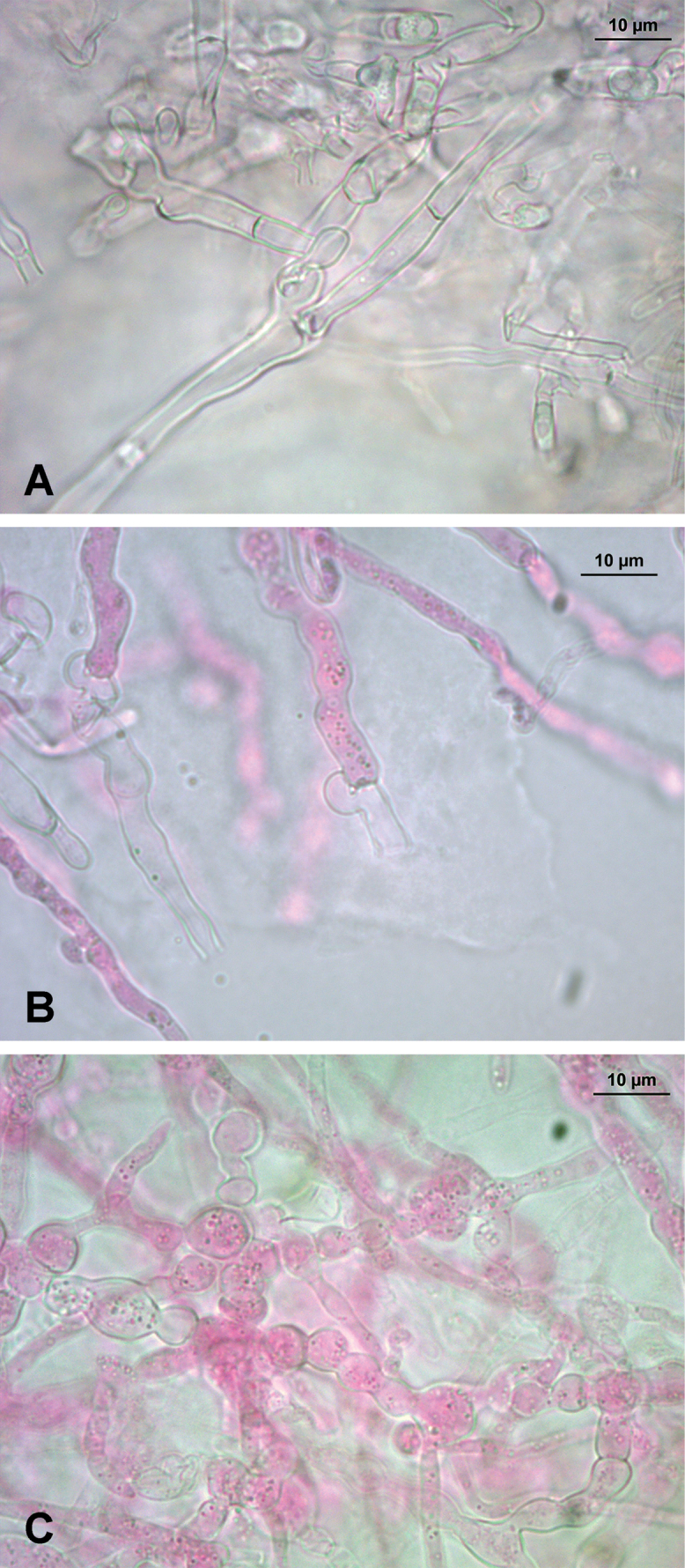 Figure 4
Figure 4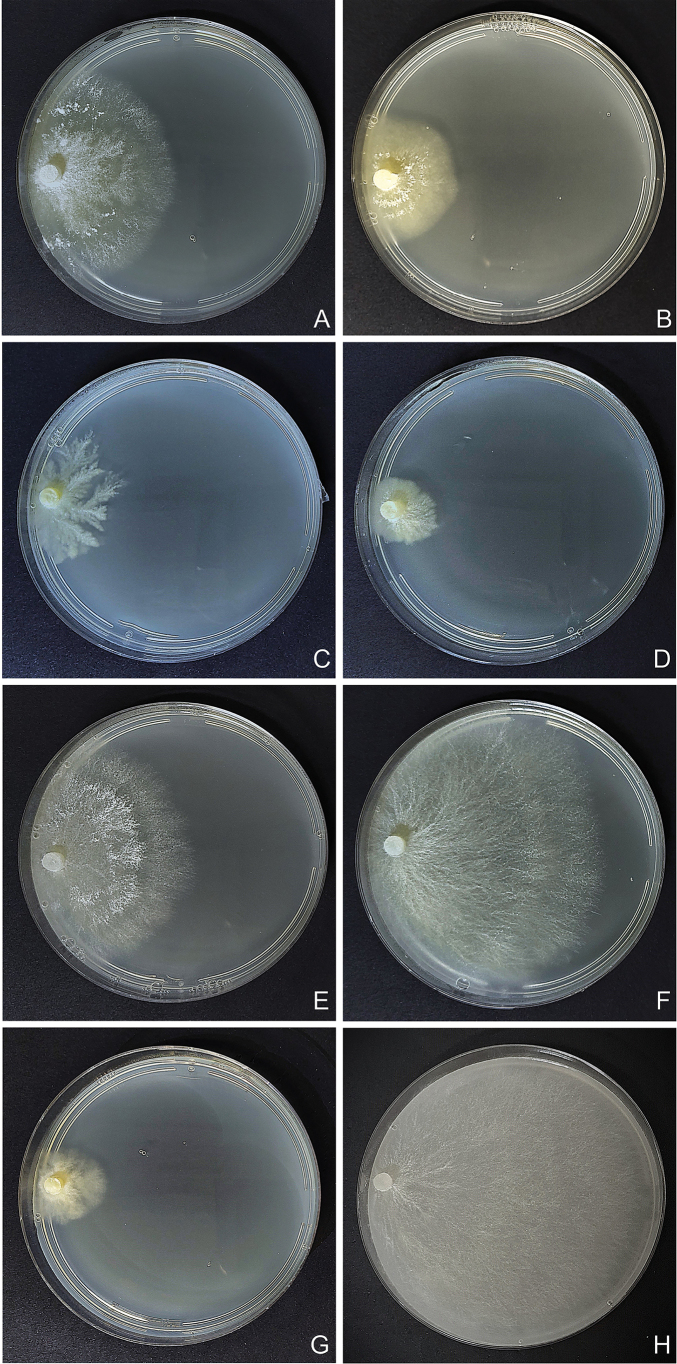 Figure 5
Figure 5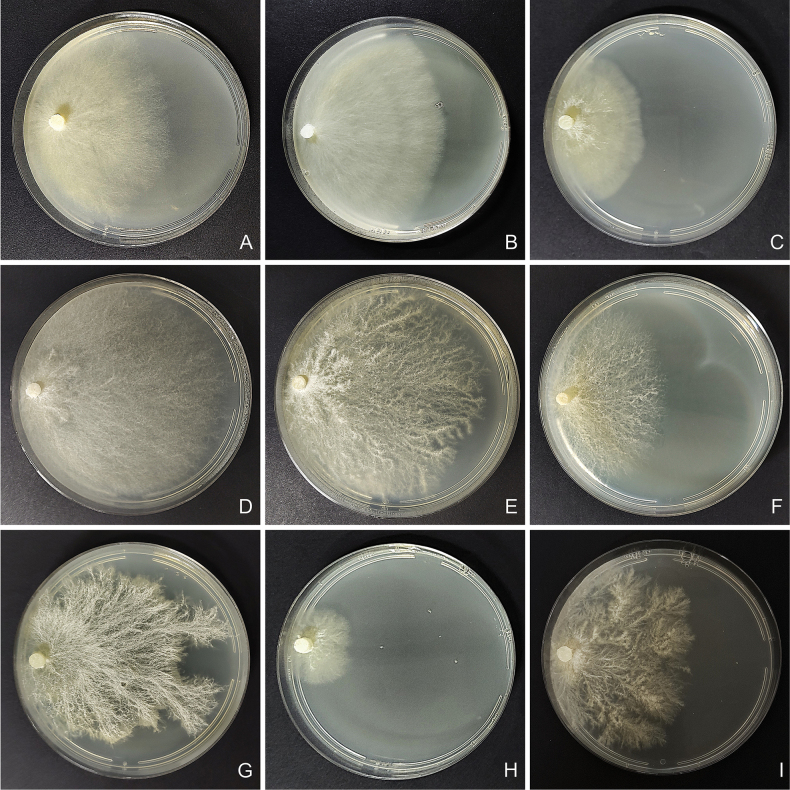 Figure 6
Figure 6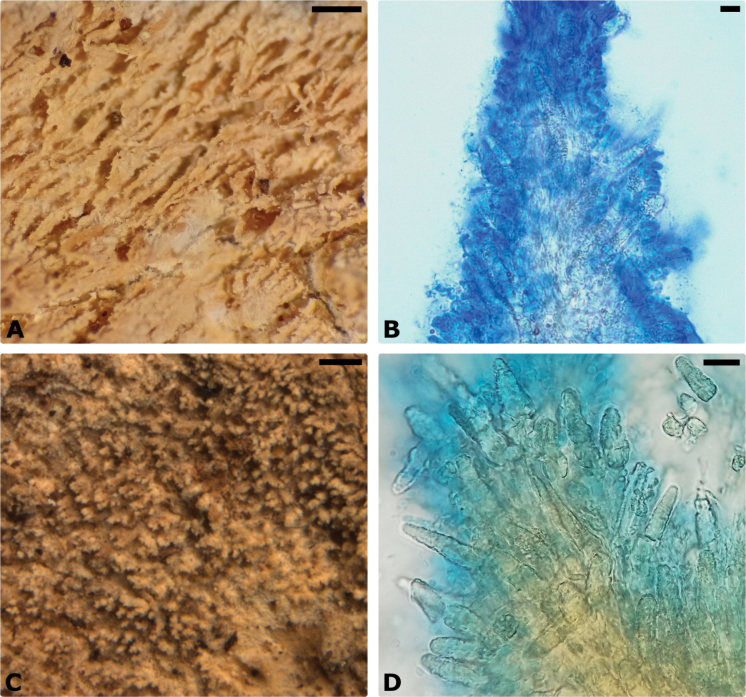 Figure 7
Figure 7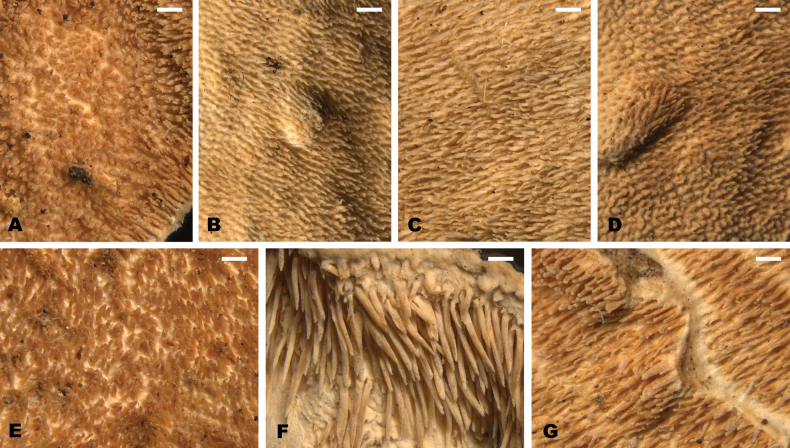 Figure 8
Figure 8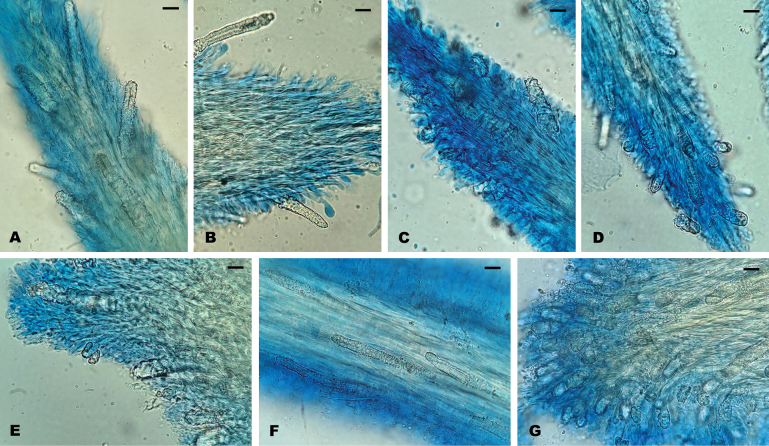 Figure 9
Figure 9 Figure 10
Figure 10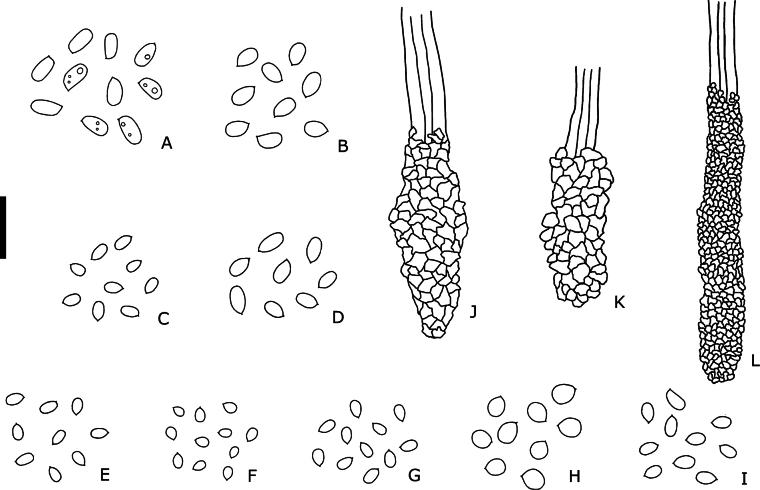 Figure 11
Figure 11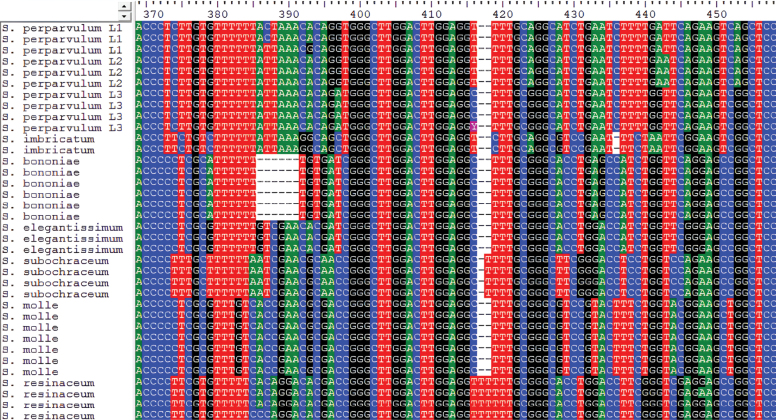 Figure 12
Figure 12| Specimen | Voucher | Loc. | GenBank No. | References | ||||
|---|---|---|---|---|---|---|---|---|
| ITS | 28S |
|
| mtSSU | ||||
|
| LAPAG370 | - |
|
|
|
| ||
|
| Miettinen 18831 | US |
|
|
|
|
|
|
|
| HHB-4100-Sp | US |
|
|
|
| ||
|
| Dai 9019 | CN |
|
|
| |||
|
| KHL 11977 | NO |
|
|
|
|
| |
|
| CLZhao 10185 | CN |
|
|
|
|
| Wu and Zhao (unpub.) |
|
| FD–136 | US |
|
|
|
| ||
|
| MCW 369/12 | BR |
|
|
|
| ||
|
| CFMR:HHB-822 | US |
|
|
|
|
| |
|
| Ryvarden 4447 | BZ |
|
| Miettinen and Rajchenberg 2012 | |||
|
| CLZhao 604 | CN |
|
|
|
|
| |
|
| X1171 | NZ |
|
|
| |||
|
| FP–105786 | US |
|
|
|
| ||
|
| KHL 13238b | EE |
|
|
|
|
| |
|
| MN 1065 | JP |
|
|
|
|
| |
|
| CLZhao 5032 | CN |
|
|
|
|
|
|
|
| CLZhao 5024 | CN |
|
|
|
|
|
|
|
| CLZhao 8722 | CN |
|
| Wu and Zhao (unpub.) | |||
|
| SWFC 006394 | CN |
| Zhao (unpub.) | ||||
|
| Sanyal 6903 | IN |
| Sanyal (unpub.) | ||||
|
| MCW 563/17 | BR |
|
|
|
| This study | |
|
| MCW 600/18 | BR |
|
|
| This study | ||
|
| MCW 693/19 | BR |
|
|
|
|
| |
|
| MCW 564/17 | BR |
|
|
|
|
|
|
|
| Zhao24988 | CN |
|
|
|
|
|
|
|
| FCUG 722 | SE |
| Hallenberg and Parmasto (unpub.) | ||||
|
| CBS: 233.56 | FR |
|
|
| |||
|
| CLZhao 5100 | CN |
|
|
|
| ||
|
| CLZhao 3153 | CN |
|
|
|
|
| |
|
| AFTOL-ID 978 | US |
|
|
|
|
| |
|
| KHL-GB | SE |
|
|
|
|
| |
|
| X596 | UG |
|
|
|
|
| |
|
| X243 | CN |
|
|
| |||
|
| FP-135667-SP | US |
|
|
|
|
|
|
| AFTOL-ID 700 | US |
|
|
|
| Wang et al. (unpub.) | ||
|
| HR97926 | CZ |
|
|
|
| ||
|
| HR98811 | CZ |
|
|
|
| ||
|
| FP-102075 | US |
|
|
|
|
| |
|
| MCW 642/18 | BR |
|
|
|
| ||
|
| CLZhao 24770 | CN |
|
|
|
|
|
|
|
| 21 | BR |
|
| Saragiotto (unpub.) | |||
| X1277 | ID |
|
|
|
| |||
|
| MCW362/12 | BR |
|
|
|
| ||
|
| MCW654/18 | BR |
|
|
|
| ||
|
| X1092 | BR |
|
|
|
| ||
|
| CBS 435.72 | US |
|
|
| |||
|
| BRNM 710170 | CZ |
|
|
|
| ||
|
| X214 | FI |
|
|
| |||
|
| HHB13202sp | US |
|
|
| |||
|
| CLZhao 11926 | CN |
|
|
|
| Wu and Zhao (unpub.) | |
|
| Yuan 6160 | CN |
|
| ||||
|
| Zhou 283 | CN |
|
| ||||
|
| Miettinen 21701 | FI |
|
|
|
|
|
|
|
| AFTOL-ID 525 | NL |
|
|
|
|
| |
|
| X670 | EC |
|
|
| |||
|
| X1253 | US |
|
|
|
| ||
|
| MCW366/12 | BR |
|
|
| |||
|
| MCW 542/17 | BR |
|
|
|
|
| |
|
| X460 | AU |
|
|
|
| ||
|
| 8024 | US |
|
|
|
|
| |
|
| MCW429/13 | BR |
|
|
|
| ||
|
| Yuan 12976 | CN |
|
|
| |||
|
| Dai 28512 | CN |
|
|
|
| ||
|
| 8182 | US |
|
|
|
|
| |
|
| AFTOL-ID 518 | - |
|
|
|
| Nilsson et al. (unpub.) | |
|
| AFTOL-ID 516 | PA |
|
|
|
|
| |
|
| M245 | BR |
|
|
| |||
|
| Spirin 2957 | RU |
|
|
|
|
| |
|
| Dai 17540 | CN |
|
|
| |||
|
| Dai 17540 | CN |
|
|
| |||
|
| AG 1615 | BR |
|
| This study | |||
|
| MCW 547/17 | BR |
|
|
| This study | ||
|
| MCW 557/17 | BR |
|
|
|
| This study | |
|
| MCW 726/22 | BR |
|
|
| This study | ||
|
| NR71 | BR |
|
| This study | |||
|
| MV446 | BR |
| This study | ||||
|
| HR 99893 | CZ |
|
|
|
| ||
|
| MT 10/19 | CZ |
|
|
|
| ||
|
| KHL 11848 | SE |
|
|
|
|
| |
|
| MCW 633/18 | BR |
|
|
|
| This study | |
|
| MCW 720/21 | BR |
|
|
| This study | ||
|
| MCW 721/21 | BR |
|
|
| This study | ||
|
| M-5230 | EC |
|
| ||||
|
| Miettinen 2091 | RU |
|
|
| |||
|
| Dai 19345 | CN |
|
|
| |||
|
| TFRI 652 | – |
|
| Chou et al. (unpub.) | |||
|
| CLZhao 4222 | CN |
|
|
|
|
|
|
|
| Dai 19442 | CN |
|
|
| |||
|
| Dai 23931 | CN |
|
|
| |||
|
| Niemelä 8246 | FI |
|
|
| |||
|
| Fp102480sp | US |
|
|
|
| ||
|
| MCW 593/17 | BR |
|
|
|
|
| |
|
| MCW 594/17 | BR |
|
|
|
| ||
|
| KHL 12268 | US |
|
|
|
|
| |
|
| MR 284 | AR |
|
|
|
| ||
|
| MCW 568/17 | BR |
|
|
| This study | ||
|
| MCW 641/18 | BR |
|
|
| This study | ||
|
| MCW 661/18 | BR |
|
| This study | |||
|
| MCW 687/19 | BR |
|
|
| This study | ||
|
| MCW 734/22 | BR |
|
|
| This study | ||
|
| MCW 739/23 | BR |
|
|
| This study | ||
|
| MV450 | BR |
| This study | ||||
|
| Dai 21107 | CN |
|
|
| |||
|
| MCW 371/12 | BR |
|
|
|
| ||
|
| KHL 11903 | SE |
|
|
|
|
| |
|
| FP-105195-Sp | US |
|
|
|
| ||
|
| SE |
|
|
|
|
| ||
|
| 524/17 | BR |
|
|
|
| This study | |
|
| 592/17 | BR |
|
| This study | |||
|
| 659/18 | BR |
|
|
|
| This study | |
|
| 692/19 | BR |
|
|
| This study | ||
|
| 543/17 | BR |
|
|
| This study | ||
|
| 710/20 | BR |
|
|
| This study | ||
|
| 742/23 | BR |
|
|
|
| This study | |
|
| 744/23 | BR |
|
|
| This study | ||
|
| MV728 | BR |
| This study | ||||
|
| MV815 | BR |
|
|
| This study | ||
|
| NR186 | BR |
|
| This study | |||
|
| MCW 419/12 | BR |
|
|
|
| ||
|
| Kulju 1004 | FI |
|
|
|
|
| |
|
| Miettinen 13705 | ID |
|
|
|
|
| |
|
| CLZhao 3122 | CN |
|
|
| |||
|
| MCW 540/17 | BR |
|
| This study | |||
|
| MCW 551/17 | BR |
|
|
|
| This study | |
|
| MCW 665/19 | BR |
|
|
| This study | ||
|
| MCW 679/19 | BR |
|
|
| This study | ||
|
| 1195 | SE |
|
|
| |||
|
| CLZhao 10638 | CN |
|
|
|
| ||
|
| CLZhao 4069 | CN |
|
|
| |||
|
| 730/22 | BR |
|
| This study | |||
|
| 746/23 | BR |
|
|
|
| This study | |
|
| 748/23 | BR |
|
|
|
| This study | |
|
| 761/24 | BR |
|
|
|
| This study | |
|
| CLZhao | CN |
|
|
| |||
|
| KHL 12316 | US |
|
|
|
|
| |
|
| RU |
|
|
|
| |||
|
| MCW 436/13 | BR |
|
|
|
| ||
|
| MCW 743/23 | BR |
|
|
| This study | ||
|
| MCW 760/24 | BR |
|
|
|
| This study | |
|
| CLZhao 23586 | CN |
|
|
| |||
|
| CLZhao 1445 | CN |
|
|
|
|
| |
| FD-26 | US |
|
|
|
| |||
| Miettinen 9300 | ID |
|
|
| ||||
| Miettinen 14391 | ID |
|
|
|
| |||
| Miettinen 13755 | ID |
|
|
| ||||
|
| MCW410/12 | BR |
|
|
| |||
|
| X510 | VE |
|
|
| |||
| Node | Mean of stem age / 95% | PP | Mean of crown age / 95% | PP | Period |
|---|---|---|---|---|---|
|
| 118.97 (140.89–96.26) | 1 | 86.4 (107.49–67.49) | 1 | Late Cretaceous |
|
| 49.37 (62.69–35.79) | 0.95 | 39.24 (50.4–28.27) | 1 | Eocene |
|
| 28.19 (38.57–18.59) | 1 | 21.74 (30.76–13.37) | 1 | Oligocene |
| “ | 49.37 (62.69–35.79) | 0.95 | 32.61 (44.97–20.95) | 1 | Eocene |
| “ | 41.77 (56.13–28.76) | 1 | 28.19 (38.57–18.59) | 1 | Eocene |
