The genome sequence of the Osiris Blue, Cupido osiris (Meigen, 1830) (Lepidoptera: Lycaenidae)
Andreas Sanchez, Yannick Chittaro, Kay Lucek, Charlotte J. Wright, Joana I. Meier, Mark L. Blaxter, Sivasankaran Kuppusamy, Sophie Tandonnet, María Morán Saiz

TL;DR
The genome of the Osiris Blue butterfly is sequenced, providing insights into its genetic makeup and contributing to a larger project on European Lepidoptera.
Contribution
The study provides a high-quality genome assembly for the Osiris Blue butterfly, including haplotype-specific scaffolding and mitochondrial genome data.
Findings
The genome assembly includes two haplotypes totaling 481.43 and 479.83 megabases.
Haplotype 1 is scaffolded into 24 chromosomal pseudomolecules, including the Z sex chromosome.
The mitochondrial genome is assembled with a length of 15.36 kilobases.
Abstract
We present a genome assembly from a male Cupido osiris (Osiris Blue; Arthropoda; Insecta; Lepidoptera; Lycaenidae). The assembly contains two haplotypes with total lengths of 481.43 megabases and 479.83 megabases. Most of haplotype 1 (98.72%) is scaffolded into 24 chromosomal pseudomolecules, including the Z sex chromosome. Haplotype 2 was assembled to scaffold level. The mitochondrial genome has also been assembled, with a length of 15.36 kilobases. This work is part of Project Psyche, a collaborative programme generating genomes for European butterflies and moths.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1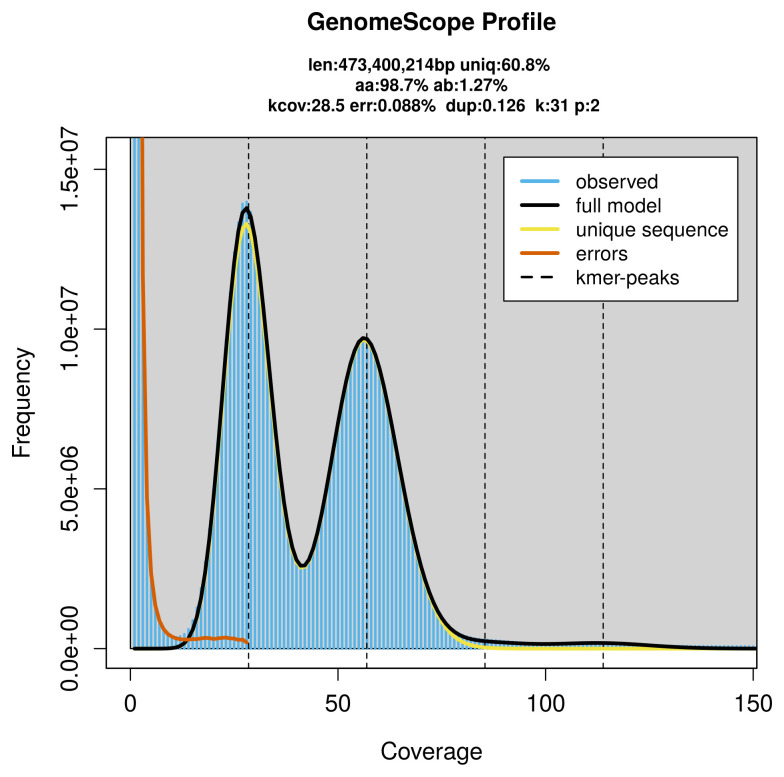 Figure 2
