Taxonomic and phylogenetic analyses reveal two new Comoclathris taxa (Dothideomycetes, Pleosporaceae) from Inner Mongolia, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2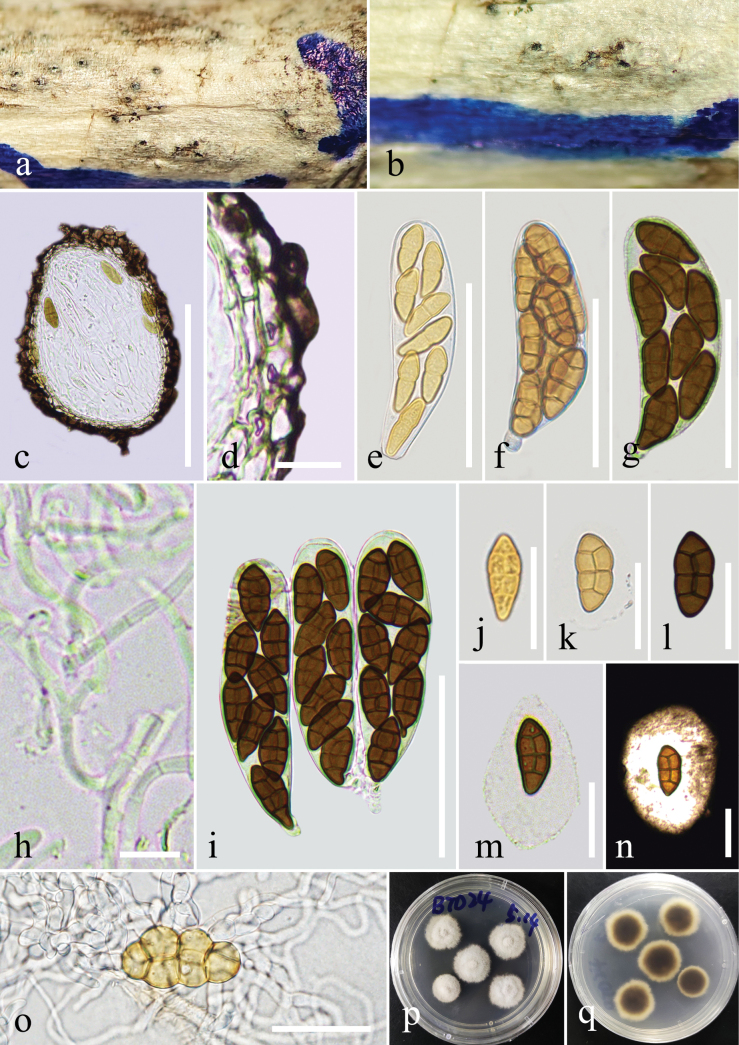 Figure 3
Figure 3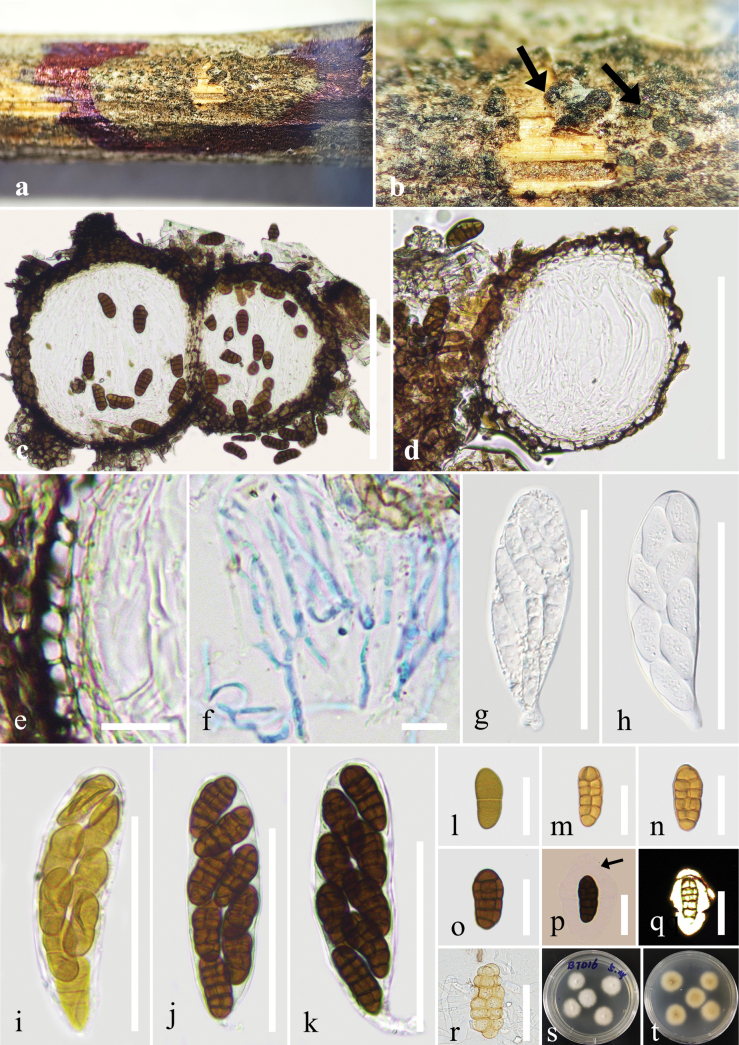 Figure 4
Figure 4| Taxon name | Strain number |
|
| SSU | References | |
|---|---|---|---|---|---|---|
|
| MCC9771T |
|
| — | — |
|
|
| MN3-2019 |
|
| — | — |
|
|
| CBS 366.52 |
|
| — |
|
|
|
| WA0000074564T |
|
| — | — |
|
|
| MFLUCC 15-0465T |
|
|
|
|
|
|
| MFLUCC 15-0476 |
|
|
| — |
|
|
| CCMJ 13076T |
|
|
|
|
|
|
| CCMJ 13077 |
|
|
|
|
|
|
| CBS 156.53 | — |
|
|
|
|
|
| CBS 157.53 | — |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| MFLU 20-0391T |
|
|
|
|
|
|
| MFLU 20-0397T |
|
|
|
|
|
|
| MFLU 20-0399 |
|
|
| — |
|
|
| MFLUCC 18-0773T |
|
|
| — |
|
|
| CBS 467.76 | — |
|
|
|
|
|
| CH-16 |
|
| — | — |
|
|
| MFLUCC 15-0073T |
| — | — | — |
|
|
| MFLUCC 14-0968T |
|
|
| — |
|
|
| MFLUCC 14-0561 |
|
|
| — |
|
|
| MFLU 20-0385T |
|
|
|
|
|
|
| MFLU 18-1236 |
|
|
|
|
|
|
| GMBCC2306T |
|
|
|
| Unpublished |
|
| GMBCC2319 |
|
|
|
| Unpublished |
|
| MFLUCC 14-0974 |
|
|
| — |
|
|
| MFLUCC 14-1159T |
|
|
| — |
|
|
| MFLU 15-0203T |
|
|
|
|
|
|
| MFLU 16-0234 |
|
|
|
|
|
|
| MFLUCC 14-0962T |
|
|
|
|
|
|
| MFLU 16-0229T |
|
|
|
|
|
|
| MFLUCC 13-0763T |
|
|
| — |
|
|
| MFLUCC 13-0817 |
|
|
| — |
|
|
| MFLUCC 13-0214T |
|
|
| — |
|
|
| CBS 602.72 |
|
| — | — |
|
|
| CCMJ 13078T |
|
|
|
|
|
|
| CCMJ 13079 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CBS 523.66 |
|
|
|
|
|
|
| CBS 246.79 |
|
|
|
|
|
| Species | Ascospore | Hosts | Collection locations | References | ||
|---|---|---|---|---|---|---|
| Color | Septa | Sheath | ||||
|
| Dark brown | 3–8 transverse septa and 3–5 vertical septa | N/A |
| India |
|
|
| Yellow to pale brown | 6–8 transverse septa | N/A | Soil sample | Antarctica |
|
|
| Mid to dark reddish-brown | 6 transverse septa and 1 vertical septum | With a sheath 2–3 μm wide | Canada |
| |
|
| Yellow to pale brown | 4 transverse septa and 2–3 vertical septa | Surrounded by a thick mucilaginous sheath |
| Italy |
|
|
| N/A | N/A | N/A |
| India |
|
|
| N/A | N/A | N/A |
| N/A |
|
|
| Brown | 3 transverse septa and 1 vertical septum | Surrounded by a thick mucilaginous sheath | China |
| |
|
| Dark reddish-brown | 3 transverse septa and 1 vertical septum | With a uniform sheath 2–3 μm wide | USA |
| |
|
| Mid yellowish-brown | 3 transverse septa and 1 vertical septum | With a sheath 2–3 μm wide | USA |
| |
|