|
| - | - | 27.76 (42.34–14.4) | 1 | Oligocene |
|
| - | - | 34.13 (52.0–17.04) | 1 | Eocene |
|
| - | - | 52.37 (69.56–36.15) | 0.91 | Eocene |
| “ | 65.24 (82.26–49.37) | 1 | 49.94 (66.53–34.63) | 1 | Paleocene |
|
| 86.4 (107.49–64.89) | 1 | 37.07 (57.52–17.59) | 1 | Eocene |
| Week 1 | Week 3 | Week 6 | |||||
|---|---|---|---|---|---|---|---|
| Species | Specimen | Avg. growth (mm) | Clamps | Avg. growth (mm) | Clamps | Avg. growth (mm) | Clamps |
|
| NR71 | 8.2 | - | 35 | - | 73.5 | - |
| 726/22 | 4.5 | + | 17 | + | 49.5 | + | |
|
| 720/21 | 1.7 | + | 19 | + | 39.2 | + |
| 721/21 | 1.3 | + | 12 | + | 22.2 | + | |
|
| 734/22 | 10 | + | 40 | + | 68.2 | + |
| 739/23 | 18.2 | + | 57 | + | 76 | + | |
|
| 710/20 | 9 | + | 39 | + | 65 | + |
| 742/23 | 13.2 | + | 52 | + | 79.3 | + | |
| 744/23 | 8 | + | 30 | + | 57.7 | + | |
|
| 551/17 | 5.1 | + | 27.5 | + | 71.5 | + |
| 665/19 | 9.5 | + | 31.5 | + | 65.7 | + | |
| 679/19 | 6.6 | + | 29 | + | 74 | + | |
|
| 730/22 | 16 | + | 71 | + | 75.3 | + |
| 748/23 | 4.5 | + | 16 | +/- | 30.5 | - | |
| 761/24 | 5.9 | + | 47 | + | 59 | + | |
|
| 743/23 | 0.8 | + | 13 | + | 36.25 | + |
| 760/24 | 16 | + | 69.2 | + | 77.4 | + | |
| Species | Basidiome | Teeth | Spores (Lm x Wm) | Cystidia |
|---|---|---|---|---|
|
| Resupinate to effused-reflexed. Corky to somewhat pliable. Pilei when present forming on the center and margins of the basidiomes. | Cylindrical, with acute to truncate apices, somewhat clustered and flattened. With an evident granulose aspect due to protuding cystidia; 4–6 per mm; Lm = 0.51 mm. | Ellipsoid. 3.1 × 2.0 µm. | Skeletocystidia elongated, covered with a thick cap of crystals, projecting above the hymenium; (20)35–65 × 5–9(10) µm. |
|
| Resupinate. Forming small patches on the substratum that fuse as they grow. Membranaceous and pliable. | Conical to filiform, with acute apices, solitary, very thin and waxy; 6–8 per mm; Lm = 0.41 mm. | Ellipsoid. 4.2 × 2.5 µm. | Skeletocystidia elongated, covered with a cap of thin crystals, projecting above the hymenium; 20–70(75) x 4–8 µm. |
|
| Resupinate to effused-reflexd. Pilei forming on the margins of the basidiomes. Waxy to corky. | Cylindrical, with round to acute apices, mostly solitary, 4–6 per mm; Lm = 0.5 mm. | Subglobose to broadly ellipsoid. 3.9 × 3.2 µm. | Skeletocystidia covered with a cap of crystals, protruding into the hymenium or slightly above it; 30–50(60) × 7–10 µm. |
|
| Resupinate. Usually forming large patches. Soft to membranaceous, pliable and easily tearing. | Cylindrical to filiform, with acute apices., mostly solitary, 5–7 per mm. Lm = 0.45. | Ellipsoid. 3.1 × 2.1 µm. | Skeletocystidia coarsely encrusted with large crystals, immersed in the trama or with the apices protruding into the hymenium; (20)30–60 × 7–13(15) µm. |
|
| Resupinate to effused-reflexed. Papery and pliable, easily tearing. When present, small pilei forming on the margins of the basidiomes. | Conical to cylindrical, with acute apices, solitary; 4–7 per mm; Lm = 0.43 mm (reaching up to 0.7 mm in effused-reflexed basidiomes). | Subglobose to broadly ellipsoid. 2.7 × 1.9 µm. | Skeletocystidia covered with a cap of crystals, immersed in the trama or with the apices projecting above the hymenium; (20)25–42(48) × (4)6–12(15) µm. |
|
| Resupinate. Membranaceous to papery and pliable when flesh, turning waxy and tough upon drying. | Cylindrical, with truncate to acute apices, often bifurcate and laterally fused; clustered when dried. 5–6 per mm; Lm = 0.52. | Ellipsoid. 3.0 × 2.0 µm. | Skeletocystidia coarsely encrusted with large crystals, immersed in the trama or with the apices protruding into the hymenium; 25–45(50) × (7)9–11(12) µm. |
|
| Effused-reflexed, with imbricate pilei. Papery to corky and somewhat pliable. | Cylindrical, with truncate to acute apices, solitary; 3–5 per mm; Lm = 1.52. | Subglobose. 3.8 × 3.3 µm. | Skeletocystidia elongated, covered with a thin cap of crystals, immersed in the trama or more rarely projecting outwards; 25–50 × 5–8(9) µm. |
|
| Effused-reflexed. With very small imbricate pilei forming a wavy pattern on the basidiomes. | Conical to cylindrical, with acute to round apices, sometimes flattened, solitary or arising from a common base; 4–6 per mm; Lm = 0.85 mm. | Ellipsoid. 3.6 × 2.3 µm. | Skeletocystidia covered with a cap of crystals, immersed in the trama or with the apices protruding into the hymenium; 20–60 × 7–10 µm. |
| Species/Specimen (voucher) | Length | Lm | Width | Wm | Q |
| n |
|---|---|---|---|---|---|---|---|
| 4.0–5.1(–5.3) | 4.5 | 2.3–2.9(–3.1) | 2.6 | (1.60–)1.63–1.85(–1.88) | 1.75 | 39 | |
| (3.8–)3.9–4.9 | 4.3 | 2.4–3.0(–3.1) | 2.8 | (1.38–)1.39–1.76(–1.78) | 1.56 | 36 | |
|
| 2.6–3.6 | 3.1 | 1.8–2.5 | 2 | 1.38–1.65 | 1.51 | 137 |
| NR71 | (2.8–)3.0–3.5 | 3.2 | (1.8–)1.9–2.3(–2.4) | 2.1 | 1.43–1.60(–1.63) | 1.54 | 24 |
| 547/17 | (2.5–)2.8–3.4(–3.5) | 3 | 1.8–2.4 | 2 | (1.39–)1.40–1.63 | 1.51 | 25 |
| 557/17 | (2.7–)2.8–3.6 | 3.1 | 1.9–2.3(–2.4) | 2 | (1.38–)1.40–1.63(–1.63) | 1.52 | 34 |
| AL1615 | (2.8–)2.9–3.3 | 3.1 | 1.9–2.2 | 2.1 | (1.43–)1.45–1.58(–1.60) | 1.5 | 30 |
| 726/22 | (2.7–)2.8–3.2(–3.4) | 3 | (1.8–)1.9–2.3(–2.4) | 2.1 | 1.38–1.60 | 1.46 | 24 |
|
| (3.4–)3.5–5.0(–5.1) | 4.2 | 2.2–2.9(–3.0) | 2.5 | (1.42–)1.35–1.88(–1.92) | 1.64 | 94 |
| 633/18 | (3.4–)3.5–4.0 | 3.7 | 2.2–2.5(–2.7) | 2.4 | 1.48–1.68(–1.73) | 1.57 | 20 |
| 720/21 | (3.5–)3.6–5.0(–5.1) | 4.4 | (2.2–)2.3–2.9(–3.0) | 2.6 | 1.42–)1.43–1.88(–1.92) | 1.66 | 50 |
| 721/21 | (3.8–)3.9–4.6(–4.7) | 4.2 | (2.3–)2.4–2.6(–2.7) | 2.5 | (1.50–)1.52–1.81 | 1.68 | 24 |
|
| 3.5–4.3(–4.5) | 3.9 | 3–3.7(–4.0) | 3.2 | (1.13–)1.15–1.28(–1.30) | 1.21 | 220 |
| 3.3–3.9(–4.0) | 3.6 | 2.7–3.3 | 3 | 1.16–1.24(–1.26) | 1.2 | 15 | |
| (3.5–)3.6–4.4(–4.6) | 3.9 | 3.0–3.6(–3.9) | 3.2 | (1.10–)1.13–1.28 | 1.21 | 20 | |
| KHL9806 | (3.6–)3.7–4.2(–4.3) | 3.9 | (3.0–)3.1–3.7(–3.9) | 3.4 | (1.10–)1.15–1.22(–1.23) | 1.16 | 15 |
| LR23000 | (3.6–)3.7–3.9 | 3.8 | 2.8–3.1(–3.3) | 3 | (1.15–)1.19–1.33(–1.36) | 1.27 | 15 |
| LR23024 | (3.6–)3.7–4.2 | 3.9 | (3.1–)3.2–3.6 | 3.4 | (1.10–)1.12–1.22(–1.24) | 1.16 | 15 |
| MV634 | (3.6–)3.7–4.3(–4.5) | 3.9 | 2.9–3.4(–3.50) | 3.1 | (1.17–)1.19–1.35(–1.38) | 1.28 | 35 |
| MWC593/17 | (3.5–)3.7–4.4(–4.5) | 4 | 3.0–3.6(–3.80) | 3.3 | (1.14–)1.17–1.27 | 1.21 | 25 |
| MWC594/17 | 3.6–4.3(–4.5) | 3.9 | (2.7–)2.0–3.5(–3.6) | 3.2 | 1.14–1.34(–1.40) | 1.23 | 25 |