Figure 2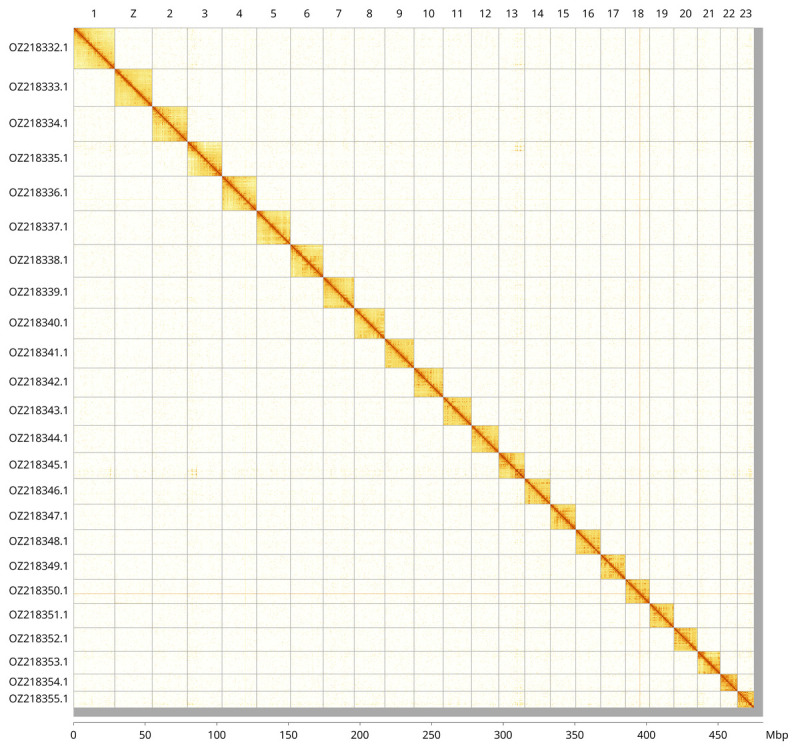 Figure 3
Figure 3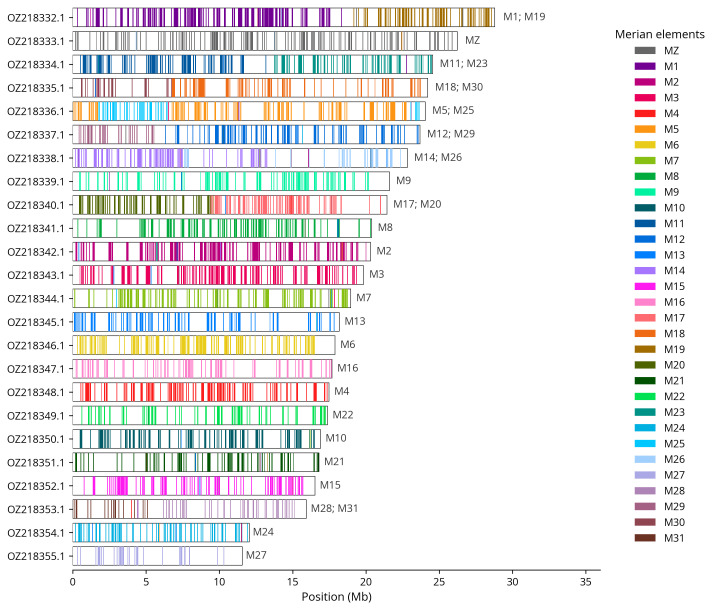 Figure 4
Figure 4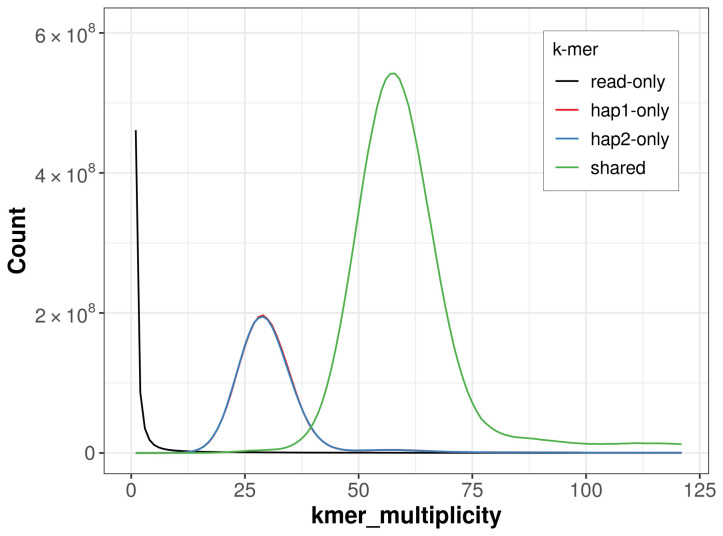 Figure 5
Figure 5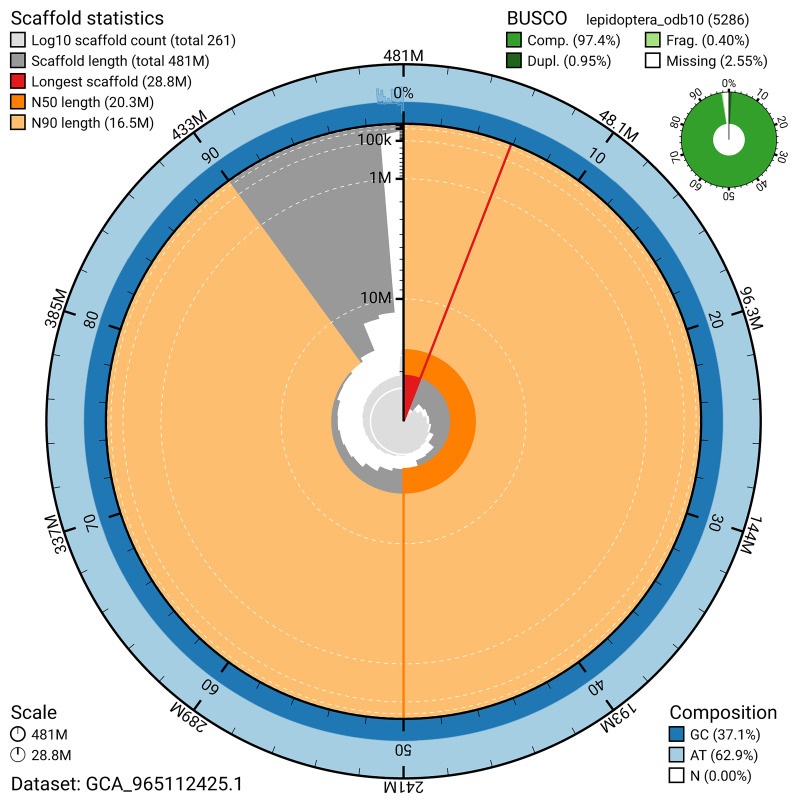 Figure 6
Figure 6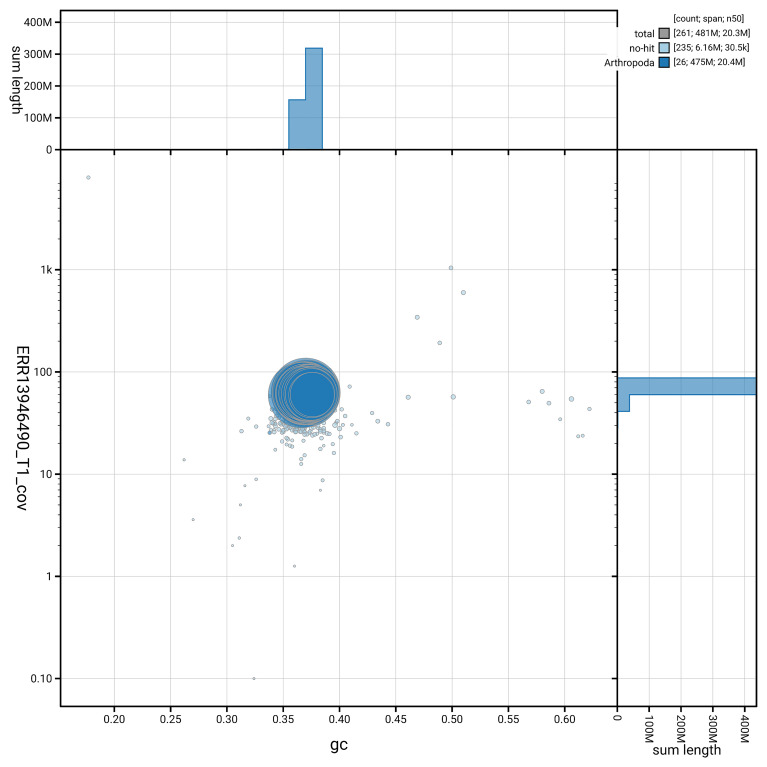 Figure 7
Figure 7| Platform | PacBio HiFi | Hi-C |
|---|---|---|
|
| ilCupOsir1 | ilCupOsir1 |
|
| SAN28000112 | SAN28000112 |
|
| SAMEA115109942 | SAMEA115109942 |
|
| SAMEA115109978 | SAMEA115109979; SAMEA115109977 |
|
| thorax | abdomen; head |
|
| Revio | Illumina NovaSeq X |
|
| ERR13946490 | ERR13947518; ERR13947517 |
|
| 2.82 million | 1 259.01 million |
|
| 27.78 Gb | 190.11 Gb |
| Assembly name | ilCupOsir1.hap1.1 | ilCupOsir1.hap2.1 |
|---|---|---|
|
| GCA_965112425.1 | GCA_965112415.1 |
|
| chromosome | scaffold |
|
| 481.43 | 479.83 |
|
| 24 | scaffold-level |
|
| 358 | 250 |
|
| 7.22 Mb | 7.02 Mb |
|
| 260 | 167 |
|
| 20.3 Mb | 20.27 Mb |
|
| 28.79 | - |
|
| Z | - |
|
| Mitochondrion: 15.36 kb | - |
| INSDC accession | Molecule | Length (Mb) | GC% | Assigned Merian elements |
|---|---|---|---|---|
| 1 | 28.79 | 37 | M1;M19 | |
| 2 | 24.55 | 37 | M11;M23 | |
| 3 | 24.22 | 37 | M18;M30 | |
| 4 | 24.05 | 37 | M25;M5 | |
| 5 | 23.69 | 37 | M12;M29 | |
| 6 | 22.83 | 37 | M14;M26 | |
| 7 | 21.61 | 36.50 | M9 | |
| 8 | 21.44 | 37 | M17;M20 | |
| 9 | 20.40 | 37 | M8 | |
| 10 | 20.30 | 37.50 | M2 | |
| 11 | 19.83 | 37.50 | M3 | |
| 12 | 18.96 | 37 | M7 | |
| 13 | 18.18 | 37 | M13 | |
| 14 | 17.89 | 37 | M6 | |
| 15 | 17.69 | 37 | M16 | |
| 16 | 17.48 | 37 | M4 | |
