Precision Microbiome Modulation: Exploring Lactobacillus spp. as A Targeted Strategy for Type 2 Diabetes Management
Ann Sze Cheah, Ji Wei Tan, Faizul Jaafar, Wendy Wai Yeng Yeo

TL;DR
This paper explores how Lactobacillus bacteria can be used in personalized treatments to help manage type 2 diabetes more effectively.
Contribution
The paper proposes integrating Lactobacillus spp. into precision medicine for T2DM using multi-omics and AI.
Findings
Lactobacillus spp. improve insulin resistance through glucose metabolism and anti-inflammatory effects.
Strain-specific survivability and safety issues hinder clinical use of Lactobacillus spp.
Personalized approaches using microbiome and genetic data may optimize Lactobacillus-based therapies.
Abstract
The International Diabetes Federation projects a prevalence of 783.2 million cases of type 2 diabetes mellitus (T2DM) by 2045. The escalation in healthcare expenditure and adverse effects of current drugs necessitate a clinically effective and low-cost alternative therapeutic option with minimal health complications. Lactobacillus spp., a well-studied gut probiotic has shown promising antidiabetic effects against T2DM in several randomized controlled trials. However, the exact mechanisms of Lactobacillus spp. in regulating the glycaemic profile are not well elucidated, limiting the optimization of probiotic-based interventions. Therefore, this paper aims to explore the various antidiabetic mechanisms of Lactobacillus spp., with a focus on their integration into precision medicine approaches. Lactobacillus spp. supplementation alleviates insulin resistance in T2DM by modulating glucose…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
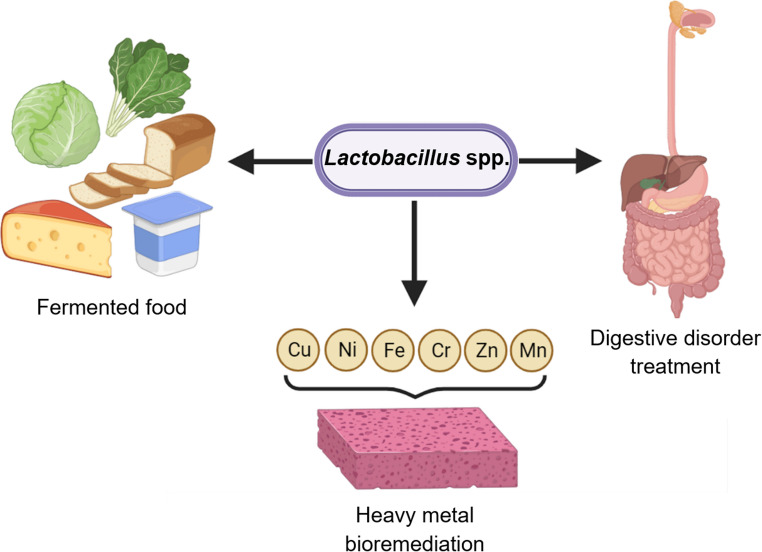 Figure 1
Figure 1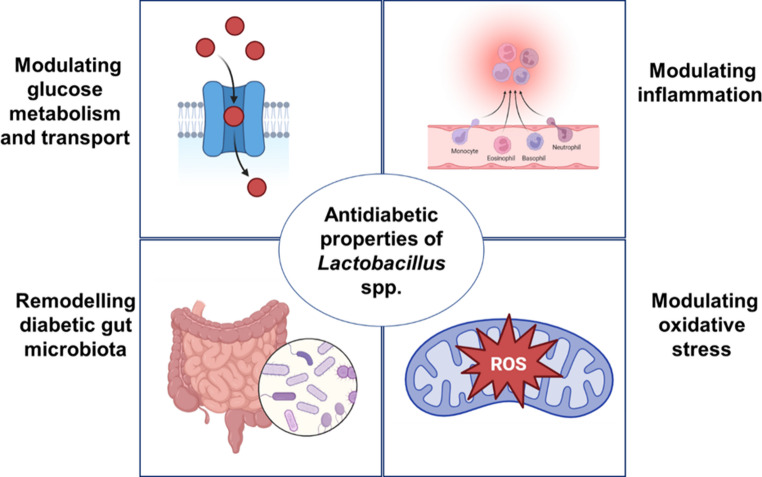 Figure 2
Figure 2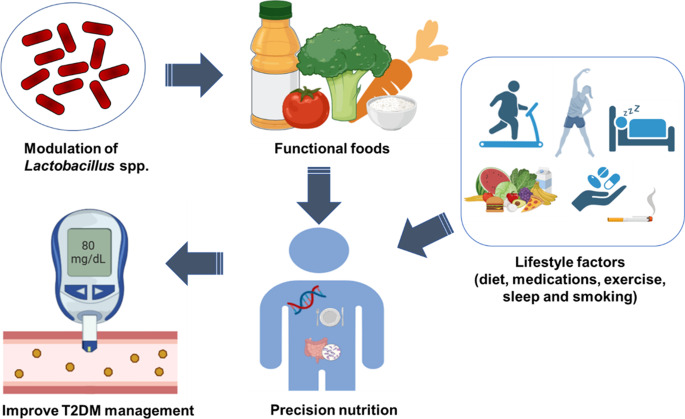 Figure 3
Figure 3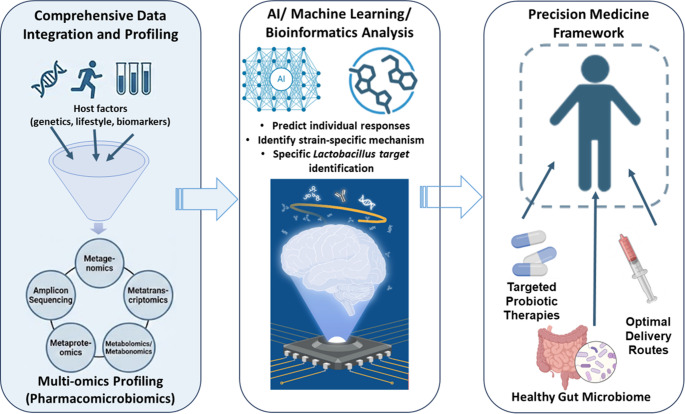 Figure 4
Figure 4- —Monash University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Probiotics and Fermented Foods · Gastrointestinal motility and disorders