| MWC621/17 | (3.5–)3.6–4.2(–4.3) | 3.9 | 3.1–3.6(–3.8) | 3.4 | (1.05–)1.10–1.22 | 1.15 | 30 |
| MWC676/19 | 3.5–4.4(–4.6) | 3.9 | (2.7–)2.8–3.4(–3.9) | 3.1 | (1.16–)1.18–1.34(–1.37) | 1.25 | 25 |
|
| 2.5–3.5(–3.6) | 3.1 | 1.8–2.4(–2.5) | 2.1 | 1.20–1.60 | 1.45 | 113 |
| 641/18 | 3.0–3.5(–3.6) | 3.3 | 2.0–2.4 | 2.3 | (1.41–)1.42–1.57(–1.60) | 1.47 | 32 |
| 661/18 | 3.1–3.5(–3.6) | 3.3 | 2.0–2.4 | 2.2 | (1.43–)1.45–1.60 | 1.49 | 19 |
| 734/22 | 2.9–3.3 | 3.1 | 2.0–2.3 | 2.1 | 1.36–1.55(–1.58) | 1.46 | 25 |
| 739/23 | (2.9–)3.0–3.6 | 3.3 | (2.0–)2.1–2.4(–2.5) | 2.3 | (1.33–)1.36–1.52(–1.58) | 1.46 | 11 |
| MV450 | (2.4–)2.5–2.9(–3.0) | 2.7 | 1.8–2.0(–2.1) | 1.9 | (1.20–)1.25–1.53(–1.56) | 1.38 | 26 |
|
| 2.3–3.0(3.2) | 2.7 | 1.7–2.2 | 1.9 | 1.25–1.55(1.58) | 1.42 | 204 |
| 524/17 | 2.3–2.9(–3.0) | 2.6 | 1.7–2.2 | 1.9 | 1.25–1.42 | 1.34 | 30 |
| LR24589 (holotype) | 2.3–2.9(–3.0) | 2.6 | 1.7–2.1(–2.2) | 1.9 | (1.29–)1.30–1.47(–1.50) | 1.38 | 35 |
| 659/18 | 2.6–3.0(–3.1) | 2.8 | (1.7)1.8–2.1 | 1.9 | (1.38–)1.40–1.53 | 1.46 | 35 |
| MV815 | 2.3–2.8(–2.9) | 2.6 | 1.7–2.1(–2.2) | 1.9 | (1.27–)1.30–1.50(–1.53) | 1.4 | 30 |
| 592/17 | 2.7–3.2 | 3 | (1.8–)1.9–2.2 | 2 | (1.30–)1.33–1.53 | 1.46 | 34 |
| 710/20 | (2.5–)2.6–3.0 | 2.8 | 1.8–2.1 | 1.9 | 1.37–1.53(1.58) | 1.44 | 25 |
| 742/23 | (2.6–)2.7–2.9 | 2.8 | 1.9–2.1 | 2 | (1.37–)1.38–1.47(–1.53) | 1.42 | 15 |
|
| 2.7–3.4(–3.5) | 3 | 1.7–2.4 | 2 | 1.38–1.59(–1.60) | 1.49 | 52 |
| 551/17 | 2.8–3.4(–3.5) | 3.1 | 1.8–2.4 | 2.1 | 1.38–1.58(–1.60) | 1.47 | 35 |
| 665/19 | 2.7–3.0 | 2.8 | 1.7–2.0(–2.1) | 1.8 | (1.38–)1.42–1.59 | 1.54 | 17 |
|
| (3.2–)3.3–4.2 | 3.8 | 2.5–3.8 | 3.3 | 1.06–1.24(–1.30) | 1.15 | 93 |
| PACA 22824 | 3.7–4.0 | 3.8 | 3.0–3.5 | 3.2 | 1.14–1.23 | 1.19 | 5 |
| SP 97591 (holotype) | (3.2–)3.4–3.8(–4.0) | 3.6 | 2.5–3.5 | 3.1 | (1.06–)1.09–1.23(–1.30) | 1.15 | 20 |
| 730/22 | (3.3–)3.4–4.1 | 3.8 | (2.9–)3.0–3–8(–4.1) | 3.4 | (1.06–)1.08–1.19(–1.20) | 1.13 | 32 |
| 746/23 | (3.5–)3.7–4.2 | 3.9 | (3.0–)3.2–3.7 | 3.4 | 1.11–1.24(–1.29) | 1.16 | 24 |
| 748/23 | 3.2–3.9 | 3.6 | 2.6–3.4 | 3.1 | (1.12–)1.14–1.19(–1.22) | 1.16 | 12 |
|
| 3.3–4.1 | 3.6 | 2.0–2.7(–2.8) | 2.3 | (1.37–)1.39–1.65(–1.67) | 1.52 | 72 |
| 743/23 | 3.3–3.8 | 3.5 | 2.0–2.6 | 2.3 | 1.42–1.67 | 1.54 | 38 |
| 760/24 | (3.2–)3.3–3.8(–4.7) | 3.5 | 2.1–2.7(–2.8) | 2.4 | (1.37–)1.39–1.57(–1.64) | 1.5 | 34 |
| Monosporic culture n. |
|
|
|
|
|
|
|---|---|---|---|---|---|---|
|
| + | + | - | - | - | - |
|
| + | + | + | - | - | - |
|
| + | + | + | - | - | - |
|
| + | + | + | - | - | - |
|
| - | - | - | |||
|
| - | - | - | |||
|
| - | - | - |
| 1 | Basidiomes soft and brittle, hyphal system monomitic to pseudo-dimitic, cystidia tapering towards the apex or more rarely clavate |
|
| – | Basidiomes membranaceous to waxy, hyphal system dimitic, cystidia clavate |
|
| 2 | Hymenophore in shades of brown, farinaceous, aculei cluttered and somewhat indistinct, hyphae golden yellow in mass |
|
| – | Hymenophore white to beige, not farinaceous, hyphae hyaline, aculei distinct |
|
| 3 | Aculei up to 0.4 mm long, basidiospores subglobose to oblong-ellipsoid, 3.5–4(–4.5) × 2.5–3(–3.5) μm |
|
| – | Aculei longer, up to 0.8 mm, basidiospores ellipsoid to narrowly-ellipsoid, (4–)4.3–5.1(–5.3) × 2.4–3(–3.1) μm |
|
| 4 | Aculei long, conspicuous, readily visible to the naked eye, mostly ≥ 1.5 mm |
|
| – | Aculei shorter, inconspicuous, or not readily visible to the naked eye, ≤ 1.0 mm. |
|
| 5 | Basidiomes effused-reflexed, pilei small, imbricate, forming a wavy pattern, aculei 0.75–1.0 mm long (Lm = 0.85 mm) |
|
| – | Basidiomes resupinate to effused-reflexed, pilei absent or marginal, not in a wavy pattern, aculei up to 0.75 mm long (Lm < 0.7 mm) |
|
| 6 | Basidiospores 3.5–5.0 × 2.5–4.0 μm |
|
| – | Basidiospores smaller, 2.3–3.5 × 1.7–2.5 μm |
|
| 7 | Basidiospores subglobose, 3.5–4.3 × 3–3.7 μm; cystidia 7–10 µm wide |
|
| – | Basidiospores ellipsoid, 3.5–5.0 × 2.2–2.9 µm; cystidia 4–6(8) µm wide |
|
| 8 | Cystidia coarsely encrusted with chunky crystals, embedded in the trama or slightly protruding into the hymenium |
|
| – | Cystidia evenly covered with thin to medium-sized crystals, mostly projecting above the hymenium |
|
| 9 | Basidiomes waxy, aculei often bifurcate or laterally fused, subiculum dense |
|
| – | Basidiomes soft, aculei not bifurcate or laterally fused, subiculum cottony |
|
| 10 | Basidiospores broadly ellipsoid 2.3–3.0(3.2) × 1.7–2.2 µm |
|
| – | Basidiospores ellipsoid 2.6–3.6 × 1.8–2.5 µm |
|
- —Fundação de Amparo à Pesquisa do Estado de São Paulo 501100001807 https://ror.org/02ddkpn78 http://doi.org/10.13039/501100001807
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico 501100003593 https://ror.org/03swz6y49 http://doi.org/10.13039/501100003593
- —Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco 501100006162 https://ror.org/02te5rf52 http://doi.org/10.13039/501100006162
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior 501100002322 https://ror.org/00x0ma614 http://doi.org/10.13039/501100002322
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Yeasts and Rust Fungi Studies · Lichen and fungal ecology
Introduction
The genus Steccherinum Gray, typified by S. ochraceum (Pers.) Gray, is typically characterized by resupinate to effused-reflexed basidiomes with a hymenophore composed of pores, spines (or aculei), or, more rarely, a smooth surface. The basidiomes are generally thin and pale-colored, though some species may display brighter orange hues. A key feature distinguishing Steccherinum is its dimitic hyphal structure and the presence of thick-walled clavate skeletocystidia in the trama, often projecting into or above the hymenium. In most species, the cystidia are heavily encrusted, although in some cases they may be completely smooth. Overall, all species in the genus exhibit somewhat similar micromorphology, which contributes to the complexity of its taxonomy (Ryvarden 1991; Miettinen et al. 2012; Westphalen et al. 2021). Despite this, many new species have been described in Steccherinum in recent years (Westphalen et al. 2018; Liu and Dai 2021; Westphalen et al. 2021; Wu et al. 2021a; Dong et al. 2022; Dong et al. 2023; Liu et al. 2023; Wang et al. 2024), making it the largest genus in the Steccherinaceae, with over 120 associated names in the Index Fungorum database.
