Two new species of the genus Conocybe (Agaricales, Bolbitiaceae) from Punjab, Pakistan

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
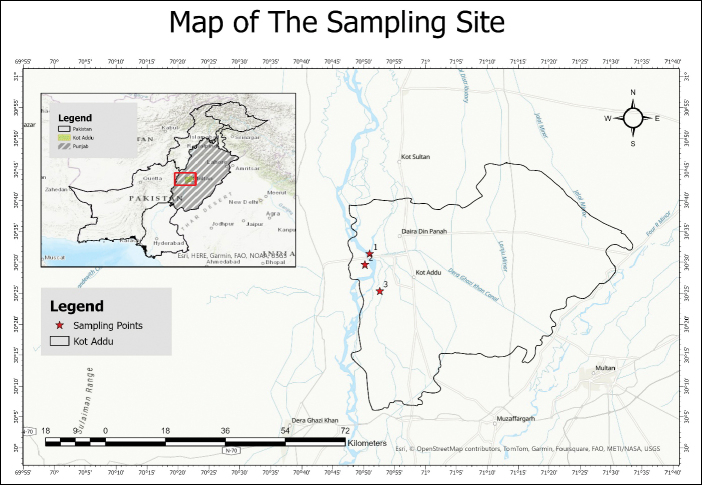 Figure 1
Figure 1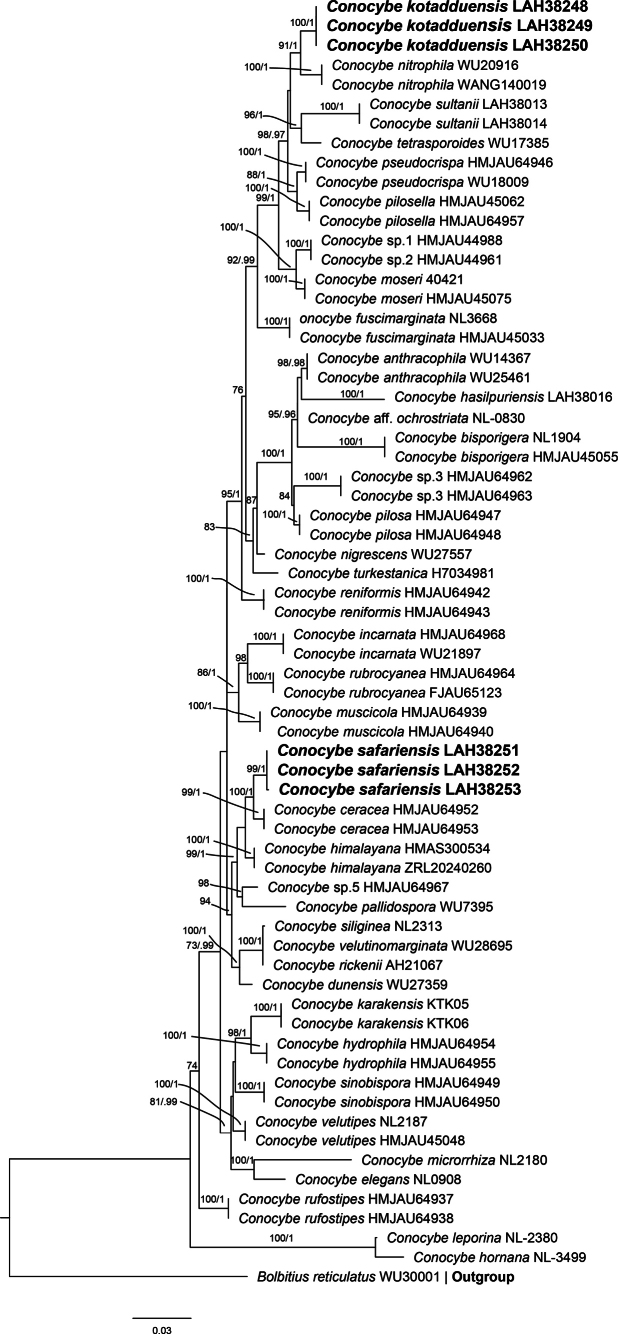 Figure 2
Figure 2 Figure 3
Figure 3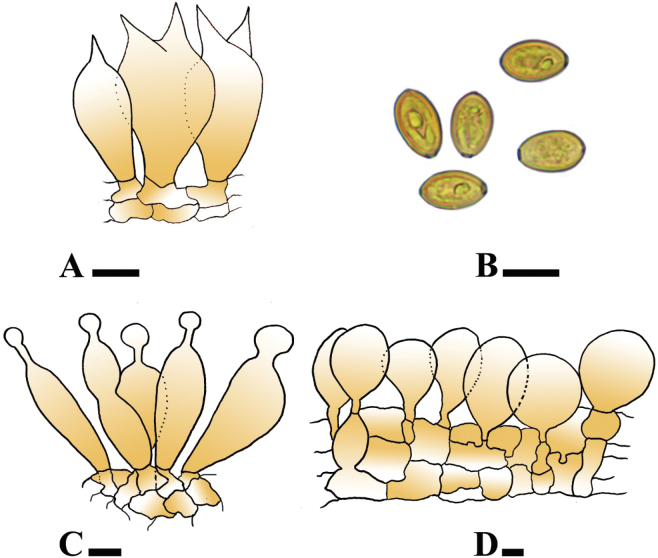 Figure 4
Figure 4 Figure 5
Figure 5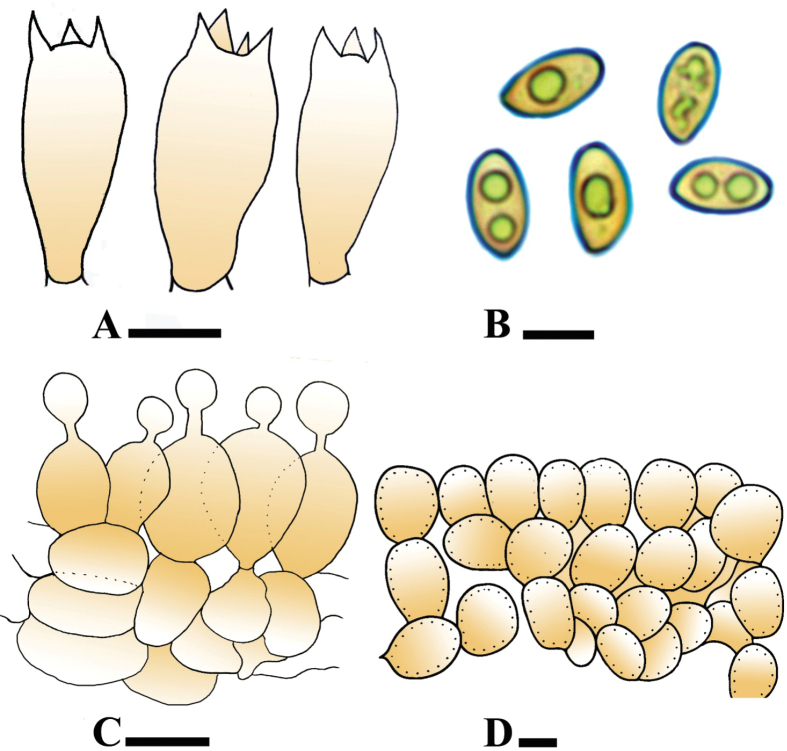 Figure 6
Figure 6| Species names | Locality | Voucher | GenBank No. | |
|---|---|---|---|---|
| ITS | LSU | |||
|
| Italy | WU25461 |
|
|
|
| Italy | WU14367 |
|
|
|
| Hungary | NL-0830 |
|
|
|
| China | HMJAU45055 |
| - |
|
| Hungary | NL-1904 |
|
|
|
| China | HMJAU64952 |
|
|
|
| China | HMJAU64953 |
|
|
|
| Spain | WU27359 |
|
|
|
| Sweden | NL-0908 |
|
|
|
| Sweden | NL-3668 |
|
|
|
| China | HMJAU45033 |
|
|
|
| Pakistan |
|
| |
|
| Slovakia | NL-3499 |
|
|
|
| China | HMAS 300534 |
|
|
|
| China | ZRL20240260 |
|
|
|
| China | HMJAU64954 |
|
|
|
| China | HMJAU64955 |
|
|
|
| China | HMJAU64968 |
| - |
|
| Finland | WU21897 |
|
|
|
| Sweden | NL-2105 |
|
|
|
| Pakistan | KTK05 |
| - |
|
| Pakistan | KTK06 |
| |
|
| Pakistan |
|
| |
|
| Pakistan |
|
| |
|
| Pakistan |
|
| |
|
| Hungary | NL-2380 |
|
|
|
| Hungary | NL-2180 |
|
|
|
| Germany | 40421 |
| - |
|
| China | HMJAU45075 |
|
|
|
| China | HMJAU64939 |
|
|
|
| China | HMJAU64940 |
|
|
|
| China | HMJAU64942 |
|
|
|
| China | HMJAU64943 |
|
|
|
| Italy | WU27557 |
|
|
|