| Dark yellowish-brown | 4 transverse septa and 1 vertical septum | With a sheath 1.5–4 μm wide |
| India |
|
|
| Brown | 7 transverse septa and 1 vertical septum | Surrounded by amucilaginous sheath |
| Italy |
|
|
| Dark reddish-brown | 4 transverse septa and 1 vertical septum | With a sheath 2–3 μm wide |
| India |
|
|
| Dark brown | 6 transverse septa and 1–2 vertical septa | Mucilaginous sheath | Italy |
| |
|
| Brown or pale brown | 2–4 transverse septa and 1–2 vertical septa | Without sheath |
| Russia, Ukraine |
|
|
| Mid reddish-brown | 7 transverse septa and 1 vertical septum | With a sheath 1–2 μm wide | Sweden |
| |
|
| N/A | N/A | N/A |
| Greece, Italy |
|
|
| N/A | N/A | N/A |
| India |
|
|
| Brown | 6–8 transverse septa and 1–2 vertical septa | Mucilaginous sheath |
| Italy |
|
|
| Brown to reddish-brown | 4–5 transverse septa and 1–2 vertical septa | Surrounded by a distinct, hyaline, mucilaginous 3–8 μm wide sheath |
| USA |
|
|
| yellowish-brown | 3–5 transverse septa and 1–2 vertical septa | With a thick mucilaginous sheath | Italy |
| |
|
| yellowish-brown | 3–5 transverse septa and 1–2 vertical septa | With a thick mucilaginous sheath | Italy |
| |
|
| Dark reddish-brown | 3 transverse septa and 1 vertical septum | With a sheath 2–4 μm wide | USA |
| |
|
| N/A | N/A | N/A |
| Greece |
|
|
| Mid reddish-brown | 4 transverse septa and 1 vertical septum | With a uniform sheath 3–4 μm wide |
| Iran |
|
|
| Yellow to light brown | 3 transverse septa and 2 vertical septa | Surrounded by a thick, hyaline, a mucilaginous sheath |
| Russia |
|
|
| Dark reddish-brown | 5 transverse septa and 1 vertical septum | With a uniform sheath, 1–2 μm wide |
| USA |
|
|
| N/A | N/A | N/A |
| N/A |
|
|
| N/A | N/A | N/A | Italy |
| |
|
| Mid reddish-brown | 4 transverse septa and 1 vertical septum | With a uniform sheath 1–2 μm wide | Canada |
| |
|
| Pale brown | 4–7 transverse septa and 1–2 vertical septa | Surrounded by a thick mucilaginous sheath |
| Italy |
|
|
| Brown | 6–7 transverse septa and 2–4 vertical septa | Surrounded by a thick mucilaginous sheath |
| Italy |
|
|
| Pale brown | 5–7 transverse septa and 1 vertical septum | Surrounded by a thick mucilaginous sheath |
| Italy |
|
|
| Mid yellowish-brown | 4 transverse septa and 1 vertical septum | With a uniform sheath 2–3 μm wide | USA |
| |
|
| Brown to reddish-brown | 4–5 transverse septa and 1–2 vertical septa | Surrounded by a distinct, hyaline, mucilaginous 5–9 μm wide sheath | Italy |
| |
|
| Mid yellowish-brown | 3 transverse septa and 1 vertical septum | N/A | USA |