Morphologically, Junghuhnia Corda shares several similarities with Steccherinum, with the two genera traditionally distinguished by the presence of a poroid hymenophore in Junghuhnia and a hydnoid in Steccherinum (Ryvarden 1991). However, molecular studies have demonstrated that Steccherinum exhibits considerable variation in hymenophore morphology, encompassing species with both pores and spines (Miettinen et al. 2012; Miettinen and Ryvarden 2016; Westphalen et al. 2021). Consequently, the morphological distinction between the two genera has become less clear, and molecular data currently support the recognition of only two species in Junghuhnia: J. crustacea (Jungh.) Ryvarden and J. pseudocrustacea H.S. Yuan (Yuan et al. 2019). Additionally, Cabalodontia Piątek and Etheirodon Banker, both genera within the Steccherinaceae, also present somewhat similar morphological features to Steccherinum. However, Cabalodontia can be differentiated by its more fragile basidiomes, a monomitic hyphal structure, and cystidia that are typically broader at the base, tapering toward the apex. In contrast, Etheirodon is characterized by purplish basidiomes with strongly fimbriate margins, as well as smaller cystidia with thinner walls (Westphalen et al. 2021).
Although Steccherinum and its allied genera form a large group with numerous taxa distributed in temperate and tropical regions, their diversity in the Neotropics remains poorly explored, with very few studies dedicated to the group. To date, ten Steccherinum species have been described from the region, including six odontioid to hydnoid taxa (S. basibadium Banker, S. diversum Hjortstam & Melo, S. filiferum Yurchenko & K.H. Larss., S. larssonii Westphalen & Motato-Vásq., S. perparvulum Hjortstam & Ryvarden, and S. subochraceum Bononi & Hjortstam) (Hjortstam 1999; Westphalen et al. 2021; Yurchenko et al. 2023) and four poroid taxa (S. amapaense A.M.S. Soares & Ryvarden, S. neonitidum Westphalen & Tomšovský, S. undigerum (Berk. & M.A. Curtis) Westphalen & Tomšovský, and S. polycystidiferum (Rick) Westphalen, Tomšovský & Rajchenb.) (Hyde et al. 2017; Westphalen et al. 2018). In addition, one species has been described in Etheirodon (E. purpureum Westphalen) and another in Cabalodontia (C. delicata Westphalen & Motato-Vásquez) (Westphalen et al. 2021). Nonetheless, many Steccherinum specimens deposited in Brazilian herbaria are misidentified as S. ochraceum, a species originally described from northern Europe with a temperate distribution. To address this gap, the present study aimed to expand the knowledge of Steccherinum diversity in the Neotropics, with emphasis on odontioid/hydnoid taxa, through morphological examination of specimens, mycelial culture studies, and phylogenetic analyses, including divergence-time estimates for the Steccherinaceae. Our findings support the occurrence of several morphologically similar cryptic species of Steccherinum in the Neotropics. Seven new species are described and discussed, with key diagnostic features provided for their morphological differentiation.
Materials and methods
Morphological studies
The studied specimens were collected in southern and southeastern Brazil between 2017 and 2024. Additional collections from SP, ICN, HURM, PACA, K, and O (abbreviations according to Thiers continuously updated) were examined for morphological revision and comparison. Sections of basidiomes were observed under a compound microscope. Cotton blue (Merck 1275) in a lactic acid solution were used to assess cyanophilic reactions of spores and hyphae (denoted as CB+ or CB−) and to measure microstructures. Since all species in the Steccherinaceae are negative in Melzer’s reagent, these reactions are not shown in the descriptions, although they were tested. A minimum of 25 measurements of each structure was taken when possible. Illustrations of the cystidia and basidiospores were prepared using a drawing tube at 1000× magnification. Measurement abbreviations and codes used are as follows: Lm × Wm = mean length and width; Q = range of length/width ratio; Qm = mean length/width ratio; and n = x/y (where x = number of measurements from y specimens). Numbers in parentheses indicate extreme sizes observed in less than 5% of the measurements.
DNA extraction and PCR amplification
DNA was extracted from either fungal cultures or small sections of dried basidiomes using a lysis buffer containing 2% CTAB, 1.4 M NaCl, 0.10 M Tris-HCl, and 20 mM EDTA, incubated at 65 °C for a minimum of 2 hours. After a chloroform extraction step, DNA was precipitated with isopropyl alcohol (Doyle 1987). PCR amplification of the ITS1-5.8S-ITS2 rDNA (ITS), 28S rDNA (LSU), and mtSSU regions was performed using primers ITS1/ITS4, LR0R/LR7, and MS1/MS2, respectively (Nikolcheva and Bärlocher 2004), following the methods outlined by Tomšovský et al. (2010). Amplification of the translation elongation factor 1-α (tef1-α) gene was carried out using the primers 983F/2218R or 983F/1567R (Matheny et al. 2007). The region spanning domains A and C of the largest RNA polymerase II subunit (rpb1) was amplified using primers rpb1-Af/rpb1-Cr (Matheny et al. 2002). For tef1-α and rpb1, a touchdown PCR protocol was used, in which the annealing temperature gradually decreased from 60 °C to 50 °C. After amplification, the products were purified with the ExoSAP-IT enzyme (Thermo Fisher), following the manufacturer’s protocols. Sequencing was performed by MacroGen Inc. (Seoul, South Korea), using the same primers as those employed in PCR amplification.
Culture studies
Spore prints from freshly collected specimens were obtained and used to prepare both monosporic and polysporic cultures. The cultures were grown on Malt Extract Agar (MEA) or Potato Dextrose Agar (PDA) at 25 °C. Intraspecimen mating system tests were conducted following Hallenberg (1984), using ten monosporic cultures in each confrontation. After identification of the mating types, monosporic culture confrontations among closely related taxa were carried out according to Hallenberg (1984). For mycelial growth rate analysis and morphological observation and comparison, culture preparation was adapted from the methods outlined by Nobles (1965). To this end, inocula measuring approximately four millimeters of mycelium were placed at the edge of 9 cm diameter plates containing MEA. For each specimen, five plates were prepared and incubated at 25 °C for six weeks. Weekly, the following characteristics were examined: 1) macromorphology of the mycelium; 2) growth rate expressed in millimeters; and 3) micromorphology of the inner and advancing zones of the mycelium. Growth rate measurements were performed in triplicate for each specimen, with each plate measured from the inoculum endpoint to the edge of the mycelium.
Phylogenetic analyses
Phylogenetic analyses were conducted using two datasets: one focused on the genus Steccherinum and another on Cabalodontia. The Steccherinum dataset included ITS, 28S, and tef1-α markers and encompassed all species with sequence data currently available. The Cabalodontia dataset included five molecular markers (ITS, 28S, tef1-α, rpb1, and mtSSU). Reference sequences were selected from Miettinen et al. (2012), Justo et al. (2017), and Westphalen et al. (2021), together with additional sequences retrieved through BLAST searches in the NCBI database. A summary of the sequences used in this study is provided in Table 1.
Sequence alignments were performed using MAFFT 7 online (http://mafft.cbrc.jp/alignment/server/) under the auto mode strategy. tef1-α introns were excluded from the analyses. The Steccherinum dataset was divided into three partitions: ITS, 28S, and tef1-α. The Cabalodontia dataset was divided into six partitions: ITS, 28S, tef1-α, rpb1, rpb1 introns, and mtSSU. Bayesian inference (BI) analyses were carried out in MrBayes 3.2.6 (Ronquist et al. 2012), with substitution models selected for each partition based on AICc values computed in jModelTest 2.1.4 (Darriba et al. 2012). The selected models were GTR + I + G for ITS and rpb1, TIM3 + I + G for 28S, TIM2 + I + G for tef1-α, and TPM2uf + I + G for rpb1 introns. The proportion of invariable sites (I) and gamma-distributed rates (G) were set according to the models selected for each partition. Four independent MCMC chains were run for 10 million generations, sampling every 1000 generations. The first 25% of trees were discarded as burn-in, and the remaining trees were used to generate a 50% majority-rule consensus tree. Posterior probabilities greater than 0.9 were considered strongly supported and values above 0.8 moderately supported. The Maximum Likelihood (ML) analysis was conducted in RAxML-HPC 8 (Stamatakis 2014) using a rapid bootstrap analysis and a search for the best-scoring ML tree. The same partitioning scheme used for BI was adopted, applying the GTRGAMMA model. Bootstrap values above 80% were considered statistically significant. All analyses were performed through the CIPRES Science Gateway portal (Miller et al. 2011).
Divergence time estimation
Divergence times were estimated using BEAST v2.7.7 (Bouckaert et al. 2019) with a five-gene dataset composed of ITS + 28S + tef1-α + rpb1 + mtSSU sequences aligned with MAFFT 7 online (Katoh and Standley 2013) under the auto mode strategy. Archaeomarasmius leggetti Hibbett, D. Grimaldi & Donoghue (Hibbett et al. 1997) and Quatsinoporites cranhamii S.Y. Sm., Currah & Stockey (Smith et al. 2004) were used as secondary fossil calibrations for Agaricales and Hymenochaetales, respectively. Six partitions were selected (ITS, 28S, tef1-α, rpb1, rpb1 introns, and mtSSU), and GTR + G was evaluated as the best-fit evolutionary model for the rpb1 introns partition and GTR + I + G for the other five partitions in jModelTest using the Corrected Akaike Information Criterion (AICc) (Darriba et al. 2012). Detailed parameters and fossil calibrations followed those used by Wang et al. (2023) and Li et al. (2025a) when generating the XML file in BEAUti v2. Two independent analyses of 100 million generations, sampling every 10,000 generations, were performed. Chain convergence was evaluated in Tracer v1.7.1 (Rambaut et al. 2018), and the two runs were combined, discarding 20% of states from each as burn-in, in LogCombiner v2.7.7 (Bouckaert et al. 2019), rendering a file with 8000 trees. A Maximum Clade Credibility (MCC) tree was then summarized, annotating clades with ≥ 0.8 posterior probability, in TreeAnnotator v2.7.7 (Bouckaert et al. 2019). The resulting tree was visualized in FigTree v1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/) to obtain the mean ages and 95% Highest Posterior Density (HPD) values (Drummond and Rambaut 2007). A 95% HPD marks the shortest interval that contains 95% of the sampled values. Since the main focus of this study was on Steccherinum and its related genera, taxa of Steccherinaceae with sequence data for at least three molecular markers were prioritized. However, for genera represented by only two available markers, those data were nevertheless included to ensure adequate representation in the analyses.
Results
The newly obtained molecular data revealed five new lineages in Steccherinum, represented by the new species S. bononiae, S. elegantissimum, S. molle, S. resinaceum, and S. undulatum (Fig. 1), and two new species in Cabalodontia: C. albofulva and C. brunnea (Fig. 2). Sequence data for S. perparvulum and S. subochraceum are presented here for the first time and confirm them as distinct lineages in Steccherinum (Fig. 1). Furthermore, S. lincangense and S. tenuissimum are nested within Cabalodontia and are combined into that genus.
Steccherinum phylogenetic tree of ITS–28S–tef1-α regions conducted by Bayesian analysis (for legends and numbers, see Table 1). Numbers at branches indicate Bayesian posterior probability and maximum likelihood bootstrap values. The scale bar indicates the number of expected substitutions per position. Neotropical taxa are highlighted in bold. Species with newly obtained sequence data are highlighted in colors. Type voucher specimens are indicated with a †.
Cabalodontia phylogenetic tree of ITS–28S–tef1-α–rpb1–mtSSU regions conducted by Bayesian analysis (for legends and numbers, see Table 1). Numbers at branches indicate Bayesian posterior probability and maximum likelihood bootstrap values. The scale bar indicates the number of expected substitutions per position. Neotropical taxa are highlighted in bold. New species are highlighted in colors. Type voucher specimens are indicated with a †.
In our phylogenetic analyses, S. xanthum nested in a single lineage with C. albofibrillosa (Fig. 1). The ITS sequences of both species, including the paratypes of S. xanthum (CLZhao 5032 and CLZhao 5024), are nearly identical, differing by only about two base pairs. Therefore, we consider them conspecific, with the older name C. albofibrillosa taking priority. The same applies to S. subcollabens, whose type material (Dai 19345) presents ITS sequences identical to the type of S. formosanum (TFRI 652) and represents a synonym of the latter (Fig. 1).
Notably, S. perparvulum encompasses three distinct lineages (Figs 1, 3)exhibiting variations in the ITS and tef1-α regions. However, no clear morphological differences or culture variation regarding morphology or growth rates were observed to distinguish these lineages. Mating tests conducted further corroborated that the observed genetic differences represent separate biological species. Consequently, we designated this group as the S. perparvulum species complex for now (see the Taxonomy section for further comments).
Maximum Clade Credibility (MCC) tree with divergence time estimations of Steccherinaceae inferred from Bayesian evolutionary analysis based on an ITS–28S–tef1-α–rpb1–mtSSU dataset. Mean ages of the nodes with at least 0.8 posterior probability were annotated along with the 95% highest posterior densities, which are marked by horizontal bars. Divergence time values and the scale bar indicate millions of years (Myr).