Introduction
Type 2 diabetes mellitus (T2DM) is a metabolic disorder that manifests as hyperglycaemia, arising from insulin resistance and impaired pancreatic beta cell function [1]. Common risk factors for T2DM include genetic predisposition, a sedentary lifestyle and high-fat diets [2]. Poor management of T2DM results in persistent hyperglycaemia and leads to various adverse complications such as neuropathy, nephropathy and retinopathy [3]. Furthermore, uncontrolled hyperglycaemia weakens the immune system, making diabetic patients more prone to infections such as skin, urinary tract, respiratory and diabetic foot infections, often caused by pathogens such as E. coli and Klebsiella [4]. Studies revealed that inflammatory response plays a pivotal role in the development of insulin resistance, a hallmark of T2DM [5]. It is suggested that inflammation triggers the production of pro-inflammatory molecules including cytokines and adipocytokines through various pathways, including oxidative and metabolic stress which interfere with insulin signalling pathways, exacerbating insulin resistance and hyperglycaemia [5, 6].
The International Diabetes Federation estimated a global diabetes prevalence of 536.6 million in 2021 and an expected 46% increase to 783.2 million in 2045, with middle-income countries being the most affected, attributing to substantial population growth [7]. Additionally, the global healthcare expenditure for the management of diabetes and its associated complications, projected to surpass one trillion USD by 2045, poses a substantial financial burden especially to individuals from low-middle income countries which account for approximately 80% of the patient population [7]. The anticipated prominent rise in T2DM prevalence in middle-income countries and concomitant economic burden manifest the need for an alternative treatment. The inter-individual variability in drug response can implicate adverse drug reactions, paradoxically necessitating further prolonged treatment and imposing significant health and economic burdens [8].
