The conserved microtubule-associated protein Mini Spindles promotes dendrite branching and limits terminal branch elongation in Drosophila class III and IV dendritic arborization neurons
Claire Kittock, Noor Anvery, Mala Misra

TL;DR
This study shows that the protein Msps helps regulate dendrite branching and growth in fruit fly neurons.
Contribution
The novel finding is that Msps promotes dendrite branching and limits elongation during larval growth in Drosophila neurons.
Findings
Knocking down Msps reduces dendrite branching but increases terminal dendrite length.
Msps RNAi disrupts proportional dendrite growth during larval development.
Msps is essential for dynamic dendrite arbor expansion during rapid organismal growth.
Abstract
Microtubule dynamics influence neuron morphogenesis. We investigated the role of the conserved microtubule-associated protein Mini Spindles (Msps) in the morphogenesis of branched dendrite arbors using Drosophila melanogaster larval class III and class IV dendritic arborization neurons as two models of branch organization. In both classes, knocking down msps expression reduced dendrite branching but increased terminal dendrite length. In msps RNAi class IV da neurons, dendrite growth failed to scale in proportion to increasing larval size between the second and third instar. These results suggest that Msps is required for the dynamic expansion of dendrite arbors during periods of rapid organismal growth.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
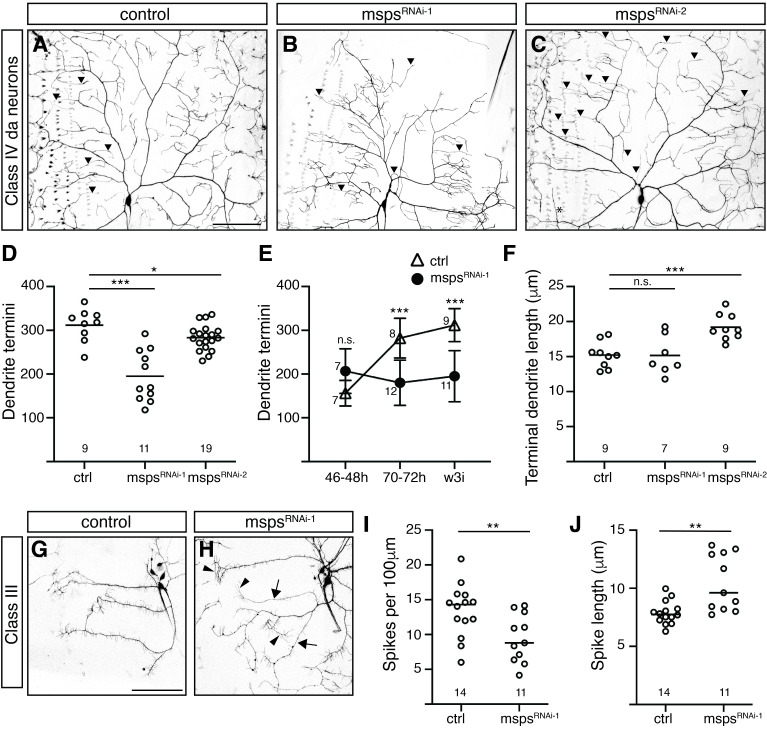 Figure 1
Figure 1|
|
|
|
|
|
|
|
ppk-GAL4, UAS-CD4:tdGFP VK0003 |
Gift from Elizabeth Gavis, Princeton University |
BDSC_35836 BDSC_32079 |
(Bhogal et al., 2016) |
|
|
1003.3-GAL4; UAS-mCD8-GFP |
Gift from Elizabeth Gavis, Princeton University |
FBti0128079 |
(Hughes & Thomas, 2007; Lee & Luo, 1999) |
|
|
UAS-CD4:tdGFP VK0003 |
Bloomington Stock Center |
BDSC_35836 |
(Han et al., 2011) |
|
|
UAS-mspsRNAi HMS01906 |
Bloomington Drosophila Stock Center (Transgenic RNAi Project) |
BDSC_38990 |
(Misra et al., 2016; Das et al., 2017) |
|
|
UAS-mspsRNAi JF01613 |
Bloomington Drosophila Stock Center (Transgenic RNAi Project) |
BDSC_31138 |
(Gao et al., 2024) |
|
|
yw |
Laboratory stock |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrotubule and mitosis dynamics · Neurobiology and Insect Physiology Research · Developmental Biology and Gene Regulation
Description
Neurons modify their structure and connectivity throughout their development in response to shifting internal and environmental cues. These changes require cytoskeletal rearrangements, and many previous studies have identified cytoskeleton-associated proteins as mediators of structural adaptation and refinement (Ouzounidis et al., 2023). *Drosophila melanogaster * larval dendritic arborization neurons (da neurons) are a valuable model for investigating the role of cytoskeletal regulators in neuron morphogenesis due to their complex branching patterns and their accessibility for imaging. In embryos and larvae, four classes of abdominal da neurons extend dendrites across cutaneous receptive fields to detect distinct environmental stimuli (reviewed in Singhania & Grueber, 2014). Class IV da neurons (C4da) and class III da neurons (C3da) have the most complex arbors but differ in branching structure. C4da neurons adopt an ordered branching structure, where primary dendrites bifurcate to form higher-order branches. In contrast, C3da arbors have long primary branches decorated with short, unbranched filopodial extensions (“spikes”) that emerge roughly perpendicular to the parent branch ( Fig. 1A and 1G; Grueber et al., 2002). In both neuron subtypes, terminal dendritic processes exhibit dynamic extension and retraction during larval stages (Grueber et al., 2003; Tsubouchi et al., 2012).