To compose the divergence-time estimation, we used a concatenated five-gene dataset, including 107 specimens, of which 97 belong to 19 different genera of the Steccherinaceae. Ten additional taxa from Agaricales and Hymenochaetales were used to represent the genetic diversity of the two secondary fossil calibrations used in the analysis. The MCC tree recovered (Fig. 3) estimates the ancestor of the Steccherinaceae with a stem age of approximately 118.97 Myr (95% height = 140.89–96.26 Myr) and a crown age of 86.4 Myr (95% height = 107.49–67.49 Myr), emerging in the early to late Cretaceous period. Estimated stem ages for its genera range from 86.4 Myr (95% height = 107.49–64.89 Myr) for Citripora Miettinen to 22.97 Myr (95% height = 32.76–14.39 Myr) for the Antrodiella/Antella clade. The genus Steccherinum was recovered with a mean stem age of 49.37 Myr (95% height = 62.69–35.79 Myr) and a mean crown age of 39.24 Myr (95% height = 50.4–28.27 Myr), placing its origin in the Eocene. The estimated ages for its species range from 31.09 Myr (95% height = 43.66–18.15 Myr) to 0.44 Myr (95% height = 1.07–0.02 Myr). On the other hand, Cabalodontia was recovered with a mean stem age of 28.19 Myr (95% height = 38.57–18.59 Myr) and a mean crown age of 21.74 Myr (95% height = 30.76–13.37 Myr), placing its origin in the Oligocene. The estimated ages for its species range from 7.1 Myr (95% height = 13.05–3.16 Myr) to 1.91 Myr (95% height = 3.74–0.53 Myr). Table 2 summarizes the estimated divergence times of the main nodes of the Steccherinaceae with at least 0.8 posterior probability. The international chronostratigraphic chart follows Cohen et al. (2013; updated) (URL: http://www.stratigraphy.org/ICSchart/ChronostratChart2022-10.pdf).
Table 2.: Estimated divergence times of main nodes of the Steccherinaceae. PP stands for “Posterior Probabilities”. Nodes with PP < 0.8 were not annotated.
Regarding the culture studies carried out, all mycelia showed very similar micromorphology, with thin-walled, regularly clamped hyphae and abundant intercalary chlamydospores in the older areas of the mycelium, becoming scarcer in the advancing zone (Fig. 4). However, some specimens exhibited loss of clamp connections. Specimen NR71 presented only simple septa during the six-week study, in contrast to specimen 726/22, both belonging to S. bononiae, which displayed regularly clamped hyphae. Specimen 748/23 of S. subochraceum exhibited variation in the presence or absence of clamp connections, with a total loss of clamps after the third week of growth. However, the mycelium was checked again two months after the end of the study, and the hyphae were regularly clamped once more. Regarding the growth rates, specimens 742/23 (S. perparvulum), 730/22 (S. subochraceum), and 760/24 (S. undulatum) showed the highest rates, whereas specimens 721/21 (S. elegantissimum), 748/23 (S. subochraceum), and 743/23 (S. undulatum) showed the lowest rates (Table 3). Overall, all cultures presented a similar macromorphology, exhibiting thin mats with floccose mycelia (Figs 5, 6). In contrast, two specimens of S. subochraceum (730/22 and 761/24) presented strongly fimbriate and denser mycelia (Fig. 6G, I), similar to specimen 665/19 of S. resinaceum (Fig. 6E). Interestingly, S. subochraceum displayed notable intraspecific variation, with differences in both mycelial macromorphology and growth rates (Fig. 6G–I, Table 3).
Microscopic features of Steccherinum spp. cultures. A generative hyphae with simple septa in strain 748/23 (S. subochraceum) B generative hyphae with clamps in strain 761/24 (S. subochraceum) C intercalary chlamydospores in strain 665/19 (S. resinaceum).
Macroscopic aspect of Steccherinum spp. cultures at six weeks. A, BS. bononiae. C, DS. elegantissimum. E, FS. molle. G, HS. undulatum. A 726/22. B NR71. C 720/21. D 721/21. E 734/22. F 739/23. G 743/23. H 760/24.
Macroscopic aspect of Steccherinum spp. cultures. A–CS. perparvulum. D–FS. resinaceum. G–IS. subochraceum. A 710/20. B 742/23. C 744/23. D 551/17. E 665/19. F 679/19. G 730/22. H 748/23. I 761/24.
Full descriptions and comments on the new taxa are presented below, as well as discussions of species with newly obtained molecular data (S. subochraceum and S. perparvulum). A summary of the main features of morphologically similar hydnoid neotropical Steccherinum spp. is provided in Table 4, and the basidiospore measurements obtained in this study are shown in Table 5. An identification key to Brazilian corticioid species of Steccherinum and Cabalodontia is also presented. For poroid species of Steccherinum, see Westphalen et al. (2018).
Taxonomy
Cabalodontia
albofulva
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & Gugliotta sp. nov.
8A3C1C10-5A46-5595-8925-2C6A8D0C996E
861265
Etymology.
Refers to the white basidiomes that become yellowish when dried.
Macroscopic and microscopic aspects of Cabalodontia spp. A, BC. albofulva (563/17). C, DC. brunnea (600/17). Scale bars: 0.5 mm (A, C); 10 µm (B, D).
Aculei in hymenophores of Neotropical Steccherinum spp. AS. bononiae (726/22). BS. elegantissimum (721/21). CS. molle (734/22). DS. perparvulum (710/20). ES. resinaceum (665/19). FS. subochraceum (746/23). GS. undulatum (743/23). Scale bars: 0.5 mm.
Detail of the trama and cystidia of Neotropical Steccherinum spp. AS. bononiae (726/22). BS. elegantissimum (721/21). CS. molle (734/22). DS. perparvulum (710/20). ES. resinaceum (665/19). FS. subochraceum (746/23). GS. undulatum (743/23). Scale bars: 10 µm.
Fresh basidiomes in situ. AS. subochraceum. BS. undulatum. Scale bars: 0.5 cm.
Microscopic features of Neotropical Steccherinum and Cabalodontia species. A–I Basidiospores. J–L Skeletocystidia. AC. albofulva. BC. brunnea. CS. bononiae. DS. elegantissimum. ES. molle. FS. perparvulum. GS. resinaceum. HS. subochraceum. IS. undulatum. J Coarsely encrusted tapering cystidia (C. brunnea, 600/17). K Skeletocystidia encrusted with large crystals (S. molle, 734/22). L Skeletocystidia covered with thin crystals (S. elegantissimum, 721/21). Scale bar: 10 µm.
Diagnosis.
Differs from C. queletii by thinner and more fragile aculei and smaller basidiospores.
Typification.
Brazil. Rio Grande do Sul: São Francisco de Paula, Parador Hampel, 19 Jun. 2017, M.C. Westphalen 563/17 (SP 467080).
Description.
Basidiomes adnante, annual, resupinate, not detaching, forming small patches on the substrate, very soft and brittle; sterile margins white, membranaceous, up to 1 mm wide. Hymenophore odontioid, white to cream when fresh, becoming yellowish to beige upon drying. Aculei thin, up to 0.8 mm long, somewhat clustered and very brittle upon drying, 6–8 per mm. Subiculum, white to cream, homogeneous, cottony, thin and fragile, up to 0.5 mm thick.
Hyphal system monomitic, hyphae loosely arranged; generative hyphae clamped, thin- to slightly thick-walled, hyaline, often branching near the septa, 3–5 μm wide, CB–. Skeletocystidia abundant, arising from sclerified generative hyphae in the subiculum and projecting into the trama and above the hymenium, clavate, usually tapering towards the apex, encrusted portion 25–60(–70) × 6–11 μm; thin-walled leptocystidia present on tips of the aculei, clavate to capitate, 4–7 μm wide. Basidia clavate, tetrasterigmate, 12–15 × 3.5–5 μm. Basidiospores ellipsoid, hyaline, thin-walled, IKI–, CB–, (4–)4.3–5.1(–5.3) × 2.4–3.0(–3.1) μm.
Habitat and distribution.
Known only from the type locality in Araucaria forests in Rio Grande do Sul State. Growing on dead logs of unidentified angiosperms.
Notes.
Cabalodontia albofulva is characterized by fragile basidiomes with thin aculei measuring 0.5–0.8 mm long and ellipsoid basidiospores measuring (4–)4.3–5.1(–5.3) × 2.4–3.0(–3.1) µm. Cabalodontia queletii (Bourdot & Galzin) Piątek is morphologically similar but differs by having thicker aculei and larger basidiospores, measuring 5–6 × 3–3.5 µm (Bernicchia and Gorjón 2010). Phylogenetically, C. albofulva is closely related to C. lincangense, from which it differs by approximately eight bp in the ITS sequences, four in the tef1-α coding region, and five in the rpb1 coding region. Morphologically, however, C. lincangense is distinct, displaying more widely spaced and longer aculei (2–4 per mm, 1–1.5 mm long) and shorter basidiospores measuring (3.5–)3.8–4.2(–4.5) × (2.3–)2.5–2.8 µm (Dong et al. 2024). Cabalodontia albofulva is currently known only from its type locality in southern Brazil. Despite extensive sampling in the surrounding region, including adjacent Araucaria forests, no additional specimens have been collected, suggesting that it could represent a rare taxon.
Cabalodontia
brunnea
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & Regio sp. nov.
D92BD90A-E142-5A88-AFB4-6AB82AD3C90D
861267
Etymology.
Refers to the brownish basidiomes.
Diagnosis.
Characterized by the pale brown basidiomes with short, irregular spines, hyaline to yellow hyphae, and basidiospores (3.7–)4–4.7(–4.9) × 2.4–3.0(–3.1) μm.
Typification.
Brazil. Rio Grande do Sul: Caxias do Sul, Cânion Palanquinhos, 18 Sep. 2017, M.C. Westphalen 600/17 (SP 512588).
Description.
Basidiomes adnate, resupinate, annual, not detaching, forming confluent large patches on the substrate, soft and somewhat brittle; sterile margins absent or very thin, fimbriate to byssoid, up to 1 mm wide. Hymenophore pale brown to ochraceous when fresh, unchanging or slightly darker upon drying, formed by small irregular spines, up to 0.5 mm long, often somewhat cluttered, very brittle upon drying, 6–9 per mm. Subiculum beige to pale yellowish brown, homogeneous, thin and fragile, cottony, up to 0.5 mm thick.
Hyphal system monomitic, hyphae more densely arranged towards the trama and looser near the substratum; generative hyphae clamped, thin- to thick-walled, often branching near the septa or at clamp connections, 2.5–5 μm wide, hyaline to pale yellow, golden yellowish-brown in mass, CB+. Skeletocystidia abundant, heavily encrusted, arising from sclerified generative hyphae in the subiculum and projecting toward the trama and above the hymenium, clavate or more commonly tapering towards the apex, encrusted portion 25–70(–100) × 5–9(–12) μm. Basidia clavate, tetrasterigmate, 13–16 × 4–5 μm. Basidiospores ellipsoid, hyaline, thin-walled, (3.7–)4–4.7(–4.9) × 2.4–3.0(–3.1) μm, IKI–, CB–.
Habitat and distribution.
Known only from the type locality in Araucaria forests in Rio Grande do Sul State. Growing on dead logs of unidentified angiosperms.
Notes.
Cabalodontia brunnea is distinguished within the genus by its pale brown basidiomes with irregular short spines and a somewhat farinaceous appearance due to the encrusted cystidia. Microscopically, it presents yellowish hyphae and ellipsoid basidiospores measuring (3.8–)4–4.7(–4.9) × 2.4–3.0(–3.1) μm. As in the case of C. albofulva, this species is known only from its type locality in southern Brazil. Despite extensive sampling in the region, including nearby Araucaria forests, no additional specimens have been found, suggesting that it could represent a rare taxon.
Phylogenetically, C. brunnea is closely related to C. delicata, a species also found in Araucaria forests and high-altitude Atlantic rainforest areas (Westphalen et al. 2019). However, C. delicata is relatively common in these habitats and differs from C. brunnea by its pale white to cream-colored basidiomes, shorter aculei, and smaller, subglobose to broadly ellipsoid basidiospores.
Cabalodontia
lincangense
Taxon classificationAnimaliaPolyporalesSteccherinaceae
(J.H. Dong & C.L. Zhao) Westphalen & Regio comb. nov.
468D4A42-6770-5C55-95F4-07B47008A6D5
861268
Basionym.
Steccherinum lincangense J.H. Dong & C.L. Zhao, Mycosphere 15 (1): 1252. 2024.
Notes.
Cabalodontia lincangense was originally described based on a phylogeny that did not adequately represent Cabalodontia and other Steccherinaceae lineages, resulting in its placement in Steccherinum. In our analysis, the species nests within Cabalodontia, sharing morphological features such as an odontioid hymenophore, a monomitic hyphal system with clamped hyphae, and skeletocystidia tapering toward the apex. It is closely related to C. albofulva but can be differentiated by more widely spaced and longer aculei (2–4 per mm, 1–1.5 mm long) and shorter basidiospores measuring (3.5–)3.8–4.2(–4.5) × (2.3–)2.5–2.8 µm (Dong et al. 2024).
Cabalodontia
tenuissima
Taxon classificationAnimaliaPolyporalesSteccherinaceae
(C.L. Zhao & Y.X. Wu) Westphalen & Regio comb. nov.
ABB50256-A31B-533B-B19C-5DB64668F5C7
861269
Basionym.
Steccherinum tenuissimum C.L. Zhao & Y.X. Wu, PLoS ONE 16 (1): e0244520, 7. 2021.
Notes.
Cabalodontia tenuissima was originally described based on a phylogeny that included only Steccherinum spp., where it nested in a separate clade with S. xanthum, a species shown by our analyses to be a synonym of C. albofibrillosa. Morphologically, C. tenuissima differs from all other confirmed Cabalodontia spp. by presenting a dimitic hyphal system, although it shares other features of the genus, such as fragile and thin basidiomes with an odontioid hymenophore (Wu et al. 2021a). It is possible that the skeletal hyphae reported by the authors represent undifferentiated lower portions of skeletocystidia, since the incrustations are restricted to the apical region. This interpretation is also supported by the drawings presented in the original description, where clamped hyphae are abundant and few thick-walled skeletal hyphae are shown, which is not common in truly dimitic species. Such thick-walled hyphal segments are also observed in other species of the genus, such as C. brunnea and C. albofulva, but they are scarce and, when carefully observed, give rise to cystidia. Further examination of the specimens is required to confirm this hypothesis. Nevertheless, because our phylogenetic analyses are well supported and include sequences from the type material (CLZhao 3153), we chose to transfer this species to Cabalodontia. Together with Irpex oreophilus (Lindsay & Gilb.) Niemelä, these are the only two dimitic taxa recovered within the genus.