| India | WU20916 |
|
|
|
| China | WANG140019 |
| - |
|
| Austria | WU7395 |
|
|
|
| China | HMJAU64947 |
|
|
|
| China | HMJAU64948 |
|
|
|
| China | HMJAU45062 |
|
|
|
| China | HMJAU64957 |
|
|
|
| China | HMJAU64946 |
|
|
|
| Austria | WU18009 |
|
|
|
| Spain | AH21067 |
| - |
|
| China | FJAU65123 |
|
|
|
| China | HMJAU64964 |
|
|
|
| China | HMJAU64937 |
|
|
|
| China | HMJAU64938 |
|
|
|
| Pakistan |
|
| |
|
| Pakistan |
|
| |
|
| Pakistan |
| - | |
|
| Swedan | NL-2313 |
|
|
|
| China | HMJAU64949 |
|
|
|
| China | HMJAU64950 |
|
|
| China | HMJAU44988 |
|
| |
| China | HMJAU44961 |
| - | |
| China | HMJAU64962 |
|
| |
| China | HMJAU64963 |
|
| |
| China | HMJAU64967 |
| - | |
|
| Pakistan |
|
| |
|
| Pakistan |
|
| |
|
| Turkestan | H7034981 |
|
|
|
| New Zealand | WU17385 |
|
|
|
| Germany | WU28695 |
|
|
|
| Hungary | NL-2187 |
|
|
|
| China | HMJAU45048 |
|
|
|
| Hungary | WU30001 |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Slime Mold and Myxomycetes Research
Introduction
Genus Conocybe Fayod (Bolbitiaceae), is widely distributed in the world and mostly found in temperate regions of Asia, on herbivorous dung, fertile soil, grassland, and lawns etc. (Hausknecht 2009; Song and Bau 2023). It is a toxic fungus that contains toxic substances, such as phallotoxins, psilocybin, and amatoxins, which cause health problems like liver damage, gastroenteritis, and neuropsychological problems (Wu et al. 2019; Song and Bau 