Currently, various drug options with different mechanisms of action are available for T2DM treatment. Insulin sensitizers, including biguanides and thiazolidinediones, promote insulin sensitivity, whereas both insulin secretagogues, such as sulfonylureas and meglitinides, and incretin-based therapy, such as dipeptidyl peptidase-IV (DPP-IV) inhibitors and glucagon-like peptide-1 (GLP-1) receptor agonists, stimulate insulin production [9]. Furthermore, α-glucosidase inhibitors reduce intestinal glucose absorption, whereas sodium-glucose cotransporter-2 (SGLT2) inhibitors inhibit renal glucose reabsorption [9]. However, these pharmaceutical agents have low efficacy, limited tolerability and are associated with gastrointestinal symptoms such as diarrhoea and vomiting, weight gain as well as hypoglycaemia [10].
Recent advances in genomics have ushered in a new era of precision medicine, which tailors disease prevention and treatment based on an individual’s genomic profile, lifestyle and environmental exposures [11]. One area of growing interests in the search for alternative treatment for T2DM is the emergence of pharmacomicrobiomics, which explores the influence of human microbiome variation in drug response and disposition that provides insights into the complex host-environment-human microbiome interactions [12–14]. In this context, probiotics such as Lactobacillus spp. represent a promising avenue for precision microbiome modulation to improve glycaemic control and metabolic outcomes in T2DM.
The gut microbiota is crucial in modulating host metabolism. Thus, gut dysbiosis is implicated in the pathogenesis of T2DM and other metabolic disorders as it significantly influences the regulation of inflammation, energy homeostasis, gut barrier function, glucose and lipid metabolism, insulin sensitivity as well as dietary interactions [15]. Dysbiosis is defined as the compositional imbalance of resident commensal communities, alterations in bacterial metabolic activities, or shifting of the bacterial distribution within the gut [16].The T2DM microbiota profile is characterized by an increased relative abundance of opportunistic pathogens and a decreased relative abundance of butyrate-producing bacteria, which further signifies the link between gut dysbiosis and T2DM development [17]. This paradigm shift has generated a new perspective in T2DM treatment: probiotics as a novel therapeutic alternative for a more nuanced approach to treatment through precision medicine.
Probiotics are live bacteria that benefit the host’s health when administered sufficiently [18] Unlike conventional drug regimens, probiotics are generally safer, non-invasive, more accessible, inexpensive and associated with fewer side effects [19–21]. Long-term probiotic administration also establishes a stable colonization to provide an enduring effect [22]. Recent meta-analysis has corroborated the positive role of probiotics in glycaemic control in human trials, as supported by the clinically significant decrease in fasting insulin, fasting blood glucose (FBG), and homeostasis model of assessment of insulin resistance [22, 23]. Among the probiotics, Lactobacillus species are the most studied and widely commercialized [24]. Members of the Lactobacillus genus are vital probiotic bacteria in the gut microbiome and have a prominent role as a therapeutic approach for liver diseases and cardiovascular-related diseases in clinical studies [25–27].
Multiple randomized placebo-controlled trials have demonstrated the clinical effectiveness of Lactobacillus spp. in regulating the glycaemic profile in T2DM patients [1, 28–31]. However, the mechanisms involved have not been defined clearly, which impedes probiotic therapy development. Therefore, this review aims to investigate the antidiabetic mechanisms of Lactobacillus spp. as a targeted and personalized strategy to treat T2DM. Additionally, this paper will evaluate the potential challenges associated with Lactobacillus spp. supplementation and suggest methods to improve the application feasibility.
Research Methodology
This narrative review was carried out with a structured search strategy, though not fully systematic. We conducted searches in several scientific databases, including PubMed, Scopus and Web of Science, along with other relevant sources from database inception to 1 June 2024, with the final search conducted on 1 December 2025. The search terms included key concepts related to probiotics and diabetes, such as “Lactobacillus,” “probiotics,” “gut microbiota,” “type 2 diabetes mellitus,” “insulin resistance,” “fermented foods,” and “precision medicine.”