| |
|
| Dark reddish-brown | 3 transverse septa and 1 vertical septum | N/A |
| Turkey |
|
|
| Yellow to pale brown | Muriform | Surrounded by a mucilaginous sheath |
| Italy |
|
|
| N/A | N/A | N/A |
| N/A |
|
|
| Brown | 3 transverse septa and 1 vertical septum | N/A | Austria, Denmark, England, Germany, India, Iran, Netherlands, Pakistan, Poland, Portugal, Russia, Switzerland, and USA | ||
|
| Dark reddish-brown | 3 transverse septa and 1 vertical septum | With a sheath 1–3 μm wide | USA |
| |
|
| Dark reddish-brown | 3 transverse septa and 1 vertical septum | With a uniform sheath 2–3 μm wide |
| USA |
|
|
| Mid reddish-brown | 5 transverse septa and 1 vertical septum | With a uniform sheath 1.5–3.5 μm wide | Finland, Norway, Switzerland |
| |
|
| N/A | N/A | N/A |
| India |
|
|
| Brown to dark brown | 3 transverse septa and 1 vertical septum | Surrounded by a thick mucilaginous sheath |
| China |
|
|
| Mid reddish-brown | 3 transverse septa and 1 vertical septum | N/A |
| Argentina |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Biocrusts and Microbial Ecology
Introduction
Comoclathris Clem. is a genus of ascomycetous fungi belonging to Pleosporaceae Nitschke (Pleosporales, Dothideomycetes). The genus was established by Clements (1909), with C. lanata Clem. designated as the type species. Previously, the genus Comoclathris was placed in Diademaceae Shoemaker & C.E. Babc. (Shoemaker and Babcock 1992), but recent morphological and molecular phylogenetic analyses have supported its placement in Pleosporaceae (Zhang et al. 2012; Ariyawansa et al. 2015; Hyde et al. 2024). Currently, 52 epithets of Comoclathris are listed in Index Fungorum (2026), with molecular data available for only 21 species in NCBI. This genus is characterized by ascomata with circular lid-like openings, cylindrical to clavate asci, and muriform ascospores that are reddish-brown to dark brown and applanate (Mattoo et al. 2023). While most Comoclathris species are mainly saprobic on woody substrates, dead stems, and leaves of various plants or soil (Wanasinghe et al. 2015; Thambugala et al. 2017; Brahmanage et al. 2020; Crous et al. 2021), some are endophytes (González-Menéndez et al. 2018; Mattoo et al. 2023) and phytopathogens (Moral et al. 2017). Comoclathris is a widely distributed genus, found in many countries and regions, as detailed in recent publications (Mattoo et al. 2023; Xu et al. 2024). Comoclathris was first reported in China by Xu et al. (2024), who described two novel species from Jilin and Yunnan provinces.