| 17 | 17.37 | 37 | M22 | |
| 18 | 16.90 | 37.50 | M10 | |
| 19 | 16.80 | 37 | M21 | |
| 20 | 16.52 | 37.50 | M15 | |
| 21 | 15.92 | 37.50 | M28;M31 | |
| 22 | 12.04 | 37.50 | M24 | |
| 23 | 11.57 | 37.50 | M27 | |
| Z | 26.23 | 36.50 | MZ |
| Measure | Value | Benchmark |
|---|---|---|
| EBP summary (haplotype 1) | 6.C.Q59 | 6.C.Q40 |
| Contig N50 length | 7.22 Mb | ≥ 1 Mb |
| Scaffold N50 length | 20.30 Mb | = chromosome N50 |
| Consensus quality (QV) | Haplotype 1: 59.3; haplotype 2: 59.1; combined: 59.2 | ≥ 40 |
|
| Haplotype 1: 75.72%; Haplotype 2: 75.66%; combined: 99.53% | ≥ 95% |
| BUSCO | C:97.4% [S:96.5%; D:0.9%]; F:0.4%; M:2.2%; n:5 286 | S > 90%; D < 5% |
| Percentage of assembly assigned to chromosomes | 98.72% | ≥ 90% |
- —Wellcome Trust
- —Swiss National Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLepidoptera: Biology and Taxonomy · Genomics and Phylogenetic Studies · Botany and Geology in Latin America and Caribbean
Species taxonomy
Eukaryota; Opisthokonta; Metazoa; Eumetazoa; Bilateria; Protostomia; Ecdysozoa; Panarthropoda; Arthropoda; Mandibulata; Pancrustacea; Hexapoda; Insecta; Dicondylia; Pterygota; Neoptera; Endopterygota; Amphiesmenoptera; Lepidoptera; Glossata; Neolepidoptera; Heteroneura; Ditrysia; Obtectomera; Papilionoidea; Lycaenidae; Polyommatinae; Cupido; Cupido osiris (Meigen, 1830) (NCBI:txid282382)
Background