2023; Song and Bau 2025). However, these toxic substances also have shown significant pharmacological activities, such as treatment-resistant depression and post-traumatic stress disorder (Griffiths et al. 2016; Song and Bau 2023; Song and Bau 2025). It is characterized by a conical pileus, brown-rusty lamellae, and slender stipe often covered with powdery or hairy, lecythiform cheilocystidia (Song and Bau 2023; Song and Bau 2025). The genus Conocybe consist of 12 sections, viz., Conocybe sect. Candidae Watling, sect. Conocybe Fayod, sect. Conocybella (Singer) Watling, sect. Giganteae Singer, sect. Heterocystidiae E.F. Malysheva, sect. Inopinatae Hauskn. & Contu, sect. Mixtae Singer, sect. Nodulososporae Watling, sect. Obscurae Hauskn. & Krisai, sect. Ochromarasmius (Singer) Hauskn. & Krisai, sect. Pilosellae Singer, and sect. Singerella Watling (Hausknecht and Contu 2007; Hausknecht and Krisai-Greilhuber 2007; Malysheva 2017; Song and Bau 2023; Asif et al. 2025). Our two newly identified taxa belong to sect. Pilosellae, distinguished by an incomplete white stipe, numerous hairs, and non-lecythiform caulocystidia, with C. pilosella as the type species. From Pakistan, only 14 species have been previously reported (Akram et al. 2025; Asif et al. 2025). The aim of this work is to generalize the current knowledge about species diversity of this genus and to provide the detailed descriptions of the species collected from the Pakistan.