The Inner Mongolia Autonomous Region is located in northern China and lies within a temperate semi-arid zone, with ecological types covering typical steppe, meadow steppe, desert steppe, as well as sandy land and forest-steppe transition zones (Han et al. 2009; Su et al. 2011; Yan et al. 2025). The vegetation of this region is dominated by herbaceous plants of the Asteraceae Bercht. & J.Presl, Fabaceae Lindl., and Poaceae (R. Brown) Barnhart families, and although steppe vegetation predominates, the types are relatively diverse rather than strictly monotypic (Fu et al. 2021; Wang et al. 2024). Despite this lack of vegetation diversity, fungi are important in these regions, as they may harbour new species with significant bioprospecting potential. Recent studies have shown that the fungal communities in Inner Mongolia exhibit considerable diversity. For example, analyses from 63 root samples during a study of ectomycorrhizal (EM) fungi revealed 288 operational taxonomic units (OTUs) spanning 31 lineages, indicating that fungal communities in forest/forest-edge environments may be much more diverse than anticipated (Wang et al. 2021). Further studies on arbuscular mycorrhizal (AM) fungi in grassland environments conducted by Wang et al. (2020) revealed 24 species spanning across eight genera. Despite increasing research on fungal diversity in Inner Mongolia Autonomous Region, detailed data and systematic taxonomic studies on saprobic fungi remain scarce. To date, only a few saprobic fungal novelties have been reported from Inner Mongolia, for example, Diatrype betulaceicola Z.E. Yang and Hai X. Ma, with polysporous asci described by Yang et al. (2022). The unique ecological conditions of the semi-arid grassland ecosystem in this region make saprobic fungi crucial in decomposing plant debris, cycling nutrients, and maintaining soil fertility. Therefore, studying the taxonomy of saprobic fungi is essential to unravel their hidden fungal diversity and potential applications.
Our ongoing studies in this region explore the diversity of saprobic ascomycetes inhabiting dead plant material in the desert regions of Inner Mongolia, specifically the Xiangshawan Scenic Area. During a survey of microfungi from decaying substrates, two species of Comoclathris were collected and examined. Morphological characteristics and phylogenetic analyses based on multi-locus sequence data (ITS, LSU, SSU, and rpb2) were employed to confirm their taxonomic placement within Pleosporaceae and support their establishment as novel species. This study provides new insights into the saprobic fungal diversity of northern China and expands the known distribution and species richness of the genus Comoclathris.
Materials and methods
Sample collection, morphological observation, single-spore isolation and preservation
During a fungal resource survey in the desert areas of Inner Mongolia Autonomous Region, China (Fig. 1), dead fallen branches and stems with fungal fruiting bodies were collected. Important information was recorded during the collection process (collector, date, location, and habitat) (Rathnayaka et al. 2024). After that, the samples were placed in dry plastic bags for further observation and experimentation in the laboratory. Morphological structures were examined by using an OPTEC SZ650 dissecting stereomicroscope (Chongqing, China), and the microstructures of fungi were observed and photographed by using an OLYMPUS optical microscope (Tokyo, Japan) with an OLYMPUS DP74 (Tokyo, Japan) digital camera. Micro-morphological structures were measured in the Tarosoft ® Image Framework program v. 1.3. The photo plates were edited in Adobe Photoshop CS3 Extended version 22.0.0 software (Adobe Systems, California, USA).
Vegetation and habitat in the desert area of Xiangshawan Scenic Area.
Fungi were isolated and cultured using the single-spore isolation method described by Senanayake et al. (2020). The experiment was conducted under a sterile environment. After the work surface was disinfected with 75% alcohol, an alcohol lamp was lit. First, fresh fruiting bodies were observed and selected, then cut open with a sterile blade. Spores were picked up with a sterile needle, placed in sterile water and dispersed, and finally transferred to potato dextrose agar (PDA) plates (100 mL/plate) using a pipette. From 6 hours after inoculation to 48 hours, spores were checked promptly after germination with a sterile needle and then placed on a new PDA plate at 23–28 °C. Four germinated spores were transferred to each plate. After colonies formed, the morphology was observed to ensure consistency and confirm the results of the single-spore isolation experiment. The colonies were purified and used for molecular experiments, observed for sporulation, and photographed.
Specimens were deposited at Guizhou Medical University (GMB-W), China. Living cultures are deposited in the Guizhou Medical University Culture Collection (GMBCC), China. Facesoffungi (FoF) numbers were assigned as described by Jayasiri et al. (2015), and MycoBank (MB) numbers were assigned as outlined at https://www.mycobank.org/.
DNA extraction, PCR amplification, and sequencing
Total genomic DNA was extracted from one-month-old fresh fungal mycelium (grown on PDA). The DNA Extraction Kit-BSC14S1 (BioFlux, Hangzhou, P.R. China) was used following the manufacturer’s instructions. Polymerase chain reactions (PCR) were carried out using the following primers: The internal transcribed spacer (ITS) region was amplified with the primers ITS4 and ITS5 (White et al. 1990), 28S nrRNA gene (LSU) was amplified by using the primers LR0R and LR5 (Vilgalys and Hester 1990), 18S ribosomal RNA (SSU) was amplified using the primers NS1 and NS4 (White et al. 1990), and the partial RNA polymerase II subunit (rpb2) region was amplified with primers fRPB2-5F and fRPB2-7cR (Liu et al. 1999). The PCR thermal cycle programs for ITS, LSU, and SSU were as follows: an initialization step of 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, an annealing step at 55 °C for 50 s, an elongation step at 72 °C for 1 min and a final extension step of 72 °C for 10 min; and the PCR thermal cycle programs for rpb2 was as follows: an initialization step of 95 °C for 3 min, followed by 40 cycles of 95 °C for 50 s, an annealing step at 57 °C for 50 s, an elongation step at 72 °C for 90 s and a final extension step of 72 °C for 10 min (Du et al. 2025). The DNA amplification procedure was performed by PCR in a 25 μL reaction containing 12.5 μL 2xMaster Mix (mixture of Easy Taq TM DNA Polymerase, dNTPs, and optimized buffer (Beijing Trans Gen Biotech Co., Chaoyang District, Beijing, China)), 8.5 μL ddH_2_O, 2 μL of DNA template, and 1 μL of each forward and reverse primer (10 pM). Purification and sequencing of PCR products were carried out by Sangon Biotech Co., Kunming, China.