The microtubule plus end-associated polymerase Mini Spindles *, * a homolog of Xenopus XMAP25, was previously identified as a positive regulator of dendrite branch morphogenesis in C4da neurons (Das et al., 2017; Misra et al., 2016) and dendrite pruning during metamorphosis (Tang et al., 2020). We sought to further characterize the role of msps in da neurons during larval growth by examining the effects of reduced expression. To knock down msps , we used the C4da-specific driver pickpocket-Gal4 (ppk-Gal4) or the C2/C3da-specific driver 1003.3-Gal4 to drive expression of two msps -interfering shRNAs (“ * msps ^RNAi-1^ * ” and “ * msps ^RNAi-2^ * ”) via an upstream activating sequence.
In wandering third instar larvae, loss of msps activity caused a decrease in the number of terminal branches within the dendrite arbor of the ddaC neuron, a C4da neuron positioned within the dorsal da neuron cluster ( Fig. 1B- D). This result was consistent with previously reported findings (Das et al., 2017; Misra et al., 2016; Tang et al., 2020). The severity of the effects varied between shRNAs, with * msps ^RNAi-1^ * eliciting a stronger effect (37% reduction) and * msps ^RNAi-2^ * eliciting a mild but statistically significant effect (9% reduction). When we compared the number of dendritic termini at 48h and 72h after egg laying (AEL) to our results from wandering third instar (112-120h AEL), we found that the number of termini increased at each time point in control neurons but remained constant for * msps ^RNAi-1^ * neurons ( Fig. 1E ). These findings support the conclusion that Msps is essential for the addition and/or maintenance of new branches during the period of “scaling growth”, 48h-120h AEL, when neuron arbors expand so their receptive fields can keep up with increasing larval surface area (Parrish et al., 2009). Previous studies have identified Msps as a regulator of acentrosomal microtubule nucleation (Deng et al., 2021); others have found that acentrosomal microtubule nucleation is essential for dendrite branch growth and stabilization (Ori-McKenney et al., 2012). Msps may be part of the molecular infrastructure that links these processes.
We also observed that * msps ^RNAi-1^ * and * msps ^RNAi-2^ * C4da neurons occasionally exhibited very long terminal branches. The average terminal dendrite length was significantly higher in * msps ^RNAi-2^ * neurons than control (23% increase; Fig. 1F ). This was not true in * msps ^RNAi-1^ * neurons; however, the effect may have been masked by severely depleted arbors overall. Together, our results suggest that Msps may function to promote the initiation of higher order dendrite branches while simultaneously protecting against excessive growth or erroneous maintenance of existing dendrites.
To determine if Msps plays a similar role in other neuronal subtypes, we examined the impact of knocking down *msps * expression in C3da neurons. As in C4da neurons, expression of the shRNA * msps ^RNAi-1^ * caused a reduction in the total number of branches – in this case, spike-like filopodia – in the dorsal class III neuron ddaA. This resulted in a decrease in spike density (29% reduction; Fig. 1G- H, I). In addition, the average length of spikes increased in * msps ^RNAi-1^ * ddaA neurons, echoing earlier observations in dorsal C4da neurons (33% increase; Fig. 1G- H, J). Further investigations will be required to confirm these results in * msps ^RNAi-2^ * neurons. Of note, previous studies found that the distribution of cytoskeletal components varies in these two da neuron subtypes (Andersen et al., 2005; Tsubouchi et al., 2012). In C4da neurons, microtubules are present throughout the arbor; in C3da neurons, however, the filopodial spikes that protrude from primary dendritic branches are notably actin-rich and microtubule-poor. Our observations suggest that microtubule-associated proteins like Msps may significantly impact the growth and stability of these actin-based structures despite their relatively low microtubule content.
In conclusion, our findings suggest that the microtubule plus-end polymerase Mini Spindles promotes branch initiation and limits terminal dendrite growth in both C4da and C3da neurons during the period of scaling growth in *Drosophila * larvae. It is yet unclear whether Msps restricts terminal dendrite length by preventing excessive growth or by inhibiting maintenance. Tang and colleagues (2020) found that loss of msps function led to excessive maintenance of dendrites and a failure of pruning in C4da neurons during metamorphosis. They attributed this change in stability to the randomization of microtubule orientation in dendrites, a phenomenon that has also been linked to defects in branching (Sears & Broihier, 2016). Future studies may investigate whether Msps protects against excessive maintenance and stabilization of terminal dendrites to enable dendrite flexibility for growth, retraction, and branch initiation during the dynamic process of scaling neuron growth in *Drosophila * larvae.