Steccherinum
bononiae
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & Gugliotta sp. nov.
17B1C77B-D1D8-52CE-A66B-FD9B1EF5B93C
858851
Etymology.
In honor of Dr. Vera Bononi, for her contribution to the knowledge of Steccherinum in Brazil.
Diagnosis.
Distinguished by the combination of resupinate to effused-reflexed basidiomes with waxy spines upon drying, elongated skeletocystidia protruding above the hymenium, aculei measuring up to 0.75 mm, and basidiospores 2.6–3.6 × 1.8–2.5 µm.
Typification.
Brazil. São Paulo: São Luis do Paraitinga, Parque Estadual da Serra do Mar, Núcleo Santa Virgínia, Trilha Poço do Pito, 06 Jun. 2017, M.C. Westphalen 557/17 (SP 512686).
Description.
Basidiomes adnate, resupinate to effused-reflexed, easily separable from the substratum and usually detaching upon drying, membranaceous to papery and pliable when fresh, turning corky and somewhat waxy upon drying; pilei when present arising from effused parts of the basidiomes and at the margins, usually imbricate and narrow, up to 8 mm wide, pileus surface cream to beige, fimbriate, faintly zonate; sterile margins entire, smooth, pelliculose, up to 2 mm wide, white to cream. Hymenophore hydnoid, pale salmon to pale yellowish when fresh, turning beige to tan upon drying, aculei 0.25–0.75 × 0.1–0.25 mm, with acute apices, with a pilose appearance from the protruding cystidia, solitary or more rarely laterally fused, somewhat crowded, 6–9 per mm. Subiculum cream to beige, homogeneous, slightly dense, up to 0.8 mm thick.
Hyphal system dimitic; subicular hyphae compact, not agglutinated; aculei tramal hyphae intertwined, parallel; generative hyphae clamped, thin to slightly thick-walled, hyaline, occasionally branched, 2–4 µm wide, more abundant in the base of the subiculum; skeletal hyphae thick-walled to almost solid, hyaline to slightly yellowish, 2–4.5 µm wide. Skeletocystidia abundant, arising from the trama and protruding above the hymenium, somewhat elongated, covered with medium-sized crystals, (20–35–65 × 5–9(–10) µm; leptocystidia present, often scattered and more abundant at the apical portion of the aculei, clavate to fusoid, smooth to apically encrusted. Basidia clavate, tetrasterigmate, 10–14 × 4–5 µm. Basidiospores broadly ellipsoid, hyaline, thin-walled IKI–, CB–, 2.6–3.6 × 1.8–2.5 µm.
Mating system.
Tetrapolar. Monosporic cultures obtained from specimen NR71.
Habitat and distribution.
Known from southern, southeastern, and northeastern Brazil. Likely widespread in the Brazilian Atlantic Rainforest.
Specimens examined.
Brazil • Pernambuco: Olinda, 7 GAC - Batalhão do Exército, 16 Jun. 2018, R.S. Chikowski RC1625 (URM 93107). • Rio Grande do Sul: São Francisco de Paula, FLONA, 12 Mar. 2022, M.C. Westphalen 726/22 (ICN 213868); • Dom Pedro de Alcântara, RPPN Mata do Prof. Baptista, 17 Nov. 2022, N.C. Regio NR71 (ICN 213869). • São Paulo: São Paulo, Parque Estadual das Fontes do Ipiranga, 09 May 2014, A.M. Gugliotta 1615 (SP 512683); • Parque Estadual Cantareira, Núcleo Engordador, Trilha da Cachoeira, 24 Apr. 2017, M.C. Westphalen 547/17 (SP 512685); • Ribeirão Grande. Parque Estadual Intervales, Trilha roda d’água, 07 Jul. 2015, V. Motato-Vásquez MV446 (SP 512675).
Notes.
Basidiomes of S. bononiae exhibit considerable variation, ranging from completely effused to effused-reflexed, and aculei vary in size from 0.25 to 0.75 mm long. The basidiospore size and shape in this species resemble those of S. molle and S. resinaceum. However, S. molle can be distinguished by its softer, membranaceous basidiomes, slightly shorter spines, and wider skeletocystidia, which are more deeply embedded in the trama and covered with larger, chunky crystals. Steccherinum resinaceum, in turn, can be distinguished by its denser basidiomes, laterally fused aculei that are often bifurcated at the apices, and embedded cystidia covered with large crystals. Phylogenetically, S. bononiae forms a sister clade to S. elegantissimum, but the latter can be distinguished by its thinner, shorter aculei and larger basidiospores. Additionally, S. elegantissimum typically grows on thin branches, forming small concrescent patches, whereas S. bononiae usually forms basidiomes as a single patch. Both species share the presence of elongated and projecting cystidia covered with a somewhat organized cap of crystals, but these crystals are slightly larger in S. bononiae (Fig. 9).
Steccherinum
elegantissimum
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & R.M. Silveira sp. nov.
EA3F572F-5228-56E3-B8A2-7CAA67DB4127
858852
Etymology.
Refers to the delicate basidiomes with thin aculei and the elongated projecting cystidia.
Diagnosis.
Characterized by effused basidiome with small aculei (up to 0.5 mm long), a cottony subiculum, elongated and thin skeletocystidia protruding above the hymenium, and basidiospores with (3.4–)3.5–5.0(–5.1) × 2.2–2.9(–3.0) µm.
Typification.
Brazil. São Paulo: Ribeirão Grande. Parque Estadual Intervales, 28 Feb. 2018, M.C. Westphalen 633/18 (SP 512681).
Description.
Basidiomes adnate, effused, usually not detaching when dried, but easily separable if pulled from the substratum, formed by the coalescence of several small patches, membranaceous and pliable when fresh, turning papery and fragile after drying; sterile margins entire, byssoid, smooth to finely fimbriate, thin, up to 0.5 mm wide. Hymenophore hydnoid, cream to pale salmon when fresh, more or less unchanging or slightly duller upon drying, aculei 0.3–0.5 (0.55) × 0.1–0.2 mm, with acute apices, pilose from the protruding cystidia, solitary or rarely fused at the base, sub-distant, 5–8 per mm. Subiculum white, homogeneous, with a loose cottony and soft consistency, up to 0.3 mm thick.
Hyphal system dimitic, subicular hyphae loosely arranged, tramal hyphae intertwined and somewhat more densely arranged; generative hyphae clamped, thin to slightly thick-walled, hyaline, occasionally branched, 2–3 µm wide; skeletal hyphae thick-walled to almost solid, abundant throughout the basidiome, hyaline, 2–4 µm wide. Skeletocystidia abundant, arising from the trama and protruding above the hymenium, somewhat thin and elongated, covered with small crystals, 20–70(–75) × 4–8, but rarely above 6 µm wide; leptocystidia present, often scattered and more abundant at the apical portion of the aculei, clavate to fusoid, smooth. Basidia clavate, tetrasterigmate, 13–15 × 4.5–5 µm. Basidiospores ellipsoid to narrowly ellipsoid, hyaline, thin-walled, IKI–, CB–, (3.4–)3.5–5.0(–5.1) × 2.2–2.9(–3.0) µm.
Mating system.
Tetrapolar. Monosporic cultures obtained from specimens 720/21 and 721/21.
Habitat and distribution.
Known from Araucaria forest and Atlantic Rainforest areas in southern and southeastern Brazil.
Specimens examined.
Brazil • Rio Grande do Sul: Canela, FLONA, 22 Oct. 2021, M.C. Westphalen 720/21 and 721/21 (ICN 213870 and 213871).
Notes.
This species can be recognized by its small, sub-distant aculei and basidiomes formed by several concrescent small patches, typically growing on thin branches. Microscopically, the basidiospores are larger compared to other Neotropical Steccherinum species, measuring (3.4–)3.5–5.0(–5.1) × 2.2–2.9(–3.0) µm. Additionally, S. elegantissimum has the longest and thinnest cystidia in the group, covered by a layer of thin crystals (Figs 9B, 11L). The basidiomes with a cottony subiculum resemble those of S. perparvulum and S. molle, but the latter two species have smaller basidiospores and typically form larger patches that cover wider branches or logs.
Steccherinum
molle
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & Minosso sp. nov.
374EBD76-0E29-51DE-AD9A-CBEE1650DB00
858854
Etymology.
Refers to the soft and pliable consistency of the basidiomes.
Diagnosis.
Recognized by soft basidomes with a cottony subiculum, small aculei up to 0.7 mm long, skeletocystidia usually embedded in the trama and encrusted with large crystals, and basidiospores 2.5–3.5(–3.6) × 1.8–2.4(–2.5) µm.
Typification.
Brazil. Rio Grande do Sul: Dom Pedro de Alcântara, RPPN Mata do Prof. Baptista, 10. Sep. 2022, M.C. Westphalen 734/22 (ICN 213874).
Description.
Basidiomes adnate, resupinate, easily separable from the substratum, usually detaching upon drying, coalescing to form large patches, soft and membranaceous when fresh, unchanging to slightly papery upon drying, but remaining soft and pliable, easily tearing; sterile margins thin, cottony, smooth to finely floccose, up to 1.5 mm wide. Hymenophore hydnoid, cream to pale orange when fresh, unchanged upon drying, aculei (0.3–)0.4–0.5(–0.7) × 0.1–0.2(–0.25) mm, with acute apices, sometimes slightly asperulate from the protuding cystidia, solitary or more rarely fused at the base, sub-distant, 5–7 per mm. Subiculum white, homogeneous, loose, and cottony, very thin, up to 0.4 mm thick.
Hyphal system dimitic, subicular hyphae very loosely arranged, tramal hyphae intertwined and somewhat densely arranged, subparallel; generative hyphae clamped, thin to slightly thick-walled, hyaline, occasionally branched, 2–3 µm wide; skeletal hyphae thick-walled to almost solid, hyaline, 2–4 µm wide. Skeletocystidia abundant, immersed in the trama or protruding into the hymenium, coarsely encrusted with large crystals, (20–)30–60 × 7–13(–15); leptocystidia present, more commonly seen on the apices of the aculei, smooth to finely encrusted, mostly clavate. Basidia clavate, tetrasterigmate, 10–14 × 4–5 µm. Basidiospores ellipsoid, hyaline, thin-walled, IKI–, CB–, 2.5–3.5(–3.6) × 1.8–2.4(–2.5) µm.
Mating system.
Tetrapolar. Monosporic cultures obtained from specimen 734/22.
Habitat and distribution.
Known from Atlantic Rainforest areas in southeastern and northeastern Brazil and Araucaria Forests in southern Brazil. Mostly found in high-altitude regions above 700 m. Likely widespread throughout the Atlantic rainforest biome.
Specimens examined.
Brazil • Rio Grande do Sul: São Francisco de Paula, Hotel Parador Hampel, Trilha, 19 Jun. 2017, M.C. Westphalen 568/17 (SP 512692); • ibid., FLONA, 14 May 2018, M.C. Westphalen 641/18 (SP512689); • Farroupilha, Parque dos Pinheiros, 20 Apr. 2018, M.C. Westphalen 661/18 (SP 512690); • ibid., 27 Mar. 2019 M.C. Westphalen 687/19 (SP 512691); • ibid., 21 Jan. 2023 M.C. Westphalen 739/23 (ICN 213875). • São Paulo: Ribeirão Grande, Parque Estadual Intervales, Trilha da gruta, 07 Jul. 2015, V. Motato-Vásquez MV450 (SP 512672). • Sergipe: Areia Branca, Parque Nacional Serra de Itabaiana, 25 Jul. 2025, R.S. Souza RSS236 (URM).
Notes.
This species can be recognized mainly by the soft and pliable basidiomes with a cottony subiculum, usually forming large patches on the substratum, small aculei, and embedded cystidia encrusted with large crystals. Steccherinum resinaceum presents similar basidiospores and cystidia but can be distinguished by the harder, waxy basidiomes and laterally fused or bifurcate aculei. Steccherinum perparvulum is similar macroscopically but presents papery basidiomes upon drying, not soft or pliable, shorter basidiospores, and slightly thinner cystidia encrusted with smaller crystals.
Steccherinum
perparvulum
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Hjortstam & Ryvarden
0A01D71B-AC67-55D2-8EB9-CF34490005C2
Description.
Full description in: Hjortstam and Ryvarden (2008).
Mating system.
Tetrapolar. Monosporic cultures obtained from specimens 710/20 and 742/23.
Habitat and distribution.
Widespread in southern to southeastern Brazil, mainly in high-altitude areas.
Specimens examined.
Brazil • Rio Grande do Sul: Nova Roma do Sul, ponte velha, 05 Apr. 2017, M.C. Westphalen 524/17 (SP512658); • São Francisco de Paula, CPCN Pró-Mata, 17 May 2018, M.C. Westphalen 659/18 (SP 512661); • ibid., 22. Apr. 2023 M.C. Westphalen 744/23 (ICN 213878); • ibid., 19 Apr; 2024, N.C. Regio NR186 (ICN 213879); • ibid., FLONA, 19 Apr. 2023, M.C. Westphalen 742/23 (ICN 213877); • Caxias do Sul, Parque da Gruta Nossa Senhora de Lourdes, 28 Mar. 2019, M.C. Westphalen 692/19; ibid., 09. Nov. 2020, M.C. Westphalen 710/20 (ICN 213876). • São Paulo: Moji-Guaçu, Fazenda Campininha, 29–30 Jan 1987, D. Pegler, K. Hjortstam & L. Ryvarden 24589 (O - holotype); • Parelheiros, Parque Estadual da Serra do Mar, Núcleo Curucutu, 16 Nov. 2016, V. Motato-Vásquez MV815 (SP 512656); • São Paulo, Parque CIENTEC, 07 Jul. 2016, V. Motato-Vásquez MV728 (SP 512657); • ibid., Parque Estadual Cantareira, Núcleo Engordador, Trilha da Cachoeira, 24 Apr. 2017, M.C. Westphalen 543/17 (SP 512659); • Santo André, Reserva Biológica do Alto da Serra de Paranapiacaba, 25 Aug. 2017, M.C. Westphalen 592/17 (SP 512660).
Notes.