Materials and methods
Sampling and study area (Fig. 1)
Specimens were collected during Jun-November 2020–2023, in Bet Faqirwali and Safari Park, bed of the Indus River, District Kot Addu, (30°14'44"N, 70°51'04"E, 134 m a.s.l.), Punjab Province, Pakistan. The climate of Kot Addu is very hot during summer and mild in winter with an approximate highest and lowest temperature of 51 °C and -1 °C respectively; average annual rainfall is 127 mm (Haqnawaz et al. 2023a, 2024a, b; Li et al. 2025). Vegetation of the sampling site is dominated by subtropical plant species such as Albizia chinensis (Osbeck) Merrill Calotropis procera (Ait.) Ait., Hort., Dalbergia sissoo Roxb., Mangifera indica L. Saccharum bengalense Retz., S. officinarum L., Tamarix aphylla (L.) Warb., and Vachellia nilotica (L.) P.J.H.Hurter & Mabb, (Stewart 1972; Haqnawaz et al. 2023a, b). Samples were photographed, tagged, and then dried in front of the electric fan heater at about 35 °C temperature and were deposited in the Lahore Herbarium (LAH), Institute of Botany, University of the Punjab, Lahore, Pakistan.
Maps of sampling sites where the novel taxa were found, shows with the red color.
Morpho-anatomical characterization
Macroscopic characteristics such as size, shape, and color of pileus and stipe were noted, based on fresh samples. Description terminology by Vellinga (2001) was followed, while for color determination, the Munsell color chart was followed (Munsell 1994). Congo red was used for dyeing. The microscopic features like size, shape, and color of basidiospores, basidia, cheilocystidia, pileipellis, and caulocystidia were observed using a compound microscope (OLYMPUS BX43, Tokyo, Japan), and measurements were taken using calibrated Piximètre software connected to a compound microscope through a HDCE-X5 microscopic camera. In the case of basidiospores, at least 50 spores were measured. The formula [n/m/p] indicates ‘n’ the number of basidiospores measured from ‘m’ basidiomata of ‘p’ collection. Basidiospores measurements are presented as (a–) b–c (–d), where b–c indicates 90% of the calculated values, while values in parenthesis are extreme lowest or extreme highest values; Q indicates the individual spore length/width ratio while Qav. is the average length/width ratio of all basidiospores (Ge et al. 2010).
Molecular phylogenetic analyses
DNA was extracted from the dried specimens using the 2% CTAB method (Bruns 1995). Primers used during amplifications were ITS1F 5’-CCT GGT CAT TTA GAG GAA GTA A-3’ as forward and ITS4 5’-TCC TCC GCT CTA TTG ATA TGC-3’ as reverse for nrITS region (>775 bp) while LROR 5’-ACC CGC TGA ACT TAA GC-3’ as forward and LR5 5’-TCC TGA GGG AAA CTT CG-3’ as reverse for nrLSU region (>910 bp) (Gardes and Bruns 1993, White et al. 1990). The Polymerase Chain Reaction (PCR) was performed in a 50 µl reaction volume: containing 5 µl 10× Econo Taq Buffer (Lucigen, Middleton, Wisconsin, USA), 1 µl dNTPs, 2.5 µl of each primer (10 µM/µl), 0.25 µl of Econo Taq® DNA Polymerase (Lucigen), 28.75 µl H_2_O and 10 µl DNA template. All PCR amplifications were carried out with the cycling program for 35 cycles as follows, denaturation at 94 °C for 30 sec, annealing at 54 °C for 1 min, extension at 71 °C for 2 min, with an initial denaturation at 94 °C for 2 min and a final extension of 71 °C for 5 min. PCR products were then sequenced and analyzed (Usman and Khalid 2020; Haqnawaz et al. 2023b).