The Osiris blue – Cupido osiris (Meigen, 1829) – is widespread in Europe and Central Asia ( Tolman & Lewington, 1997), ranging from the Iberian Peninsula to eastern Mongolia, including the southern part of France, Italy, the west and south of the Balkan Peninsula, Turkey, Kyrgyzstan and southern Russia. However, its distribution is highly fragmented, with large occurrence gaps, particularly in central Europe and the central Balkan Peninsula ( Kudrna et al., 2015; Tshikolovets, 2011).
The species shows strong sexual dimorphism, with males having a dark violet-blue upper surface, while females are brown. The latter is very similar to Cupido minimus (Füssli, 1775), C. lorquinii (Herrich-Schäffer, 1850) and Cyaniris semiargus (Rottemburg, 1775), and identification is not always easy. The underside of the wings is light grey with a large suffusion of blue at the base. It is adorned with small black eye spots.
The species is generally univoltine and adults are mainly seen from May to July, depending on altitude. A partial second generation is reported in the south of its range and at low altitudes. The species is found from the plains to altitudes above 2 000 m in the European Alps ( Ligue Suisse pour la Protection de la Nature, 1987). It colonises sunny mesophilic calcareous grasslands ( Van Swaay, 2002) and rocky roadsides, where sainfoins ( Onobrychis spp.), its main host plants, grow in abundance. The occasional use of bladder senna ( Colutea arborescens) is also reported ( Clarke, 2024; Lafranchis et al., 2015). Other suggested host plants ( Tshikolovets, 2011; Verity, 1943), including common kidney vetch ( Anthyllis vulneraria) and pea vine ( Lathyrus spp.), do not seem to be confirmed.