We included original research articles involving human or animal models that met the following criteria: (i) the study investigated Lactobacillus spp. or products enriched with Lactobacillus as the primary intervention or exposure; and (ii) it reported outcomes related to glycemic control, insulin resistance or gut microbiota composition in the context of T2DM. Both clinical trials and in vivo studies were considered. Only studies published in English were included. Editorials, conference abstracts and studies not involving Lactobacillus or not relevant to T2DM-related outcomes were excluded.
Targeted Lactobacillus spp. In T2DM
The Lactobacillus genus, the lactic acid bacteria, comprises gram-positive bacteria characterized by their rod shape, non-spore-forming nature, and facultative anaerobic capacity [32]. Lactobacillus constitutes the normal flora in the gastrointestinal tract, oropharynx, and female genitourinary tract [33]. Lactobacillus are present in faecal samples either as transiently derived allochthonous strains or native autochthonous strains, accounting for approximately 0.01 to 0.06% of all faecal bacterial species in adults [27, 34, 35]. Lactobacillus spp. possess the inherent metabolic capacity to produce lactic acid upon endogenous carbohydrate metabolism and thus, are traditionally used for food fermentation [34]. Probiotic supplements and food products fortified with Lactobacillus spp. have demonstrated effectiveness in ameliorating digestive disorders such as lactose intolerance, irritable bowel syndrome, chronic diarrhoea and functional constipation [36–39]. Moreover, Lactobacillus have been exploited for their bioremediation potential. Lactobacillus fermentum (L. fermentum) and Lactobacillus plantarum (L. plantarum) strains demonstrate resistance to and rapid biosorption against heavy metals such as nickel in various sources, including aquatic waters, industrial wastewater effluents, and soil [40–42]. Figure 1 summarizes the current applications of Lactobacillus spp.Fig. 1. Current applications of Lactobacillus spp. (Created with BioRender.com)
Numerous clinical studies have compared the differential gut microbiota profile between T2DM patients and healthy controls from various countries. In a Southern Chinese population, a higher abundance of total Lactobacillus, including Lactobacillus acidophilus (L. acidophilus), Lactobacillus bulgaricus (L. bulgaricus), and Lactobacillus rhamnosus (L. rhamnosus), was reported in age- and gender-adjusted T2DM patients [43]. This trend has been consistently observed in multiple populations, in either newly diagnosed or long-standing diabetic patients as summarized in Table 1 [44–51]. A study reported a significantly higher faecal Lactobacillus count in diabetic patients, but the significant effect relationship was nullified after further adjustment for fibre intake [48]. A factor for this enrichment is the abnormally high glucose availability that provides a stimulatory environment for the metabolically flexible Lactobacillus [46].Table 1. Clinical and observational studies reporting higher Lactobacillus spp. abundance in individuals with type 2 diabetes mellitus (T2DM) compared with non-diabetic controls. The table summarizes the targeted Lactobacillus abundance, as indicated by the higher relative abundance in T2DM patients compared to healthy controls based on the sample sizes, study populations, exposure to treatment regimens, main outcomes and sample sourcesTargetSample sizeStudy populationRecipient of treatment regimensRelative abundanceMain outcomesSample sourceReferenceTotal LactobacillusT2DM: 50Control: 30Southern ChinaOnly a portion of subjects (not inclusion criteria)IncreasedHigher abundance associated with T2DMFaecal [43]Total LactobacillusT2DM: 50Control: 50JapanOnly a portion of subjects (not inclusion criteria)IncreasedDysbiosis in T2DMFaecal [44] Lactobacillus T2DM: 40Control: 13ItalyYesIncreasedAltered microbiota in T2DMFaecal [45] Lactobacillus T2DM: 16Control: 19IndiaYes (known diabetics)IncreasedCorrelated with hyperglycaemiaFaecal [46] L. acidophilus ATCC 4356 (standard strain)T2DM: 18Control: 18IranNot statedIncreasedHigher