This species is primarily distinguished by having the smallest basidiospores (2.3–3.1 × 1.7–2.2 µm) of all Neotropical hydnoid/odontioid Steccherinum, usually measuring under 3 µm long (Table 5). Macroscopically, it typically forms large basidiomes with short aculei and a papery consistency when dried. While most of the specimens examined were strictly resupinate, some exhibited small pilei along the margins. We examined the holotype of S. perparvulum and found several specimens with basidiospore sizes and general morphological characteristics consistent with the species. However, our phylogenetic analyses revealed that these specimens comprise at least three distinct lineages (Figs 1, 3). These lineages show minor differences in the ITS region, although subtle variations also occur among specimens within the same lineage (Fig. 12).
ITS2 sequences in Steccherinum spp. showing interspecific and intraspecific variation.
To determine whether the genetic differences observed represent distinct biological species or reflect broader molecular variability within the ITS region of S. perparvulum, we conducted mating tests with monosporic cultures from three different specimens (Table 6). Two of these specimens belonged to the same lineage (710 and 744, lineage 1), whereas the third represented a different lineage (742, lineage 3). The mating tests showed positive results only between specimens of the same lineage, with clamp connections present in 11 of 12 monosporic pairings. In contrast, all confrontations with specimen 742 were negative, with only simple-septate hyphae present. This further supports the hypothesis that the observed molecular differences represent separate species within the group. Unfortunately, cultures from lineage 2 were not available for compatibility testing. Further studies incorporating additional monosporic cultures would be valuable to explore potential intercompatibility within the group, especially considering that lineage 2 is phylogenetically very close to lineage 1.
Table 6.: Mating tests in the Steccherinum perparvulum species complex.
While our studies corroborate at least two different species within S. perparvulum, no morphological or biogeographical evidence was found to support their segregation. Therefore, we chose to retain these taxa under the same name for the time being, treating it as a species complex. Considering their morphological and phylogenetic affinities, this approach aims to facilitate the taxonomy of the group rather than complicating it by increasing the number of species and adding morphologically indistinguishable taxa. In addition, at present, it is not possible to define which of the lineages represents S. perparvulum s.s. Sequences from the type specimen or the type locality could further elucidate this issue and help clarify the taxonomy of this species complex.
Steccherinum
resinaceum
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & Minosso sp. nov.
589A4DB4-6D95-5DF6-AA25-9B7FAD240669
858855
Etymology.
Refers to the hard and waxy consistency of the basidiomes when dried.
Diagnosis.
Distinguished from other species in the genus mainly by the waxy and dense basidiomes upon drying, the laterally fused aculei that bifurcate at apices, and the skeletocystidia encrusted with large crystals, usually embedded in the trama.
Typification.
Brazil. São Paulo: São Luís do Paraitinga, Parque Estadual da Serra do Mar, Núcleo Santa Virgínia, Trilha Pirapitinga, 05 Jun. 2017, M.C. Westphalen 551/17 (SP 512670).
Description.
Basidiomes adnate, resupinate, easily separable from the substratum and usually detaching upon drying, membranaceous to papery and pliable when fresh, turning waxy and somewhat rigid upon drying; sterile margins entire, smooth, pelliculose, up to 1 mm wide. Hymenophore hydnoid, cream to pale yellowish when fresh, turning beige to ochraceous upon drying, aculei (0.3–)0.4–0.75 × 0.15–0.3(–0.5) mm, usually with straight to bifurcate apices with a pilose appearance from the protruding cystidia, solitary or more commonly laterally fused, somewhat crowded, 5–6 per mm. Subiculum cream to beige, homogeneous, dense, up to 0.3 mm thick.
Hyphal system dimitic, subicular hyphae very compact, tramal hyphae intertwined, subparallel; generative hyphae clamped, thin to slightly thick-walled, hyaline, occasionally branched, 2–4 µm wide, more abundant in the base of the subiculum; skeletal hyphae thick-walled to almost solid, hyaline to slightly yellowish, 2–4.5 µm wide. Skeletocystidia abundant, immersed in the trama or protruding into the hymenium, coarsely encrusted with large crystals, 25–45(–50) × (7–)9–11(–12) µm, some thinner and longer cystidia also observed immersed in the subiculum, up to 60 µm long and 6–8 µm wide; leptocystidia present, abundant in the apices of the aculei, clavate, ventricose, or capitate, smooth or with a crown of crystals. Basidia clavate, tetrasterigmate, 10–12 × 3.5–5 µm. Basidiospores broadly ellipsoid, hyaline, thin-walled IKI–, CB–, 2.7–3.4(–3.5) × 1.7–2.4 µm.
Habitat and distribution.
Known only from Atlantic Rainforest areas in southeastern Brazil.
Specimens examined.
Brazil • São Paulo: São Paulo, Parque Estadual Cantareira, Núcleo Engordador, Trilha da Cachoeira, 24 Apr. 2017, M.C. Westphalen 540/17 (SP 512671); • São Luís do Paraitinga, Parque Estadual da Serra do Mar, Núcleo Santa Virgínia, Trilha Olho d’ água, 13 Feb. 2019 M.C. Westphalen 665/19 (SP 512687, ICN 213880); • São Paulo, Parque Estadual das Fontes do Ipiranga, 18 Feb. 2019, M.C. Westphalen 679/19 (SP 512669).
Notes.
Steccherinum resinaceum is characterized by its waxy and somewhat hard basidiomes when dried, a hymenophore composed of laterally fused, often bifurcating aculei, and coarsely encrusted cystidia with large crystals. Macroscopically, the basidiomes of S. bononiae are somewhat similar but differ in having solitary aculei with a pilose appearance due to prominently protruding skeletocystidia, whereas in S. resinaceum the skeletocystidia are visible only at the apices of the aculei (Fig. 8E). Additionally, the cystidia in S. bononiae are longer and typically covered with small- to medium-sized crystals.
Steccherinum molle shares similar cystidial morphology and basidiospore size with S. resinaceum. Nonetheless, it can be distinguished by its softer basidiomes with a cottony subiculum and slightly shorter and thinner aculei. Unfortunately, we could not obtain monosporic cultures of S. resinaceum to confirm its mating system.
Steccherinum
subochraceum
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Bononi & Hjortstam
E4650FA5-EAB7-5498-B996-6BF1F05720C9
Description.
Full description in Hjortstam and Bononi (1986).
Mating system.
Tetrapolar. Monosporic cultures obtained from specimen 730/22.
Habitat and distribution.
Known from the Atlantic Rainforest in southern and southeastern Brazil.
Specimens examined.
Brazil • Rio Grande do Sul: São Salvador, Montenegro, 04 Apr. 1945, Rick s.n. (PACA 22824 - holotype of Iprex hydenus Rick); • São Francisco de Paula, FLONA, 12 Mar. 2022, M.C. Westphalen 730/22 (ICN 213881); • ibid., 25 Nov. 2023, M.C. Westphalen 748/23 (ICN 213883); • Farroupilha, Parque dos Pinheiros, 22 Jul. 2023, M.C. Westphalen 746/23 (ICN 213882); • Canela, FLONA, 13 Jul. 2024, Westphalen 761/24 (ICN 313884). • São Paulo: São Paulo, Parque Estadual das Fontes do Ipiranga, 06 Oct. 1966, H. Requejo s.n. (SP 97591 - holotype).
Notes.
S. subochraceum was described from southeastern Brazil to provide a valid name for Iprex hydenus, originally described by Rick (1959) but considered invalid due to the lack of a designated type specimen. Among Neotropical hydnoid Steccherinum species, S. subochraceum is readily distinguished by the effused-reflexed basidiomes with notably long aculei (Figs 8F, 10A), usually measuring over 1.5 mm long (0.85–2.25 × 0.15–0.45 mm) (Table 4). Microscopically, it presents thin, elongated cystidia [25–60(–70) × 5–8(–10) µm] immersed in the trama, sometimes protruding at the apices of the aculei, and subglobose basidiospores measuring (3.2–)3.3–4.2 × 2.5–3.8 µm. Steccherinum larssonii and Steccherinum basibadium share a similar basidiospore size range (3.5–4.5 × 3.0–3.5 µm and 3.6–4.5 × 3.1–3.4 µm, respectively) but can be distinguished by their significantly shorter aculei, which reach a maximum length of 0.75 mm. In addition, Steccherinum basibadium forms more developed pilei with a brownish surface, whereas S. larssonii often produces completely resupinate basidiomes or only small pilei at the margins (Maas Geesteranus 1974; Westphalen et al. 2021).
The subicular generative hyphae of S. subochraceum are regularly clamped, although simple-septate hyphae were observed in the trama of some specimens. Simple-septate hyphae were readily observed in specimen 746/23 but were less common in 730/22. Phylogenetically, S. subochraceum forms a sister clade to S. undigerum, a Neotropical species with somewhat similar basidiome morphology (effused-reflexed with imbricate pilei) and that also exhibits a strongly fimbriate mycelium in culture. However, S. undigerum differs in having a poroid hymenophore with dentate dissepiments and slightly larger basidiospores (4–5 × 3.5–4.5 μm).
Steccherinum
undulatum
Taxon classificationAnimaliaPolyporalesSteccherinaceae
Westphalen & R.M. Silveira sp. nov.
82022089-43AC-52D2-8067-11C6C4DAE67C
858853
Etymology.
Refers to the basidiomes with small, wavy pilei.
Diagnosis.
Recognized mainly by the combination of basidiomes formed by several small, wavy, imbricate pilei, aculei with 0.75–1 mm long, and ellipsoid basidiospores 3.3–4.1 × 2.0–2.7(–2.8) µm.
Typification.
Brazil. Rio Grande do Sul: São Francisco de Paula, CPCN Pró-Mata, 20 Apr. 2023, M. C. Westphalen 743/23 (ICN 213872).
Description.
Basidiomes adnate, concrescent, effused-reflexed, with several small, wavy, imbricate pilei, formed by the coalescence of several small pilei with conjoined reflexed bases, usually not detaching when dried, but easily separable if pulled from the substratum, somewhat membranaceous and pliable when fresh, turning papery to corky upon drying and somewhat waxy; pilei small, up to 4 mm wide and 9 mm in length, pilear surface cream to beige, tomentose, sulcate, and sometimes faintly zonate; sterile margins entire, somewhat cottony, smooth to finely fimbriate, up to 1.5 mm wide. Hymenophore hydnoid, at first orange in young and fresh specimens, then turning pale salmon to beige with age and upon drying, aculei 0.75–1.0 × 0.2–0.4 mm, with acute to round apices, sometimes slightly asperulate from the protruding cystidia, solitary or more rarely laterally fused, crowded, 4–6 per mm. Subiculum white, homogeneous, up to 0.6 mm thick.
Hyphal system dimitic, subicular hyphae compact, not agglutinated, tramal hyphae parallel, intertwined, and densely arranged; generative hyphae clamped, thin to slightly thick-walled, hyaline, occasionally branched, 2–4 µm wide; skeletal hyphae thick-walled to almost solid, abundant throughout the basidiome, hyaline, 2–5 µm wide. Skeletocystidia abundant, immersed in the trama or with the apices protruding into the hymenium, covered with a thick cap of small crystals, 20–60 × 7–10 µm; leptocystidia present, often scattered and somewhat inconspicuous, mostly clavate or with a rounded apex, smooth. Basidia clavate, tetrasterigmate, 12–16 × 4–5 µm. Basidiospores ellipsoid, hyaline, thin-walled, IKI-, CB-, 3.3–4.1 × 2.0–2.7(–2.8) µm.
Habitat and distribution.
Known only from Araucaria forests in southern Brazil.
Specimens examined.
Brazil • Rio Grande do Sul: Canela, FLONA, 13 Jul. 2024, M.C. Westphalen 760/24 (ICN 213873).
Notes.
This species can be distinguished by its basidiomes consisting of small, wavy, imbricate pilei (Fig. 10B). One of the collections studied (760/24) exhibited a bright orange coloration when fresh, whereas the other (743/23) displayed a more faded salmon hue. This suggests that only young, fresh specimens display brighter colors, which gradually turn paler as they mature or dry. The aculei of S. undulatum are the second largest in the group, being smaller only than those of S. subochraceum. Microscopically, the species also features slightly larger basidiospores when compared to S. molle, S. resinaceum, and S. bononiae, but smaller than those of S. elegantissimum.
Although surveys for Steccherinum species have been ongoing since 2017, only two specimens of S. undulatum have been found, both in Araucaria forests in southern Brazil, approximately 45 km apart. This suggests that it is likely rarer than other species in the group, which have been more frequently collected and are more widely distributed.
Identification key to Brazilian corticioid species of Steccherinum and Cabalodontia
**: **
Discussion
Through the integration of morphological, molecular, and culture data, this study expands the knowledge of Steccherinum in the Neotropics by describing five new species and providing newly obtained molecular data on S. perparvulum and S. subochraceum. Along with S. larssonii and S. basibadium, these seven species share many morphological traits and can only be reliably distinguished through a combination of characteristics, such as aculei size, basidiospore size and shape, and the incrustation and position of cystidia. However, considerable overlap in these features among taxa makes species identification challenging without a thorough comparison of multiple specimens. Furthermore, molecular data revealed that S. perparvulum represents a species complex comprising at least three distinct lineages that are morphologically indistinguishable and exhibit no clear differences in distribution, with some lineages occurring sympatrically. In addition to the new data on Steccherinum, two new species and two new combinations are proposed in Cabalodontia, doubling the number of known taxa in the genus.
Morphology and ecology of Cabalodontia spp.
The genus Cabalodontia differs from Steccherinum mainly by its fragile and brittle basidiomes, a monomitic hyphal system, and cystidia tapering toward the apex. However, some clavate cystidia were also observed in the analyzed specimens, resembling those found in Steccherinum s.s. In addition, two taxa recovered within Cabalodontia, C. tenuissima and Irpex oreophilus, have been described as dimitic. The placement of I. oreophilus within Cabalodontia was discussed by Westphalen et al. (2021), who noted that the absence of sequence data from the type specimen or type locality prevents confirmation of its generic placement. Therefore, a conservative approach is followed here, and no new combination is proposed for this species at present.