The eggs are laid individually on the inflorescences of the host plants. The caterpillars feed on the flowers and fruits. Being facultative myrmecophiles, they are often accompanied by ants of the genera Formica, Camponotus, Lasius and Plagiolepis ( Lafranchis et al., 2015). Caterpillars spend the late summer, autumn and winter on the ground or on their host plant and pupate only in spring, usually among dead leaves on the ground. Adults can easily be seen nectaring on flowering sainfoin.
Although the species is not globally threatened (LC according to Van Swaay et al., 2010) and may even be locally common, especially in the mountains, it has become very rare in its northern distribution range and is endangered in several countries, including Switzerland ( Wermeille et al., 2014), Austria ( Höttinger & Pennerstorfer, 2005) or is already extinct as in Germany ( Reinhardt & Bolz, 2011). The occupied habitats are often small and locally threatened by urbanisation and intensification of agricultural practices, including overfertilisation, overgrazing and excessive mowing. In the south, the abandonment of grasslands threatens the availability of its host plant and, consequently, the butterfly. Climate change represents an additional threat ( Obregón et al., 2016).
The reference genome for Cupido osiris provides an invaluable resource for studying the population structure of this species and for shedding light on the evolution of colour polymorphism in this group ( Hinojosa et al., 2020).
Methods
Sample acquisition
The specimen used for genome sequencing was an adult male Cupido osiris (specimen ID SAN28000112, ToLID ilCupOsir1; Figure 1), collected from Ormont-Dessus, Switzerland (latitude 46.3553, longitude 7.204; elevation 1 559 m) on 2023-06-28. The specimen was collected and identified by Andreas Sanchez (Info fauna).
Voucher photograph of the Cupido osiris (ilCupOsir1) specimen used for genome sequencing.
Nucleic acid extraction
Protocols for high molecular weight (HMW) DNA extraction developed at the Wellcome Sanger Institute (WSI) Tree of Life Core Laboratory are available on protocols.io ( Howard et al., 2025). The ilCupOsir1 sample was weighed and triaged to determine the appropriate extraction protocol. Tissue from the thorax was homogenised by powermashing using a PowerMasher II tissue disruptor. HMW DNA was extracted in the WSI Scientific Operations core using the Automated MagAttract v2 protocol. DNA was sheared into an average fragment size of 12–20 kb following the Megaruptor®3 for LI PacBio protocol. Sheared DNA was purified by automated SPRI (solid-phase reversible immobilisation). The concentration of the sheared and purified DNA was assessed using a Nanodrop spectrophotometer and Qubit Fluorometer using the Qubit dsDNA High Sensitivity Assay kit. Fragment size distribution was evaluated by running the sample on the FemtoPulse system. For this sample, the final post-shearing DNA had a Qubit concentration of 32.71 ng/μL and a yield of 1 537.37 ng, with a fragment size of 13.9 kb.
PacBio HiFi library preparation and sequencing
Library preparation and sequencing were performed at the WSI Scientific Operations core. Libraries were prepared using the SMRTbell Prep Kit 3.0 (Pacific Biosciences, California, USA), according to the manufacturer’s instructions. The kit includes reagents for end repair/A-tailing, adapter ligation, post-ligation SMRTbell bead clean-up, and nuclease treatment. Size selection and clean-up were performed using diluted AMPure PB beads (Pacific Biosciences). DNA concentration was quantified using a Qubit Fluorometer v4.0 (ThermoFisher Scientific) and the Qubit 1X dsDNA HS assay kit. Final library fragment size was assessed with the Agilent Femto Pulse Automated Pulsed Field CE Instrument (Agilent Technologies) using the gDNA 55 kb BAC analysis kit. The sample was sequenced on a Revio instrument (Pacific Biosciences). The prepared library was normalised to 2 nM, and 15 μL was used for making complexes. Primers were annealed and polymerases bound to generate circularised complexes, following the manufacturer’s instructions. Complexes were purified using 1.2X SMRTbell beads, then diluted to the Revio loading concentration (200–300 pM) and spiked with a Revio sequencing internal control. The sample was sequenced on a Revio 25M SMRT cell. The SMRT Link software (Pacific Biosciences), a web-based workflow manager, was used to configure and monitor the run and to carry out primary and secondary data analysis. Specimen details, sequencing platforms, and data yields are summarised in Table 1.