abundance in T2DMFaecal [47] Lactobacillus gasseri DSM 20243 T (standard strainT2DM: 50Control: 50TaiwanNo (newly diagnosed diabetics)Initially increased; lost after fibre adjustmentDiet strongly modified abundanceFaecal [48] Lactobacillus groupT2DM: 49Control: 40Not mentionedOnly a portion of subjects (not inclusion criteria)IncreasedGenus-level expansion in T2DMFaecal [49] Lactobacillus T2DM: 11Control: 35IndiaNo (newly diagnosed diabetics)IncreasedHigher abundance in early diseaseFaecal [50] Lactobacillus T2DM: 87Control: 49United States (T2DM); not mentioned for control groupYesIncreasedAltered metabolic phenotypeUrine [51]
While the consensus suggests a positive correlation between Lactobacillus spp. and T2DM, contradicting findings have also been reported. A recent clinical study suggested that low detection frequencies of Lactobacillus spp. in faecal samples were correlated with diabetes development [52]. In a Romanian cohort, Lactobacillus species were significantly lower in the T2DM group [53]. Similarly, L. acidophilus, L. salivaricus, and L. fermentum were significantly lower in stool samples of Iranian T2DM patients than in healthy subjects [54]. Significantly lower stool L. acidophilus count was also detected in T2DM patients from Egypt [55]. Another Romanian study demonstrated a lower abundance of L. acidophilus, L. plantarum, and Lactobacillus reuteri (L. reuteri) in T2DM subjects [56]. Yet, the same paper determined no significant changes between the diabetic and control groups in four other Lactobacillus subgroups: L. fermentum,* Lactobacillus salivaricus* (L. salivaricus), Lactobacillus johnsonii (L. johnsonii), and Lactobacillus ruminis (L. ruminis) [56].
The gut microbiota encompasses a diverse community of microorganisms, typically the members of the genus Lactobacillus, which are the most important probiotic bacteria that live in symbiosis with their host [27]. The interactions between Lactobacillus spp. and their hosts are greatly influenced by diet, genetics and lifestyle. This symbiotic relationship is crucial to maintain the development of the metabolic system [57], which contributes to the unique microbiome in each person’s gut. Sedighi et al. (2017) revealed higher levels of Lactobacillus were found in T2DM patients as compared to the healthy group. This observation agreed with numerous studies found in Table 1. These findings may be valuable for devising strategies to control T2DM by modifying the gut microbiota, suggesting the potential of leveraging precision medicine based on targeting Lactobacillus spp. in the treatment of diabetes.
Together, these observations support the concept of precision microbiome modulation, whereby Lactobacillus-based interventions are tailored to an individual’s baseline gut microbiota, metabolic phenotype, dietary intake and medication exposure instead of following uniform response pattern. The heterogeneous patterns of Lactobacillus abundance observed across populations (Table 1) suggest that probiotic efficacy is context-dependent and may reflect adaptive microbial responses to host glucose availability, fibre intake as well as pharmacological treatments.
Heterogeneity and Confounding Factors in Lactobacillus Abundance
The inconsistent associations between Lactobacillus and T2DM across studies are likely driven by methodological and biological heterogeneity. Several other factors may contribute to the discrepancy in these findings, namely the small sample size, different inclusion criteria of the study and heterogeneity of the study population. In addition, the use of 16 S rRNA sequencing for the detection of a reference strain may not fully represent the vast microbiome composition and abundance of the genus. Differences between metataxonomics (16 S rRNA gene sequencing) and metagenomics (whole shotgun metagenomic sequencing) may produce different levels of taxonomic resolution and different functional interpretation of microbiome data [58]. As a result, the illustrated trends can be underestimated or overestimated since the host metabolism may be related to a dysbiotic shift in gut microbiota instead of alterations of a single species [48].