Phylogenetic analyses
A combined dataset of ITS, LSU, SSU, and rpb2 was used for the phylogenetic analyses. The quality of the raw sequences was checked in BioEdit v.7.2.6.1 (Hall 1999), and the forward and reverse sequences were spliced with Geneious 9.1.8 (Kearse et al. 2012). Sequences obtained from this study were searched in the GenBank database (http://blast.ncbi.nlm.nih.gov/) using BLAST to identify the closely related taxa of our strains. The additional sequences included in the analyses were collected from previous publications (Mattoo et al. 2023; Xu et al. 2024) and downloaded from GenBank (Benson et al. 2013). Phylogenetic analyses were carried out with 41 sequences (Table 1). The FASTA file was used for constructing the Randomized Accelerated Maximum Likelihood (RAxML) and Bayesian Inference analyses (BI) was performed using the OFPT (Zeng et al. 2023) with the protocol. Then, the FASTA file was converted to PHYLIP and NEXUS formats in ALTER for RAxML and BI phylogenetic analyses, respectively (Glez-Peña et al. 2010).
The CIPRES Science Gateway platform was used to carry out the RAxML and BI analyses (Miller et al. 2010). The RAxML tree, generated with 1,000 bootstrap replicates, was analyzed using RAxML-HPC2 on XSEDE (8.2.12) (Stamatakis et al. 2008; Stamatakis 2014) with the GTR+I+G model of evolution and bootstrap support. The BI tree was performed with MrBayes on XSEDE (3.2.7a) (Ronquist et al. 2012) by the Markov Chain Monte Carlo (MCMC) method to evaluate posterior probabilities (BYPP) (Richard and Lippmann 1991; Rannala and Yang 1996; Zhaxybayeva and Gogarten 2002). The best-fit nucleotide substitution models for each dataset were then selected using the Bayesian information criterion (BIC) from 22 common DNA substitution models with rate heterogeneity, as implemented in ModelFinder (Kalyaanamoorthy et al. 2017). The best model for ITS was TIM2e+I+G4, K2P+I for LSU, TPM2u+F+I for SSU, and TN+F+G4 for rpb2. Six simultaneous Markov chains were run for 2,000,000 generations, and a tree was sampled every 100^th^ generation. The phylogenetic tree was visualized in FigTree v.1.4.2 (Rambaut 2012) and edited in Microsoft PowerPoint 2021 and Adobe Photoshop CS3 Extended version 22.0.0 (Adobe Systems, California, USA). All newly generated sequences in this study were deposited in GenBank (https://www.ncbi.nlm.nih.gov/WebSub/?form=history&tool=genbank).
Results
Phylogenetic analyses
The phylogenetic trees obtained from RAxML and Bayesian Inference analyses were essentially similar. The RAxML analysis of the combined dataset yielded the best-scoring tree (Fig. 2), which consisted of 41 taxa, and the final alignment comprised 4029 characters, including gaps (ITS: 1–540, LSU: 541–1922, rpb2: 1923–3083, SSU: 3084–4029). The final ML optimization likelihood value was -14494.444982. The matrix contained 935 distinct alignment patterns, with 34.05% of characters undetermined or missing. Parameters for the GTR+I+G model of the combined ITS, LSU, rpb2, and SSU were as follows: estimated base frequencies A = 0.254158, C = 0.228977, G = 0.268187, T = 0.248678; substitution rates AC = 1.928002, AG = 4.366339, AT = 1.437220, CG = 1.029313, CT = 7.549462, GT = 1.0; proportion of invariable sites I = 0.641653; and gamma distribution shape parameter α = 0.725266. The final RAxML tree is shown in Fig. 2.
Maximum likelihood consensus tree inferred from the combined ITS, LSU, rpb2, and SSU multiple sequence alignments. Bootstrap support values for maximum likelihood (ML, first value) equal to or greater than 60% and Bayesian posterior probabilities from MCMC analyses (BYPP, second value) equal to or greater than 0.95 are given near the nodes. The scale bar indicates expected changes per site. The tree is rooted to Neocamarosporium betae (CBS 523.66) and N. calvescens (CBS 246.79). The new isolates are indicated in red, and the ex-type strains are in bold.
In our phylogenetic analyses, we obtained results consistent with those of recent publications by Mattoo et al. (2023) and Xu et al. (2024). Our new species, Comoclathris desertica (GMBCC2307 and GMBCC2320), was well separated from C. clematidis R. Xu, Phukhams. & Yu Li (CCMJ 13076 and CCMJ 13077) and C. xanthoceratis R. Xu, Phukhams. & Yu Li (CCMJ 13078 and CCMJ 13079) in a distinct lineage with 100% ML/1.00 PP statistical support; Comoclathris xiangshawanensis (GMBCC2305 and GMBCC2318) was well separated from C. antarctica Istel, J. Pawłowska & Wrzosek (WA0000074564, ex-type) with 87% ML/0.98 PP statistical support.