Hi-C
** Sample preparation and crosslinking **
The Hi-C sample was prepared from 20–50 mg of frozen tissue from the abdomen; head of the ilCupOsir1 sample using the Arima-HiC v2 kit (Arima Genomics). Following the manufacturer’s instructions, tissue was fixed and DNA crosslinked using TC buffer to a final formaldehyde concentration of 2%. The tissue was homogenised using the Diagnocine Power Masher-II. Crosslinked DNA was digested with a restriction enzyme master mix, biotinylated, and ligated. Clean-up was performed with SPRISelect beads before library preparation. DNA concentration was measured with the Qubit Fluorometer (Thermo Fisher Scientific) and Qubit HS Assay Kit. The biotinylation percentage was estimated using the Arima-HiC v2 QC beads.
** Hi-C library preparation and sequencing **
Biotinylated DNA constructs were fragmented using a Covaris E220 sonicator and size selected to 400–600 bp using SPRISelect beads. DNA was enriched with Arima-HiC v2 kit Enrichment beads. End repair, A-tailing, and adapter ligation were carried out with the NEBNext Ultra II DNA Library Prep Kit (New England Biolabs), following a modified protocol where library preparation occurs while DNA remains bound to the Enrichment beads. Library amplification was performed using KAPA HiFi HotStart mix and a custom Unique Dual Index (UDI) barcode set (Integrated DNA Technologies). Depending on sample concentration and biotinylation percentage determined at the crosslinking stage, libraries were amplified with 10–16 PCR cycles. Post-PCR clean-up was performed with SPRISelect beads. Libraries were quantified using the AccuClear Ultra High Sensitivity dsDNA Standards Assay Kit (Biotium) and a FLUOstar Omega plate reader (BMG Labtech). Prior to sequencing, libraries were normalised to 10 ng/μL. Normalised libraries were quantified again to create equimolar and/or weighted 2.8 nM pools. Pool concentrations were checked using the Agilent 4200 TapeStation (Agilent) with High Sensitivity D500 reagents before sequencing. Sequencing was performed using paired-end 150 bp reads on the Illumina NovaSeq X. Specimen details, sequencing platforms, and data yields are summarised in Table 1.
Genome assembly
Prior to assembly of the PacBio HiFi reads, a database of k-mer counts ( k = 31) was generated from the filtered reads using FastK. GenomeScope2 ( Ranallo-Benavidez et al., 2020) was used to analyse the k-mer frequency distributions, providing estimates of genome size, heterozygosity, and repeat content. The HiFi reads were assembled using Hifiasm in Hi-C phasing mode ( Cheng et al., 2021; Cheng et al., 2022), producing two haplotypes. Hi-C reads ( Rao et al., 2014) were mapped to the primary contigs using bwa-mem2 ( Vasimuddin et al., 2019). Contigs were further scaffolded with Hi-C data in YaHS ( Zhou et al., 2023), using the --break option for handling potential misassemblies. The scaffolded assemblies were evaluated using Gfastats ( Formenti et al., 2022), BUSCO ( Manni et al., 2021) and MERQURY.FK ( Rhie et al., 2020). The mitochondrial genome was assembled using MitoHiFi ( Uliano-Silva et al., 2023), which runs MitoFinder ( Allio et al., 2020) and uses these annotations to select the final mitochondrial contig and to ensure the general quality of the sequence.
Assembly curation
The assembly was decontaminated using the Assembly Screen for Cobionts and Contaminants ( ASCC) pipeline. TreeVal was used to generate the flat files and maps for use in curation. Manual curation was conducted primarily in PretextView and HiGlass ( Kerpedjiev et al., 2018). Scaffolds were visually inspected and corrected as described by Howe et al. (2021). Manual corrections included four breaks and ten joins. This reduced the scaffold count by 9.4%. The curation process is described at https://gitlab.com/wtsi-grit/rapid-curation. PretextSnapshot was used to generate a Hi-C contact map of the final assembly.