Interpretation of the hyphal system in Cabalodontia may be challenging in some cases due to the presence of thick-walled hyphae that give rise to cystidia. While some authors may interpret these segments as skeletal hyphae, all examined Cabalodontia specimens show abundant generative hyphae with clamp connections. This contrasts with Steccherinum, where the hyphal system is clearly dimitic, with abundant skeletal hyphae. A similar pattern is evident in the original description of C. tenuissima (Wu et al. 2021a), in which the line drawings depict predominantly clamped hyphae and only a few sclerified hyphae in the subiculum. Type studies of I. oreophilus, in turn, revealed more sclerified hyphae compared to other species in the group; however, thick-walled clamped hyphae and abundant generative hyphae were also observed. For these reasons, the hyphal system in Cabalodontia is best interpreted as ranging from monomitic to pseudo-dimitic, which also helps distinguish the genus from truly dimitic taxa in Steccherinum and Junghuhnia. If future studies confirm truly dimitic taxa within the genus, its circumscription would need to be expanded.
The three Brazilian species of Cabalodontia currently known (C. albofulva, C. brunnea, and C. delicata) were collected in high-altitude areas above 800 m, mostly in Araucaria forests. This distribution suggests a preference for subtropical conditions in the Neotropics, particularly given the absence of records from northern Brazil. While C. delicata is relatively frequent in high-altitude forests, C. brunnea and C. albofulva appear to be rare and are each known from a single collection, despite several field expeditions conducted in nearby regions over the past seven years.
Morphology and ecology of Steccherinum spp.
The hymenophore morphology is a key feature to distinguish taxa in Steccherinum because of the variation in size and number of aculei per millimeter. In addition, the aculei may be solitary or fused, their apices acute, straight, rounded, or bifurcate, and pilose or not due to protruding skeletocystidia. These features, along with the general basidiome consistency when dried and the arrangement of the subicular hyphae, should be carefully examined for species identification. Although basidiospores are often of significant importance in differentiating closely related taxa, among the five new species described, only S. elegantissimum exhibits distinctly larger spores, yet still presents some overlap, especially with S. undulatum. All the remaining species have very small and similar basidiospores, which do not represent a reliable taxonomic feature to distinguish them. Nevertheless, basidiospore size and shape remain important features to consider when examining Neotropical Steccherinum spp., particularly when assessed alongside other morphological characteristics.
Regarding the cystidia, two main types of incrustations have been observed in Steccherinum spp.: (1) skeletocystidia covered with small to medium crystals and (2) coarsely encrusted cystidia with large crystals (Fig. 11K, L). In addition, while in some species the cystidia are long and project above the hymenium, in others they are embedded in the trama or have only slightly projecting apices. In contrast, cystidia in Cabalodontia are typically widened at the base, tapering toward the apex, and encrusted with large crystals (Fig. 11J). It is important to note, however, that variation in cystidial incrustation, shape, and position may occur in some cases, and these features should be considered as additional characteristics when assessing their overall appearance across different species, which can help differentiate them.
Regarding the ecology of the studied taxa, most Steccherinum species were found growing on fallen branches of angiosperms, suggesting that they occupy similar ecological niches and perform comparable ecological roles. The only exception was S. larssonii, which was collected both from fallen branches and logs of unidentified angiosperms. In addition, no clear biogeographic pattern was observed, as several species occurred within the same or nearby regions. Only S. larssonii, S. resinaceum, and S. undulatum exhibited more restricted distributions, with S. larssonii and S. resinaceum being found exclusively in southeastern Brazil and S. undulatum in montane Araucaria forests in the southern region. Nevertheless, it is highly probable that all of the species treated in this study are widespread throughout the Atlantic Rainforest biome, with some, such as S. undulatum, potentially restricted to montane environments. Broader sampling across different forest types and elevations would help clarify species ranges and their ecological preferences.
Culture studies data
The culture studies revealed distinct patterns among the analyzed taxa. While S. bononiae and S. elegantissimum exhibited slower growth rates, S. perparvulum and S. resinaceum showed comparatively faster growth. At the same time, notable intraspecific variation was observed in the growth rates of S. undulatum and S. subochraceum, highlighting variability within species. Further comparisons were made between cultures from two of the three S. perparvulum lineages; however, no significant differences in growth rates or morphological traits were detected (Table 3, Fig. 6). These findings suggest that, although growth patterns and mycelial macromorphology can be useful for taxonomic identification in Steccherinum, intraspecific variation and overlapping characteristics must be considered. To achieve more accurate results, broader sampling of cultures is necessary. Expanding the dataset could help identify more consistent patterns and refine criteria that can be used to aid in distinguishing taxa.
Through monosporic confrontations, we were able to test the mating system of five of the Steccherinum species studied (S. bononiae, S. elegantissimum, S. molle, S. perparvulum, and S. subochraceum), confirming that all are tetrapolar, a characteristic also seen in other species of the genus (Rajchenberg 2011; Westphalen et al. 2018). Additional mating tests were also conducted among monosporic cultures of the different taxa to assess potential sexual compatibility. All matings were negative, confirming that the taxa are reproductively isolated and represent distinct biological species.
Molecular data and divergence time estimates
Comparative analysis of the ITS sequences among the studied species revealed distinct regions, particularly at the beginning of ITS2, that are unique and can be used to reliably differentiate taxa within the group (Fig. 12). Although some variation was also detected in the ITS region of the S. perparvulum complex, it is less pronounced than the differences observed among the other species. For example, in the highlighted ITS2 region, variation is limited to approximately seven scattered base changes. In contrast, comparison between the sister species S. elegantissimum and S. bononiae reveals 13 base substitutions, comprising longer segments with more pronounced differences (Fig. 12). It is noteworthy that, in spite of the genetic variation observed in ITS as well as tef1-α, the species within the group exhibit a high degree of morphological similarity. This pattern could suggest that speciation in these taxa may be relatively recent, particularly given their shared ecological niches and overlapping distributions. Regarding the S. perparvulum clade, the three lineages found are estimated to have been segregated for at least 10.39 Myr (95% height = 15.73–5.46 Myr), indicating that morphological differentiation between taxa can take more than 10 million years to occur. To elucidate this, the group would benefit from further analyses encompassing the biogeographic history and diversification rates of its taxa.
Recently, Deng et al. (2025) provided the first analysis of divergence time estimates of the Steccherinaceae, encompassing 16 different genera. In turn, our study provides more comprehensive sampling with 19 genera and several new sequences of Neotropical taxa, especially of Steccherinum and Cabalodontia. The five-gene dataset utilized to obtain the MCC tree (Fig. 3) represents the most comprehensive phylogram constructed for the family to date, excluding only monotypic or poorly sequenced genera from the analysis. In this sense, the low support obtained in some of the nodes that form the backbone of Steccherinaceae likely reflects the scarcity of molecular data and the existence of yet undescribed taxa within the family. It is noteworthy that the same clades tend to shift according to the dataset applied, something that may only be resolved through increased sampling and phylogenomic analyses of the group.
Regarding the molecular dating, our analysis retrieved Steccherinaceae with a mean crown age of 86.4 Myr, with an HPD value that is consistent with the 113 Myr estimation obtained by He et al. (2024) through maximum likelihood analysis of phylogenomic data. Although slightly older, it also corroborates the mean crown age of 120.8 Myr obtained by Deng et al. (2025), placing the origin of the family in the early to late Cretaceous period. The differences between the values are probably the result of differences in the datasets and genes selected or the slightly different methodologies applied, especially the inclusion of a long-distance fossil calibration point representing the segregation of Ascomycota and Basidiomycota (Deng et al. 2025). In addition, the estimation indicates that most taxa within Steccherinaceae have emerged in the Cenozoic Era, between the Paleocene and Oligocene, with an overall younger age compared to genera of other families in Polyporales, such as Irpicaceae, Meripilaceae, and Meruliaceae (Wang et al. 2022; Li et al. 2025a; Wang et al. 2025). This more recent speciation event could explain the species complexes and morphological similarities shared among related taxa, especially in hydnoid-poroid genera such as Steccherinum, Cabalodontia, and Junghuhnia.
Considering that the majority of the known species in Steccherinaceae are wood decomposers, their evolutionary history likely follows the same path as taxa within the previously mentioned families, tracking the angiosperm radiation and expansion that started in the Late Cretaceous and presented a diversification surge in the Cenozoic Era, marked by global climatic cooling that could explain this shift (Varga et al. 2019; Zuntini et al. 2024). Nevertheless, the Neotropical distribution and speciation of the family, especially for Steccherinum spp., could be intrinsically related to the biogeographic history of the Atlantic Rainforest, from which most of the known species were described. In this sense, the Araucaria forests, present in the southern portion of the biome for at least 200 Myr, possess a complex biogeographic and evolutionary history that could be linked to the speciation processes observed in the group (Vasconcellos et al. 2024).
Although the divergence estimates presented here provide valuable insights into the evolutionary history of the group, the estimated ages of most genera should be interpreted with caution until additional molecular data allow for more comprehensive analyses encompassing the genetic diversity of Steccherinaceae and its taxa. Therefore, continued efforts to expand the molecular and phylogenomic data of the group will be essential to clarify the evolutionary relationships among its genera.
Additional neotropical Steccherinum s.l. taxa
In addition to the species discussed in this study, two other odontioid/hydnoid Steccherinum species have been described in the Neotropics: S. filiferum (Yurchenko et al. 2023) and S. diversum (Hjortstam 1999). We examined images of the S. diversum type specimen, kindly sent by the Kew Herbarium staff, and found that it differs from all the studied species by presenting basidiomes that are strongly attached to the substrate, a loose subiculum, and brittle aculei. These morphological traits deviate from typical Steccherinum s.s. This is particularly relevant given that several species originally described in Steccherinum have been shown, through phylogenetic analyses, to belong to other genera (Miettinen et al. 2012; Miettinen and Ryvarden 2016; Westphalen et al. 2021). Furthermore, in its original description, Hjortstam (1999) considered S. diversum morphologically similar to S. laeticolor (Berk. & M.A. Curtis) Banker, a species that is phylogenetically related to Junghuhnia rather than Steccherinum. Future studies incorporating new collections and molecular data may help determine if S. diversum should remain in Steccherinum or be transferred to another genus. Unfortunately, we were unable to examine specimens of S. filiferum. However, the species can be readily distinguished from other Neotropical odontioid Steccherinum by its fragile basidiomes, monomitic hyphal system, and simple-septate generative hyphae. Phylogenetically, it nests in a clade alongside S. amapaense, S. fragile, and S. laxum (Fig. 1), all of which share somewhat soft or fragile basidiomes, a monomitic to pseudo-dimitic hyphal system, and simple-septate generative hyphae. These morphological traits deviate from Steccherinum s.s., which is characterized by a dimitic hyphal system and clamped generative hyphae. This suggests that these taxa could be transferred to a new genus, provided that no existing older name is available for the group. However, S. rubigimaculatum also presents simple-septate generative hyphae but differs by having a dimitic hyphal system, and it falls into a separate, distantly related lineage. As previously mentioned, many of the outer clades in our molecular analysis showed low support, especially regarding the basal groups in Steccherinum s.l. Interestingly, S. tenue nested near the simple-septate/monomitic species but with low support, despite possessing a dimitic hyphal system and clamped generative hyphae. Additionally, previous studies (Miettinen et al. 2012; Westphalen et al. 2021) have placed S. tenue in different clades within Steccherinum, suggesting that the currently available molecular data are insufficient to resolve its true phylogenetic position or to strongly support the transfer of S. amapaense, S. fragile, S. filiferum, and S. laxum to a different genus.
In conclusion, this study represents a significant advancement in the understanding of the genus Steccherinum, providing precise morphological data, the addition of over 110 new DNA sequences from five distinct regions, mating system determination of five species, mycelial culture data, and divergence time estimates. However, it is important to highlight that several recently described species assigned to Steccherinum are not phylogenetically nested within the genus (unpublished data), requiring further studies to determine their correct generic placement. Additionally, other taxa not included in the present study due to insufficient morphological or sequence data may represent additional species within the genus in the Neotropical region, underscoring the need for continued research on the Steccherinaceae to accurately assess the group’s true diversity in the region.
Supplementary Material
XML Treatment for Cabalodontia albofulva
XML Treatment for Cabalodontia brunnea
XML Treatment for Cabalodontia lincangense
XML Treatment for Cabalodontia tenuissima
XML Treatment for Steccherinum bononiae
XML Treatment for Steccherinum elegantissimum
XML Treatment for Steccherinum molle
XML Treatment for Steccherinum perparvulum
XML Treatment for Steccherinum resinaceum
XML Treatment for Steccherinum subochraceum
XML Treatment for Steccherinum undulatum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bernicchia A, Gorjón SP (2010) Corticiaceae s.l. Fungi Europaei n° 12. Italy, Candusso.
- 2Binder M, Hibbett DS, Wang Z et al. (2006) Evolutionary relationships of Mycaureola dilseae (Agaricales), a basidiomycete pathogen of a subtidal rhodophyte. American Journal of Botany 93(4): 547–556. 10.3732/ajb.93.4.54721646215 · doi ↗ · pubmed ↗
- 3Binder M, Larsson KH, Matheny PB et al. (2010) Amylocorticiales ord. nov. and Jaapiales ord. nov.: Early diverging clades of Agaricomycetidae dominated by corticioid forms. Mycologia 102(4): 865–880. 10.3852/09-28820648753 · doi ↗ · pubmed ↗
- 4Bouckaert R, Vaughan TG, Barido-Sottani J et al. (2019) BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. P Lo S Computational Biology 15(4): e 1006650. 10.1371/journal.pcbi.1006650 PMC 647282730958812 · doi ↗ · pubmed ↗
- 5Cao T, Yu JR, Nguyễn TTT et al. (2021) Multiple-marker phylogeny and morphological evidence reveal two new species in Steccherinaceae (Polyporales, Basidiomycota) from Asia. Myco Keys 78: 169–186. 10.3897/mycokeys.78.57823 PMC 804173433883969 · doi ↗ · pubmed ↗
- 6Cohen K, Finney S, Gibbard P et al. (2013) [updated] The ICS International Chronostratigraphic Chart. Episodes 36: 199–204. 10.18814/epiiugs/2013/v 36i 3/002 · doi ↗
- 7Darriba D, Taboada GL, Doallo R et al. (2012) j Model Test 2: More models, new heuristics and parallel computing. Nature Methods 9(8): 772. 10.1038/nmeth.2109 PMC 459475622847109 · doi ↗ · pubmed ↗
- 8Deng Y, Chen M, Wang K et al. (2025) Morphological and phylogenetic analyses reveal one new genus and six new species in Irpicaceae and Steccherinaceae (Polyporales, Basidiomycota) from the Yunnan–Guizhou Plateau, Asia. IMA Fungus 16: e 172367. 10.3897/imafungus.16.172367 PMC 1277099941503134 · doi ↗ · pubmed ↗