Taxonomy
Comoclathris
desertica
Taxon classificationFungiPleosporalesDiademaceae
X.G. Tian, T.Y. Du & D.F. Bao sp. nov.
74F9D643-66EA-5A91-8FAD-FF66B536FE4B
MB861793
Facesoffungi Number: FoF19021
Etymology.
Named after the desert habitat from where the holotype was collected.
Holotype.
GMB-W1518.
Description.
Saprobic on dead branches of an unidentified wood. Sexual morph: Ascomata 90–200 × 70–190 μm (x̄ = 115 × 116 μm, n = 5), solitary, scattered or aggregated, immersed, not obvious on the surface of the host, subglobose, black, ostiole not obvious. Peridium 10–30 μm wide, comprising thin-walled cells of textura angularis, dark brown to black. Hamathecium comprising numerous, 2 μm wide, filamentous, septate, branched, cellular pseudoparaphyses, hyaline, embedded in a gelatinous matrix, extending above the asci. Asci 65–85 × 18–23 μm (x̄ = 76 × 20 μm, n = 30), bitunicate, fissitunicate, 8-spored, cylindrical-clavate, short pedicellate, apically rounded. Ascospores 16–23 × 8–10 μm (x̄ = 19 × 9 μm, n = 30), 1-seriate, slightly overlapping, fusiform, initially yellowish, becoming brown to dark brown, muriform, with 3 transversely septa and 1 vertical septum, consisting of 6 cells, upper end conical, lower end tapered gradually, apical cell mostly undivided, straight or slightly curved, smooth-walled, surrounded by a mucilaginous sheath. Asexual morph: Undetermined.
Comoclathris desertica (GMB-W1518, holotype). a, b. Appearance of ascomata on the substrate; c. Section through an ascoma; d. Peridium; e-g, i. Asci; h. Pseudoparaphyses; j–n. Ascospores (m, n surrounded by mucilaginous sheath); o. A germinated ascospore; p, q. Colonies on PDA from above and below after one week. Scale bars: 100 µm (c); 10 µm (d, h); 50 µm (e-g, i); 20 µm (j–o).
Culture characteristics.
Ascospores germinated on PDA within 12 h at 28 °C, and germ tubes were produced around spores. Colonies on PDA reached 2 cm diam. after one week at 28 °C. Colonies obverse: circular, white-cream to grey, with sparsely hairy edge; grey from below, the active hyphae at the edge are white.
Material examined.
China • Inner Mongolia Autonomous Region, Ordos City, Xiangshawan, on dead branches of an unidentified wood in the desert, 01 October 2023, J.Y. Zhang, BTD24 (GMB-W1518, holotype), ex-type, GMBCC2307, other living culture, GMBCC2320.
GenBank numbers.
GMBCC2307: ITS = PX660512, LSU = PX660516, SSU = PX660508, rpb2 = PX672979; GMBCC2320: ITS = PX660513, LSU = PX660517, SSU = PX660509, rpb2 = PX672980.
Notes.
Comoclathris desertica clustered with C. clematidis (CCMJ 13076, ex-type and CCMJ 13077) and C. xanthoceratis (CCMJ 13078, ex-type and CCMJ 13079) in the phylogenetic tree with 100% in ML and 1.00 in BYPP statistical support (Fig. 2). A nucleotide base pair differences between our new strain (GMBCC2307, ex-type) and C. clematidis (CCMJ 13076, ex-type) in ITS, LSU, SSU, and rpb2 (without gaps) showed a 2.70% (14/519 bp) difference in ITS, 1.33% (9/679 bp) difference in LSU, 0.32% (2/628 bp) difference in SSU, and 5.58% (47/842 bp) difference in rpb2. Between our new strain (GMBCC2307, ex-type) and C. xanthoceratis (CCMJ 13078, ex-type), there was a 2.35% (12/510 bp) difference in ITS, 1.18% (8/676 bp) difference in LSU, 0.32% (2/628 bp) difference in SSU, and 6% (52/866 bp) difference in rpb2.
Morphologically, C. desertica is similar to C. clematidis and C. xanthoceratis, both of which have ascospores with 3 transverse septa and a vertical septum, surrounded by a thick mucilaginous sheath. But C. desertica is different from C. clematidis and C. xanthoceratis in having immersed ascomata, and dark brown ascospores, 1-seriate, slightly overlapping, smooth-walled; while C. clematidis has immersed to erumpent ascomata, and brown ascospores, with verrucose or echinulate wall (Xu et al. 2024); while C. xanthoceratis has immersed to semi-immersed ascomata, and brown to dark brown ascospores, 1–2-seriate (Xu et al. 2024). And our collection displays comparatively smaller ascospores than these two species (viz., C. desertica: 19 × 9 μm, C. clematidis: 30 × 14 μm, C. xanthoceratis: 37 × 16 μm).
Therefore, C. desertica is introduced as a new species from China based on morphological and molecular evidence as recommended by Jeewon and Hyde (2016).