Assembly quality assessment
The Merqury.FK tool ( Rhie et al., 2020), run in a Singularity container ( Kurtzer et al., 2017), was used to evaluate k-mer completeness and assembly quality for both haplotypes using the k-mer database ( k = 31) computed prior to genome assembly. The analysis outputs included assembly QV scores and completeness statistics. The genome was analysed using the BlobToolKit pipeline, a Nextflow ( Di Tommaso et al., 2017) implementation of the earlier Snakemake version ( Challis et al., 2020). The pipeline aligns PacBio reads using minimap2 ( Li, 2018) and SAMtools ( Danecek et al., 2021) to generate coverage tracks. It runs BUSCO ( Manni et al., 2021) using lineages identified from the NCBI Taxonomy ( Schoch et al., 2020). For the three domain-level lineages, BUSCO genes are aligned to the UniProt Reference Proteomes database ( Bateman et al., 2023) using DIAMOND blastp ( Buchfink et al., 2021). The genome is divided into chunks based on the density of BUSCO genes from the closest taxonomic lineage, and each chunk is aligned to the UniProt Reference Proteomes database with DIAMOND blastx. Sequences without hits are chunked using seqtk and aligned to the NT database with blastn ( Altschul et al., 1990). The BlobToolKit suite consolidates all outputs into a blobdir for visualisation. The BlobToolKit pipeline was developed using nf-core tooling ( Ewels et al., 2020) and MultiQC ( Ewels et al., 2016), with containerisation through Docker ( Merkel, 2014) and Singularity ( Kurtzer et al., 2017). We painted Merian elements, which represent the 32 ancestral linkage groups in Lepidoptera ( Wright et al., 2024), along chromosomes using lep_busco_painter. Painting used BUSCO gene locations from the lepidoptera_odb10 set ( Kriventseva et al., 2019) and chromosome lengths from NCBI Datasets. Each complete BUSCO (including both single-copy and duplicated BUSCOs) was assigned to a Merian element using a reference database. Positions were coloured and plotted along chromosomes drawn to scale.
Genome sequence report
Sequence data
PacBio sequencing of the Cupido osiris specimen generated 27.78 Gb (gigabases) from 2.82 million reads, which were used to assemble the genome. GenomeScope2.0 analysis estimated the haploid genome size at 473.78 Mb, with a heterozygosity of 1.27% and repeat content of 39.23% ( Figure 2). These estimates guided expectations for the assembly. Based on the estimated genome size, the sequencing data provided approximately 57× coverage. Hi-C sequencing produced 190.11 Gb from 1 259.01 million reads, which were used to scaffold the assembly. Table 1 summarises the specimen and sequencing details.
Frequency distribution of k-mers generated using GenomeScope2.The plot shows observed and modelled k-mer spectra, providing estimates of genome size, heterozygosity, and repeat content based on unassembled sequencing reads.
Assembly statistics
The genome was assembled into two haplotypes using Hi-C phasing. Haplotype 1 was curated to chromosome level, while haplotype 2 was assembled to scaffold level. The final assembly has a total length of 481.43 Mb in 260 scaffolds, with 98 gaps, and a scaffold N50 of 20.3 Mb ( Table 2).
Most of the assembly sequence (98.72%) was assigned to 24 chromosomal-level scaffolds, representing 23 autosomes and the Z sex chromosome. These chromosome-level scaffolds, confirmed by Hi-C data, are named according to size ( Figure 3; Table 3). Chromosome painting with Merian elements illustrates the distribution of orthologues along chromosomes and highlights patterns of chromosomal evolution relative to Lepidopteran ancestral linkage groups ( Figure 4). Z chromosome identified based BUSCO gene painting with ancestral Merian elements ( Wright et al., 2024).
Hi-C contact map of the Cupido osiris genome assembly.Assembled chromosomes are shown in order of size and labelled along the axes, with a megabase scale shown below. The plot was generated using PretextSnapshot.
Table 3.: Chromosomal pseudomolecules in the haplotype 1 genome assembly of Cupido osiris ilCupOsir1.
Merian elements painted across chromosomes in the ilCupOsir1.hap1.1 assembly of Cupido osiris.Chromosomes are drawn to scale, with the positions of orthologues shown as coloured bars. Each orthologue is coloured by the Merian element that it belongs to. All orthologues which could be assigned to Merian elements are shown.
The mitochondrial genome was also assembled (length 15.36 kb, OZ218356.1). This sequence is included as a contig in the multifasta file of the genome submission and as a standalone record.
Assembly quality metrics
For haplotype 1, the estimated QV is 59.3, and for haplotype 2, 59.1. When the two haplotypes are combined, the assembly achieves an estimated QV of 59.2. The k-mer completeness is 75.72% for haplotype 1, 75.66% for haplotype 2, and 99.53% for the combined haplotypes ( Figure 5). BUSCO analysis using the lepidoptera_odb10 reference set ( n = 5 286) identified 97.4% of the expected gene set (single = 96.5%, duplicated = 0.9%) in haplotype 1. For haplotype 2, BUSCO analysis identified 97.6% of the expected gene set (single = 97.0%, duplicated = 0.5%).
Evaluation of k-mer completeness using MerquryFK.This plot illustrates the recovery of k-mers from the original read data in the final assemblies. The horizontal axis represents k-mer multiplicity, and the vertical axis shows the number of k-mers. The black curve represents k-mers that appear in the reads but are not assembled. The green curve (the homozygous peak) corresponds to k-mers shared by both haplotypes and the red and blue curves (the heterozygous peaks) show k-mers found only in one of the haplotypes.