Besides that, clinical cohorts often exhibit significant heterogeneity in diet, fibre intake as well as the use of antidiabetic medication such as metformin, which are known to independently modulate gut microbiota composition and function [59, 60]. This complexity is compounded by the substantial strain-level functional diversity within the Lactobacillus spp. Consequently, the species-level abundance does not reliably reflect shared metabolic or immunomodulatory traits. Notably, two strains within the same species may differ markedly in their in short-chain fatty acids (SCFA) production profiles and immune effects [61]. Furthermore, geographical location differences in lifestyle, early-life microbial exposures and endemic pathogen burden contribute to inter-population heterogeneity, thereby complicating the generalization of findings [62, 63]. Hence, addressing these limitations requires standardization of the subject selection criteria, recruitment of participants from diverse geographic regions and application high-resolution shotgun sequencing to improve metagenomics analyses. Moreover, prospective longitudinal studies may be more appropriate to elucidate the causal relationship between microbiome alterations and disease development.
Therefore, it is vital to investigate the potential factors which causes the diversity and development of the Lactobacillus spp [64]. One of the main factors is the dietary interventions, as human not only provide habitat for their living but also provide them with food. The ingested food which is utilized by Lactobacillus spp. may contribute to their growth, and thereby optimizing their impact on metabolic health, or vice versa. It is of evident in a recent systematic review and meta-analysis of randomized controlled trials on symbiotic supplementation, revealing its effects on glucose metabolism among diabetes individual [65]. This study reported an improvement in fasting plasma glucose, insulin concentrations and homeostasis model assessment of insulin resistance (HOMA-IR) in diabetes patients using the symbiotic supplementation, which is a mixture of gut bacteria (probiotics) and substances which can only be metabolised by gut microbiome (prebiotics).
Nonetheless, all these clinical studies explicitly target microbiome to obtain clinical results and improve clinical efficacy. Given that clinical samples were mainly from faecal, this can be utilised as diagnostic or prognostic tools [66]. Hence, assessment of the gut microbiome’s composition, quantity, and the relative abundance of its metabolites could serve as a distinctive predictive marker for different individuals linking to health conditions and behavioural patterns [67–69]. These emergent findings suggest that the analysis of microbiome characteristics may offer valuable insights into personalized medicine and contribute to more effective diagnostics and prognostics for individual with diabetes.
Antidiabetic Properties of Lactobacillus Spp
Role of Lactobacillus spp. in Modulating Glucose Metabolism and Transport
Lactobacillus isolates can exert therapeutic effects against T2DM by regulating the expression and activity of glucose metabolism enzymes and transporters. Upon food intake, starches are hydrolyzed to disaccharides and oligosaccharides by α-amylase before subsequent conversion by α-glucosidase into monosaccharides [70]. Therefore, inhibiting these enzymes limits carbohydrate absorption to enable better glycaemic control, which would be reflected by a proportional decrease in blood glucose levels. From recent findings, L. plantarum strains MG4229, MG4296, and MG5025, Lacticaseibacillus paracasei (L. paracasei) MG5012, Lactobacillus sakei Probio65, and L. plantarum Probio-093 exhibited potent inhibition to the enzymatic activities of α-glucosidase and α-amylase, which were on par with acarbose [70, 71].
The glucagon signalling pathway is also crucial for the regulation of glucose metabolism. Glucagon binds and activates glucagon receptor (GCGR), which activates the downstream mediators heterotrimeric Gs protein alpha-subunit (Gnas), cAMP-dependent protein kinase (PKA), and CREB-regulated transcriptional coactivator 2 (CRTC2) [72]. CRTC2 subsequently promotes the transcriptional activity of CREB to upregulate the expression of phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase), the rate-limiting enzymes of hepatic gluconeogenesis [73]. T2DM mice treated with L. rhamnosus LRa05 displayed diminished expression of several key mediators of the hepatic glucagon signalling pathway, including glucagon, GCGR, Gnas, PKA, CRTC2, PEPCK, and G6Pase [74].