Comoclathris
xiangshawanensis
Taxon classificationFungiPleosporalesDiademaceae
X.G. Tian, T.Y. Du & D.F. Bao sp. nov.
E45D5D8A-CF79-5DB2-A8EB-EBFFC06FD720
MB861794
Facesoffungi Number: FoF19022
Etymology.
Named after the type locality, “the desert region of Xiangshawan, China”.
Holotype.
GMB-W1526.
Description.
Saprobic on dead branches of an unidentified wood. Sexual morph: Ascomata 100–200 × 90–200 μm (x̄ = 132 × 118 μm, n = 10), solitary, scattered or aggregated into small groups, semi-immersed, subglobose, black, ostiole not obvious. Peridium 10–20 μm wide, comprising thin-walled cells of textura angularis, dark brown to black. Hamathecium comprising numerous, 2 μm wide, filamentous, septate, branched, cellular pseudoparaphyses, hyaline, embedded in a gelatinous matrix, extending above the asci. Asci 65–85 × 15–23 μm (x̄ = 73 × 21 μm, n = 30), bitunicate, fissitunicate, 8-spored, cylindrical-clavate, short pedicellate, apically rounded. Ascospores 17–23 × 7–11 μm (x̄ = 19 × 8.5 μm, n = 30), 1–2-seriate, broadly fusiform, initially yellowish, becoming brown, muriform, with 5(–6) transversely septa and 1–2 vertical septa, consisting of 9–14 cells, with conical or obtuse ends, straight or slightly curved, smooth-walled, surrounded by a mucilaginous sheath. Asexual morph: Undetermined.
Comoclathris xiangshawanensis (GMB-W1526, holotype). a, b. Appearance of ascomata on the substrate (arrows indicate black ascomata); c, d. Section through ascomata; e. Peridium; f. Pseudoparaphyses; g–k. Asci; l–q. Ascospores (arrow indicates mucilaginous sheath); r. A germinated ascospore; s, t. Colonies on PDA from above and below after one week. Scale bars: 100 µm (c, d); 10 µm (e, f); 50 µm (g–k); 20 µm (l–r).
Culture characteristics.
Ascospores germinated on PDA within 12 h at 28 °C and germ tubes were produced around spores. Colonies on PDA reached 2 cm diam. after one week at 28 °C. Colonies obverse: circular, white-cream, with sparsely hairy edge; cream to light yellow from below.
Material examined.
China • Inner Mongolia Autonomous Region, Ordos City, Xiangshawan, on dead branches of an unidentified wood in the desert, 01 October 2023, J.Y. Zhang, BTD16 (GMB-W1526, holotype), ex-type, GMBCC2305, other living culture, GMBCC2318.
GenBank numbers.
GMBCC2305: ITS = PX660510, LSU = PX660514, SSU = PX660506, rpb2 = PX672977; GMBCC2318: ITS = PX660511, LSU = PX660515, SSU = PX660507, rpb2 = PX672978.
Notes.
Comoclathris xiangshawanensis clustered with C. antarctica (WA0000074564, ex-type) and C. acuminata Mattoo, Nonzom & A. Ghosh (MCC9771, ex-type and MN3-2019) in the phylogenetic tree (Fig. 2). A comparison of nucleotide base pair differences between our new strain (GMBCC2305, ex-type) and C. antarctica (WA0000074564, ex-type) in ITS and LSU, the result (without gaps) showed a 1.65% (8/484 bp) difference in ITS, 0.24% (2/818 bp) difference in LSU. Between our new strain (GMBCC2305, ex-type) and C. acuminata (MCC9771, ex-type) in ITS and LSU, the result (without gaps) showed a 1.86% (9/484 bp) difference in ITS, 0.25% (2/807 bp) difference in LSU (C. antarctica (WA0000074564, ex-type) and C. acuminata (MCC9771, ex-type) have no available SSU and rpb2 sequences in NCBI and hence cannot be compared).
Morphologically, C. xiangshawanensis differs from C. antarctica and C. acuminata in having brown ascospores with 5(–6) transversely septa and 1–2 vertical septa, consisting of 9–14 cells, surrounded by a mucilaginous sheath; while C. antarctica has ascospores with 6–8 transvers septa, consisting of 10–17 cells (Crous et al. 2021); while C. acuminata has dark brown ascospores, with 3–8 transverse septa, 3–5 vertical septa, consisting of 15–21 cells, with a beak-like extension (Mattoo et al. 2023). In addition, our collection is saprobic on dead branches in desert areas in China, while C. antarctica is isolated from soil sample in Antarctica, and C. acuminata was isolated as an endophyte from the stem of Ephedra gerardiana Wall. in India (Mattoo et al. 2023).
Therefore, C. xiangshawanensis is introduced as a new species from China based on morphological and molecular evidence. To provide a better overview of the different morphologies associated with different Comoclathris species, we also compiled ascospore morphology and other ecological data in a table (Table 2).