Forward and reverse primer reads of both ITS and nrLSU regions were combined and consensus sequences were obtained using Bioedit. ver. 7.2.5 (Hall 1999). The resulting sequences were subjected to a BLAST search at NCBI (https://www.ncbi.nlm.nih.gov/guide/). The sequences of closely related species from GenBank and sequences used from a recent publication of Conocybe, were included in our final phylogenetic tree (Asif et al. 2025; Song and Bau 2025). One sequence of Bolbitius reticulatus (Pers.) Ricken was used as an outgroup for the ITS and LSU datasets. For the phylogenetic analyses, a Clustal W MUSCLE alignment was implemented in BioEdit v. 7.2.5 with manual adjustments. A combined (ITS–LSU) maximum likelihood phylogenetic tree was constructed using RAxML-HPC2 v. 8.2.12 on XSEDE (8.2.10) implemented on the CIPRES Science Gateway (Miller et al. 2010). The GTR+GAMMA nucleotide substitution model was used, with 1,000 bootstrap iterations performed using rapid bootstrapping. The resulting bootstrap values were then mapped on to the best-scoring ML tree and show in Fig. 2. The model of evolution was estimated by MrModeltest 2.2 (Nylander 2004). Markov chain Monte Carlo (MCMC) sampling in MrBayes v. 3.2.2 (Ronquist et al. 2012) was used to determine posterior probabilities (PP). Six simultaneous Markov chains were run for 1,000,000 generations, and trees were sampled every 1,000^th^ generation. Bootstrap values ≥ 50% and Bayesian PP ≥ 0.90 are indicated on the branches, which were visualized in FigTree v. 1.4.2 (Rambaut 2014). The newly generated sequences were submitted to GenBank, and short descriptions of the species were deposited in MycoBank (Robert et al. 2013). Boot strap values ≥ 50% are mentioned on the branches that were visualized in FigTree v. 1.4.2 (Rambaut 2014). The newly generated sequences were deposited in GenBank, and short descriptions of the species were deposited in MycoBank (Robert et al. 2013). All the sequences used in the final phylogenetic tree are presented in Table 1, together with voucher numbers, GenBank accession numbers, and country of origin. The newly generated sequences were deposited in GenBank and are in bold in the phylogenetic trees and Table 1.
Results
ITS & LSU phylogenetic analyses (Fig. 2, Table 1)
The combined phylogenetic tree of the nrITS & nrLSU region based on consists of 65 sequences, and Bolbitius reticulatus (WU30001) is an outgroup. Both new taxa are shown in bold, and formed separate clades from their closely related species. Conocybe kotadduensis formed a separate branch with 78% bootstrap support from its sister species, Conocybe nitrophila (Hauskn.) Yen W. Wang & S.S. Tzean (WU20916 & WANG140019); and the other closest species is Conocybe pilosella (Pers.) Kühner (HMJAU45062 & HMJAU64957). The second new species Conocybe safariensis also formed a separate branch from its sister species C. ceracea T. Bau & H.B. Song (HMJAU64952 & HMJAU64953) with high bootstrap value 94%, and C. himalayana Ke Wang, T.Z. Wei & P. Hong (HMAS300534 & ZRL20240260) with 89% bootstrap value.
Phylogenetic tree of the genus Conocybe as generated by maximum likelihood (ML) and Bayesian analyses, based on combined ITS and LSU sequences. Bootstrap values ≥ 50%, based on 1,000 replicates and Bayesian posterior probabilities (PP) ≥ 0.90, are shown at the branches. Novel sequences, generated during this study, are shown in bold.
Taxonomy
Conocybe
kotadduensis
Taxon classificationFungiAgaricalesBolbitiaceae
Haqnawaz, Niazi & Khalid sp. nov.
82AB4563-07F0-578D-8DB2-BF0990F2C48D
855941
Etymology.
Species name “kotadduensis” (Latin) refers to the type locality Kot Addu, Punjab, Pakistan.
Holotype.
Pakistan • Punjab, Kot Addu District, bed of Indus River, (30°23'27"N, 70°48'28"E, 127 m a.s.l.), on plant debris, June 14, 2023, Muhammad. Haqnawaz, KA-06 (LAH38248). GenBank: PQ409233 (nrITS), PQ409236 (nrLSU).
Diagnosis.
Conocybe kotadduensis is different from the closest species, C. nitrophila, by its relatively light gray to pale yellow, parabolic absence of thick glutinous veil and viscid in regular to slightly crenate margins of pileus, adnexed, broad, thick lamellae with 2–3 tires of lamellulae, scabrous surface of stipe, amygdaliform to ovoid basidiospores, mostly broad bi-sterigmate of basidia, clavate, utriform with median constriction, fusiform, cylindrical shape of lecythiform cheilocystidia, sphero-pedunculate to obovoid, globose to sub-globose pileipellis elements.
A, B. Basidiomata of Conocybe kotadduensis. (A. Holotype LAH38248); C. Sampling site. Scale bars: 10 mm (A, B).