The snail plot in Figure 6 summarises the scaffold length distribution and other assembly statistics for haplotype 1. The blob plot in Figure 7 shows the distribution of scaffolds by GC proportion and coverage for haplotype 1.
Assembly metrics for ilCupOsir1.hap1.1. The BlobToolKit snail plot provides an overview of assembly metrics and BUSCO gene completeness. The circumference represents the length of the whole genome sequence, and the main plot is divided into 1,000 bins around the circumference. The outermost blue tracks display the distribution of GC, AT, and N percentages across the bins. Scaffolds are arranged clockwise from longest to shortest and are depicted in dark grey. The longest scaffold is indicated by the red arc, and the deeper orange and pale orange arcs represent the N50 and N90 lengths. A light grey spiral at the centre shows the cumulative scaffold count on a logarithmic scale. A summary of complete, fragmented, duplicated, and missing BUSCO genes in the set is presented at the top right. An interactive version of this figure can be accessed on the BlobToolKit viewer.
BlobToolKit GC-coverage plot for ilCupOsir1.hap1.1.Blob plot showing sequence coverage (vertical axis) and GC content (horizontal axis). The circles represent scaffolds, with the size proportional to scaffold length and the colour representing phylum membership. The histograms along the axes display the total length of sequences distributed across different levels of coverage and GC content. An interactive version of this figure is available on the BlobToolKit viewer.
Table 4 lists the assembly metric benchmarks adapted from Rhie et al. (2021) and the Earth BioGenome Project Report on Assembly Standards September 2024. The EBP metric, calculated for the haplotype 1, is 6.C.Q59, meeting the recommended reference standard.
Table 4.: Earth Biogenome Project summary metrics for the Cupido osiris assembly.
Wellcome Sanger Institute – Legal and Governance
The materials that have contributed to this genome note have been supplied by a Tree of Life collaborator. The Wellcome Sanger Institute employs a process whereby due diligence is carried out proportionate to the nature of the materials themselves, and the circumstances under which they have been/are to be collected and provided for use. The purpose of this is to address and mitigate any potential legal and/or ethical implications of receipt and use of the materials as part of the research project, and to ensure that in doing so, we align with best practice wherever possible. The overarching areas of consideration are:
-Ethical review of provenance and sourcing of the material-Legality of collection, transfer and use (national and international).
Each transfer of samples is undertaken according to a Research Collaboration Agreement or Material Transfer Agreement entered into by the Tree of Life collaborator, Genome Research Limited (operating as the Wellcome Sanger Institute), and in some circumstances, other Tree of Life collaborators.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allio R Schomaker-Bastos A Romiguier J : Mito Finder: efficient automated large-scale extraction of mitogenomic data in target enrichment phylogenomics. Mol Ecol Resour. 2020;20(4):892–905. 10.1111/1755-0998.13160 32243090 PMC 7497042 · doi ↗ · pubmed ↗
- 2Altschul SF Gish W Miller W : Basic Local Alignment Search Tool. J Mol Biol. 1990;215(3):403–10. 10.1016/S 0022-2836(05)80360-2 2231712 · doi ↗ · pubmed ↗
- 3Bateman A Martin MJ Orchard S : Uni Prot: the Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023;51(D 1):D 523–D 531. 10.1093/nar/gkac 1052 36408920 PMC 9825514 · doi ↗ · pubmed ↗
- 4Buchfink B Reuter K Drost HG : Sensitive protein alignments at Tree-of-Life scale using DIAMOND. Nat Methods. 2021;18(4):366–368. 10.1038/s 41592-021-01101-x 33828273 PMC 8026399 · doi ↗ · pubmed ↗
- 5Challis R Richards E Rajan J : Blob Tool Kit – interactive quality assessment of genome assemblies. G 3 (Bethesda). 2020;10(4):1361–1374. 10.1534/g 3.119.400908 32071071 PMC 7144090 · doi ↗ · pubmed ↗
- 6Cheng H Concepcion GT Feng X : Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat Methods. 2021;18(2):170–175. 10.1038/s 41592-020-01056-5 33526886 PMC 7961889 · doi ↗ · pubmed ↗
- 7Cheng H Jarvis ED Fedrigo O : Haplotype-resolved assembly of diploid genomes without parental data. Nat Biotechnol. 2022;40(9):1332–1335. 10.1038/s 41587-022-01261-x 35332338 PMC 9464699 · doi ↗ · pubmed ↗
- 8Clarke HE : A checklist of European butterfly larval foodplants. Ecol Evol. 2024;14(1): e 10834. 10.1002/ece 3.10834 38192903 PMC 10771928 · doi ↗ · pubmed ↗