Discussion
Comoclathris is a widely distributed genus, recorded on all five continents viz. Asia, Europe, Africa, the Americas, and Antarctica (Mattoo et al. 2023; Xu et al. 2024). Recent studies in China and other regions have shown that Comoclathris species are distributed in a variety of environments and substrates, from humid forests to semi-arid deserts (Mattoo et al. 2023; Xu et al. 2024; this study), suggesting that their diversity and ecological adaptability may be underestimated. The discovery of C. desertica and C. xiangshawanensis in desert wood in Inner Mongolia further expands the known distribution of this genus, providing valuable data for the global taxonomic system of Comoclathris. Furthermore, the thick, gelatinous sheaths enclosing the ascospores of these two species highlight the adaptability to extremely arid environments of these species. This is in line with the general characteristics of this genus, with obvious mucilaginous sheaths surrounding the ascospores. Based on morphological information currently available for this genus (Table 2), most species have mucilaginous sheaths, with only seven species having no record of mucilaginous sheaths (viz., C. acuminata, C. antarctica, C. galatellae D. Pem, Bulgakov & K.D. Hyde, C. sisyrinchii (Speg.) Shoemaker & C.E. Babc., C. sororia (Bubák) Shoemaker & C.E. Babc., C. typhicola (Cooke) H.A. Ariy. & K.D. Hyde, and C. xerophila (Speg.) Shoemaker & C.E. Babc.) (Cooke 1872; Shoemaker and Babcock 1992; Hongsanan et al. 2020; Crous et al. 2021; Mattoo et al. 2023).
Although 52 epithets are listed in Index Fungorum (2026), only 21 species have molecular data in NCBI, and the naming of many historical species is based solely on morphological characteristics. The lack of DNA sequences, type specimen revisions, and living cultures hinder phylogenetic analysis and species delineation. Therefore, future work should collect more specimens and supplement them with molecular data of extant species to clarify species boundaries and evolutionary relationships.
Inner Mongolia is a typical semi-arid region where long-term grazing and desertification have led to the degradation of grasslands and soils. Fungi play a crucial role in the decomposition of organic matter and nutrient cycling in fragile ecosystems, but the diversity of saprobic fungi remains poorly understood. The discovery of two new Comoclathris species in the Xiangshawan desert demonstrates that even degraded or arid habitats can foster rich and unique fungal communities. These fungal groups reflect the potential ecological resilience and adaptive evolution of microfungi under extreme temperature and humidity fluctuations. This study not only enriches the known fungal diversity in northern China but also highlights the importance of ongoing mycological surveys in arid grassland and desert ecosystems for taxonomic progress and ecological conservation.
Supplementary Material
XML Treatment for Comoclathris desertica
XML Treatment for Comoclathris xiangshawanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmadpour A, Ghosta Y, Alavi F, Alavi Z, Heidarian Z (2024) Comoclathris typhicola, a new species for the funga of Iran. Mycologia Iranica 11(1): 111–116. 10.3390/jof 11030225 · doi ↗
- 2Ariyawansa HA, Phookamsak R, Tibpromma S, Kang JC, Hyde KD (2014) A molecular and morphological reassessment of Diademaceae. The Scientific World Journal 675348: 1–11. 10.1155/2014/675348 PMC 391351124526916 · doi ↗ · pubmed ↗
- 3Ariyawansa HA, Thambugala KM, Manamgoda DS, Jayawardena RS, Camporesi E, Boonmee S, Wanasinghe DN, Phookamsak R, Hongsanan S, Singtripop C, Chukeatirot E, Kang JC, Jones EBG, Hyde KD (2015) Towards a natural classification and backbone tree for Pleosporaceae. Fungal Diversity 71: 85–139. 10.1007/s 13225-015-0323-z · doi ↗
- 4Aveskamp MM, Verkley GJ, de Gruyter J, Murace MA, Perelló A, Woudenberg JH, Groenewald JZ, Crous PW (2009) DNA phylogeny reveals polyphyly of Phoma section Peyronellaea and multiple taxonomic novelties. Mycologia 101(3): 363–382. 10.3852/08-19919537209 · doi ↗ · pubmed ↗
- 5Aveskamp MM, Gruyter JD, Woudenberg JHC, Verkley GJM, Crous PW (2010) Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Studies in Mycology 65(65): 1–60. 10.3114/sim.2010.65.01PMC 283621020502538 · doi ↗ · pubmed ↗
- 6Benson DA, Cavanaugh M, Clark K, Karsch-Mizrachi I, Lipman DJ, Ostell J, Sayers EW (2013) Gen Bank. Nucleic Acids Research 41(D 1): D 36–D 42. 10.1093/nar/gks 1195 PMC 353119023193287 · doi ↗ · pubmed ↗
- 7Boerema GH, De Gruyter J, Noordeloos ME, Hamers MEC (2004) Phoma identification manual: differentiation of specific and infra-specific taxa in culture. CABI Publishing. 10.1079/9780851997438.0000 · doi ↗
- 8Brahmanage RS, Dayarathne MC, Wanasinghe DN, Thambugala KM, Jeewon R, Chethana KT, Samarakoon MC, Tennakoon DS, De Silva NI, Camporesi E, Raza M (2020) Taxonomic novelties of saprobic Pleosporales from selected dicotyledons and grasses. Mycosphere: Journal of Fungal Biology 11(1): 2481–2541. 10.5943/mycosphere/11/1/15 · doi ↗