Methods
** Drosophila strains and genetics **
*D. melanogaster * stocks were reared on Nutri-Fly Instant or Nutri-Fly Bloomington Formulation media. Crosses were performed at 25°C for analysis of C4da neurons or at 29°C for analysis of C3da neurons. The following stocks were used: ppk-GAL4 , UAS-CD4-GFP and 1003.3-GAL4; UAS-mCD8-GFP (shared by Elizabeth Gavis, Princeton University); UAS-msps ^HMS01906^ (“UAS-msps ^RNAi-1^ ” in the text) and UAS-msps ^JF01613^ (“UAS-msps ^RNAi-2^ ” in the text) (Bloomington Drosophila Stock Center). In all experiments, yw flies served as the control.
Microscopy
Images were obtained using 20X/0.7 NA air and 40X/1.23 NA oil objectives on a Zeiss LSM710. For live imaging of GFP-expressing neurons, an imaging chamber was created by adhering two small (22x22mm) coverslips 5-7mm apart on a glass slide. Larvae were mounted between the two coverslips in a 1:2:2 mix of chloroform, halocarbon 95, and halocarbon 200. A long (24x50mm) coverslip was placed on top of the sample and held in place with laboratory tape. Dorsal C3da (ddaA and ddaF) or C4da (ddaC) neurons were imaged from abdominal segments 2 and 3. Neurons were imaged using tiled capture and automatic stitching. Images of C3da neurons comprised four tiles with overall dimensions of 850x850 μm. Images of dorsal C4da neurons include two tiles with overall dimensions of 425x850 μm. For C4da neurons, the imaging field typically included the dorsal, anterior, and posterior edges of the dendritic arbor but not branches extending ventrally from the cell body.
Quantitative image analysis
All quantitative image analysis was carried out using ImageJ (v2.1.0/1.53c). The Cell Counting plugin was used to count and mark all dendrite termini within the imaging field. For C3da neurons, the total number of termini was divided by the length of the primary dendrites to obtain a measure of spike density. Dendrite length was measured using the NeuronJ plugin (Meijering et al., 2004). For Class IV neurons, all terminal dendrites dorsal to the cell body within the image field were measured on neuron ddaC. For Class III, all spikes from neuron ddaA within the image field were measured. A range of 7-19 neurons from 3-6 larvae were analyzed for each experiment.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andersen Ryan Li Yimei Resseguie Mary Brenman Jay E. 2005928 Calcium/Calmodulin-Dependent Protein Kinase II Alters Structural Plasticity and Cytoskeletal Dynamics in Drosophila The Journal of Neuroscience 25390270-64748878888810.1523/jneurosci.2005-05.200516192377 PMC 6725600 · doi ↗ · pubmed ↗
- 2Bhogal Balpreet Plaza-Jennings Amara Gavis Elizabeth R. 201611 Nanos-mediated repression of hid protects larval sensory neurons after a switch in sensitivity to apoptotic signals Development 1477-912910.1242/dev.132415 PMC 492017027256879 · doi ↗ · pubmed ↗
- 3Das Ravi Bhattacharjee Shatabdi Patel Atit A Harris Jenna M Bhattacharya Surajit Letcher Jamin M Clark Sarah G Nanda Sumit Iyer Eswar Prasad R Ascoli Giorgio A Cox Daniel N 20171012 Dendritic Cytoskeletal Architecture Is Modulated by Combinatorial Transcriptional Regulation in Drosophila melanogaster Genetics 20741943-26311401142110.1534/genetics.117.30039329025914 PMC 5714456 · doi ↗ · pubmed ↗
- 4Deng Qiannan Tan Ye Sing Chew Liang Yuh Wang Hongyan 202189 Msps governs acentrosomal microtubule assembly and reactivation of quiescent neural stem cells The EMBO Journal 40190261-418910.15252/embj.2020104549 PMC 848857234368973 · doi ↗ · pubmed ↗
- 5Gao Z Huang E Wang W Xu L Xu W Zheng T Rui M 2024118 Patronin regulates presynaptic microtubule organization and neuromuscular junction development in Drosophila.i Science 27210894410894410.1016/j.isci.2024.10894438318379 PMC 10839449 · doi ↗ · pubmed ↗
- 6Grueber Wesley B. Jan Lily Y. Jan Yuh Nung 2002615 Tiling of the Drosophila epidermis by multidendritic sensory neurons Development 129121477-91292867287810.1242/dev.129.12.286712050135 · doi ↗ · pubmed ↗
- 7Grueber Wesley B. Ye Bing Moore Adrian W. Jan Lily Y. Jan Yuh Nung 200341 Dendrites of Distinct Classes of Drosophila Sensory Neurons Show Different Capacities for Homotypic Repulsion Current Biology 1380960-982261862610.1016/s 0960-9822(03)00207-012699617 · doi ↗ · pubmed ↗
- 8Han Chun Jan Lily Yeh Jan Yuh-Nung 2011523 Enhancer-driven membrane markers for analysis of nonautonomous mechanisms reveal neuron–glia interactions in Drosophila Proceedings of the National Academy of Sciences 108230027-84249673967810.1073/pnas.1106386108 PMC 311128821606367 · doi ↗ · pubmed ↗