Description.
Pileus 10–13 mm diam., hemispherical to parabolic at young stage, conical, parabolic when old, straight shape of margins, regular to slightly crenate cap margins, velutinous surface, light gray (7.5 YR 8/1) to pale yellow (2.5 Y 8/4), with dull yellow orange (10YR 7/4) center. Lamellae adnexed, broad, thick, subdistant, fimbriate, unequal, with exceeding 1–3 tires, dull yellow orange (10 YR 7/4) to orange (7.5 YR 7/6). Stipe 25–49 mm in length, cylindrical, flexuous, central, with slightly scabrous surface, hard and dry, slightly broad toward base, orange (5 Y.R 6/6) at upper side, dull yellow orange (10 YR 7/4) at base, grayish white (N 8/0).
A–D. Microscopic structures of Conocybe kotadduensis. (A. Holotype LAH38248). A. Basidia; B. Basidiospores; C. Cheilocystidia; D. Pileipellis elements. Scale bars: 10 µm.
Basidiospores [100/2/3], (13.2–)13.5–15.5 (–16) × (8–)8.5–9.5(–10) µm, avl × avw = 14.7 × 9.0 µm, Q = 1.47–1.69, Q_av_ = 1.62, amygdaliform to ovoid, yellowish to slightly light brown in 5% KOH, thin walled, guttulate, prominent germ pore. Basidia [30/2/3] (24–)24–28(–29) × (10.5–)11–11(–11.5) µm, clavate, with 2 sterigmata with broad width, hyaline in 5% KOH, thin-walled. Cheilocystidia [30/3/3] (14–) 14.5–24(–24.5) × (5.5–)6.–10.5(–11) µm, lecythiform with clavate, utriform with median constriction, fusiform, cylindrical shap of cells, rarely lageniform, hyaline, thin walled. Pileipellis [30/2/3] (25–89 × 1637 µm, sphero-pedunculate to obovoid, globose to sub-globose, hyaline and thin walled. Stipitipellis 8–23 µm, avw = 9.8 µm, cutis, hyaline in 5% KOH, thin walled, septate, unbranched, regular. Caulocystidia [15/3/3] 9–14 × 5–7 µm, rarely present, clavate to cylindrical, hyaline in 5% KOH, thin walled. Clamp connection absent.
Ecology and habitat.
Gregarious, terrestrial, under plants of Saccharum bengalense and Cyperus rotundus.
Additional specimens examined.
Pakistan • Punjab, Kot Addu, bed of Indus River, 30°25'19"N, 70°52'44"E, 132 m a.s.l., June 26, 2020, FW-98, Muhammad Haqnawaz, (Paratype LAH38249), GenBank: PQ409234 [nrITS], PQ409237 [nrLSU]. • 30°14'44"N, 70°51'04"E, 134 m a.s.l. under Cyperus rotundus, August 15, 2021, TA-51, Muhammad Haqnawaz (Paratype LAH38250), GenBank: PQ409235 (nrITS), PQ409238 (nrLSU).
Conocybe
safariensis
Taxon classificationFungiAgaricalesBolbitiaceae
Niazi, Haqnawaz & Khalid sp. nov.
8478AAFB-264F-5B88-90FE-896C85021484
854578
Etymology.
Species name safariensis (Latin) refers to the locality of the taxon. i.e., Safari Park, Kot Addu, Punjab, Pakistan.
Holotype.
Pakistan • Punjab, Kot Addu District, bed of Indus River, (30°31'33.63"N, 70°56'29.71"E, 135 m a.s.l.), on plant debris under Tamarix aphylla, July 05, 2021, Muhammad. Haqnawaz, HQ-69 (LAH38252). GenBank: PP973139 (nrITS), PP973145 (nrLSU).
Diagnosis.
Conocybe safariensis is different from the closest species, C. ceracea, by its hemispherical, lentiform, obtusely conical, then convex to plane and straight margin of pileus, orange to light gray stipe, obovoid to lacrymoid basidiospores, obovoid, ellipsoid to globose cheilocystidia, oblong, globose, clavate and fusiform elements of pileipellis, ellipsoid to broadly ellipsoid, clavate to broadly clavate caulocystidia, and absence of pleurocystidia.
A. Basidiomata of Conocybe safariensis. (A. Holotype LAH38252); B. Sampling site, Scale bar: 10 mm (A).
Description.
Pileus 15–24 mm long, hemispherical to campanulate at young stage, sub hemispherical, lentiform, obtusely conical, then convex to plane when old, slightly regular, straight shape and sulcate cap margins, smooth, light gray to dull orange (7.5 YR 8/2, 7/4), texture soft, thin, light gray (7.5 YR 8/2), Lamellae adnate, broad, average, sub distant, even, unequal, exceeding, and 3–7 tires, orange (7.5 YR 7/6). Stipe 25–49 mm in length, flexuous, white scabrous surface, costate surface, bulbous base, orange (7.5 YR 4/3) at the base, light brownish gray (7.5 YR 7/2) to light gray (5 YR 8/1) toward base.
A–D. Microscopic structures of Conocybe safariensis. (A. Holotype LAH38252). A. Basidia; B. Basidiospores; C. Cheilocystidia; D. Pileipellis. Scale bars: 10 µm (A–D).
Basidiospores [100/2/3], (8–) 8.6–10 (–10.5) × (4.5–) 4.8–5.5(–6) µm, avl × avw = 9.5 × 5.2 µm, Q = 1.63–2.2, Qav = 1.8, obovoid to lacrymoid, yellowish green to light reddish brown in water, guttulate, thick walled, prominent germ pore present. Basidia [30/2/3] (15–)16–23(–24) × (7–)8–12(–13) µm, clavate to broadly clavate, slightly cylindrical, with 1–4 sterigmata, hyaline in 5% KOH, amyloid, thin–walled, guttulate. Cheilocystidia [40/2/3] 16–25 × 8–12 µm, lecythiform with ellipsoid to broadly ellipsoid, globose to subglobose, clavate to broadly clavate cells, hyaline, thick walled. Pleurocystidia absent. Pileipellis [30/2/3] 20–50 × 18–36 µm, irregular epithelium, oblong, globose to subglobose, clavate to broadly clavate, fusiform elements, thin-walled. Stipitipellis 8–17 µm, avw = 12.5 µm, regular, trichoderm, hyaline, thin walled, septate, unbranched, regular, clamp connection present. Caulocystidia [30/2/3] 11–33 × 7–13 µm, rearly present, cylindrical ellipsoid to broadly ellipsoid, globose to subglobose, clavate to broadly clavate cells hyaline in 5% KOH.
Ecology and habitat.
Gregarious, terrestrial, under Tamarix aphylla, on loamy soil rich in organic matter.
Additional specimens examined.
Pakistan • Punjab, Kot Addu, Noor Shah Thal, 30°31'33.63"N, 70°56'29.71"E, 135 m a.s.l., September 29, 2022, Muhammad Haqnawaz, (Paratype LAH38251), GenBank: PP973138 [nrITS], PP973144 [nrLSU]. • 30°25'21"N, 70°52'46"E 132 m. a.s.l., September 10, 2023, MQ-151, Muhammad Haqnawaz (Paratype LAH38253), GenBank: PP973140 (nrITS).
Discussion
Two new species, Conocybe kotadduensis and C. safariensis, were described based on phylogenetic analyses and morpho-anatomical descriptions. Phylogenetic analysis of nrITS and nrLSU sequence data showed that Conocybe kotadduensis and C. safariensis belong to section Pilosellae Singer (Song and Bau 2023). Conocybe kotadduensis, formed a separate branch from C. nitrophila, C. pilosella, and C. pseudocrispa with 18, 25, and 21 base pairs differences, respectively. While Conocybe safariensis is different from its sister’s species, C. ceracea, C. himalayana and C. pallidospora with 14, 19, and 38 base pairs differences, respectively.
Morpho-anatomically, Conocybe kotadduensis is distinguished from its sister species, C. nitrophila, by having pale yellow, parabolic pileus with velvety surface (vs. orange white, hygrophanous, hemispherical to a little convex pileus with smooth surface), scabrous surface of stipe (vs. smooth), amygdaliform to ovoid, yellowish to slightly light brown basidiospores (vs. broadly ellipsoid to oval, brownish orange to reddish brown), broad, fusiform, cylindrical cells of lecythiform cheilocystidia (vs. only clavate shape of cells), sphero-pedunculate to obovoid, globose to sub-globose pileipellis elements (vs. only broadly cylindrical to clavate). C. nitrophila has thick glutinous veil and viscid in fresh sample while absence in new taxon (Wang and Tzean 2015). The second closer species, Conocybe pilosella differs in having subpilose pileus covered with hair, tetra-sterigmate basidia, absent germpore, lecythiform pileocystidia (Song and Bau 2023). Another species, C. pseudocrispa, is different due to its oyster white surface with green beige to ivory center of pileus, narrowly adnate, beige-brown to ochre-brown lamellae, longitudinally striated, fine hairs, thick base of stipe, honey yellow to ochre-brown, ellipsoid to oblong basidiospores, subglobose, lageniform, lanceolate stipitipellis elements (Song and Bau 2023).
Similarly, morpho-anatomically, Conocybe ceracea differs from our second new taxon, C. safariensis, possessing subglobose, paraboloid to subcylindrical pileus, incurved margins, and presence of precipitate wax crystals on the surface of the pileus, honey yellow to melon yellow, elliptical to oblong and ovoid basidiospores, presence of pleurocystidia and subglobose, lageniform, cylindrical, or lanceolate caulocystidia (Song and Bau 2023). Similarly, C. himalayana a Chinese taxon, differs due to its decurved margins, subglobose, orange brown pileus with faintly pubescent surface, sometimes stipe forming a bulb, ovoid to ellipsoid basidiospores, lageniform cheilocystidia (Wang et al. 2024). Ecologically, C. safariensis is, terrestrial, grows under Tamarix aphylla, whereas its sister’s species C. ceracea grows under potted Orchidaceae plants, and C. himalayana grows on grassland (Song and Bau 2023; Wang et al. 2024). With these additions, the taxa of Conocybe known from Pakistan turn out to be 16.
Supplementary Material
XML Treatment for Conocybe kotadduensis
XML Treatment for Conocybe safariensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akram W, Saba M, Asif M, Maula F, Batool M, Kamal A, Noor A (2025) Combined morphological and molecular approaches reveal Conocybe pakistanensis (Bolbitiaceae), a novel species from Punjab, Pakistan. Phytotaxa 684(1): 93–104. 10.11646/phytotaxa.684.1.5 · doi ↗
- 2Asif M, Saba M, Maula F, Akram W, Kamal A, Raza M (2025) Multilocus phylogeny contributes to fungal taxonomy with three novel species of Conocybe (Bolbitiaceae) from southern Punjab, Pakistan. Mycologia 117(3): 463–479 [1–7]. 10.1080/00275514.2025.246327940068134 · doi ↗ · pubmed ↗
- 3Bruns TD (1995) Thoughts on the processes that maintain local species diversity of ectomycorrhizal fungi. Plant and Soil 170: 63–73. 10.1007/BF 02183055 · doi ↗
- 4Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for Basidiomycetes-Application to the identification of mycorrhizae and rusts. Molecular Ecology 2: 113–118. 10.1111/j.1365-294X.1993.tb 00005.x 8180733 · doi ↗ · pubmed ↗
- 5Ge ZW, Yang ZL, Vellinga EC (2010) The genus Macrolepiota (Agaricaceae, Basidiomycota) in China. Fungal Diversity 45(1): 81–98. 10.1007/s 13225-010-0062-0 · doi ↗
- 6Griffiths RR, Johnson MW, Carducci MA, Umbricht A, Richards WA, Richards BD, Cosimano MP, Klinedinst MA (2016) Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: A randomized double-blind trial. Journal of Psychopharmacology 30(12): 1181–1197. 10.1177/0269881116675513 PMC 536755727909165 · doi ↗ · pubmed ↗
- 7Haqnawaz M, Niazi AR, Khalid AN (2023 a) Two new species of Xanthagaricus from Punjab, Pakistan. Phytotaxa 583(2): 163–173. 10.11646/phytotaxa.583.2.4 · doi ↗
- 8Haqnawaz M, Niazi AR, Khalid AN (2023 b) A study on the genus Candolleomyces (Agaricales: Psathyrellaceae) from Punjab, Pakistan. BMC Microbiology 23(1): 1–8. 10.1186/s 12866-023-02938-2PMC 1033461837434121 · doi ↗ · pubmed ↗