Similarly, Lactobacillus casei (L. casei) LC89 downregulated the expression of GCGR, Gnas, PKA, CRTC2, PEPCK, and G6Pase in diabetic rats [75]. L. plantarum HAC01 supplementation to T2DM mice downregulated PEPCK and G6Pase [76]. Inhibition of the glucagon signalling pathway and gluconeogenesis decrease endogenous glucose production and reduce FBG [75]. L. acidophilus KLDS1.0901 and KLDS1.1003 promoted hepatic glycogen synthesis by downregulating glycogen synthase kinase-3 beta (GSK-3β) expression, which also lowered the FBG in T2DM mice [77].
L. plantarum PCS26 significantly lowered glucose transporter 2 (GLUT-2) protein expression in Caco-2 cells, decreasing the rate of transepithelial glucose transport [78]. L. reuteri GL-104 and L. salivarius AP-32 also inhibited hexose uptake and demonstrated significant hypoglycaemic effects in Caco-2 cells by promoting high monosaccharide consumption efficiency and lowering the expression of the intestinal hexose transporter SGLT1 [79]. Similarly, L. plantarum inhibited SGLT2 expression and was associated with improved renal insulin sensitivity [80].
Overall, these studies show that selected Lactobacillus strains may improve glycaemic control through several mechanism. They may reduce carbohydrate breakdown, lower hepatic gluconeogenesis and limit glucose transportation in the gut. Together, these effects influence glucose absorption and endogenous glucose production, which indicate that probiotics act at several metabolic checkpoints to regulate blood glucose levels.
Role of Lactobacillus spp. in Modulating Inflammation
Hyperglycaemia is known to invoke an inflammatory response by stimulating the secretion of corresponding cytokines [81]. Likewise, a study has suggested the role of inflammatory mediators in insulin resistance development and T2DM pathogenesis [82]. A study in the Kashmiri population reported higher serum levels of tumour necrosis factor-alpha (TNF-α), C-reactive protein (CRP), and interleukin-6 (IL-6) but lower IL-10 concentrations in T2DM patients [82]. The elevated CRP, TNF-α, and IL-6 levels were strongly associated with insulin resistance in diabetic patients from Faisalabad [83]. A study in a Korean cohort reported similar findings, where CRP was significantly associated with incident T2DM irrespective of gender [84]. Meanwhile, CRP is likely a hyperglycaemic by product rather than an etiological biomarker of T2DM [85].
The inflammatory cytokines TNF-α, IL-1β, and IL-6 play significant roles in impairing insulin signalling, contributing to the development and progression of T2DM. A study has shown that TNF-α phosphorylates and inactivates the serine residue of insulin receptor substrate 1, reducing GLUT-4 translocation and impairing the insulin signalling pathway [86]. TNF-α and IL-1β induce the expression of IL-6, which, at high concentrations, stimulates the expression and activity of insulin-degrading enzyme to promote insulin clearance [87–89]. Additionally, TNF-α and IL-1β induce inflammation in peripheral tissues, resulting in the chemotactic invasion and infiltration of immune cells that compromise the structural integrity of these target tissues [90]. Overall, addressing the inflammatory response may ameliorate insulin resistance and hinder T2DM progression.
Herein, the anti-inflammatory potential of Lactobacillus spp. in ameliorating the pathological state of T2DM was observed in different in vivo models. L. casei CCFM419 significantly decreased serum levels of TNF-α and IL-6 but not IL-10 in high-fat and streptozotocin-induced C57BL/6J mice [91]. L. casei CCFM419 increased acetic acid and total SCFA, especially butyrate [91]. SCFAs have been postulated to actuate free fatty acid receptors 2 and 3 to initiate inhibitory signalling cascades against inflammation and inhibit histone deacetylase to reduce transcription of pro-inflammatory genes [92]. Meanwhile, Lactobacillus gasseri (L. gasseri) CKCC1913 reduced the expression of IL-6 and TNF-α in high-fat diet-induced insulin-resistant C57BL/6J mice [93]. In a T2DM zebrafish model, L. rhamnosus GG supplementation downregulated the relative expressions of IL-1β and TNF-α, which correlated positively to blood glucose levels [94].
In streptozotocin (STZ)-induced diabetic rats, L. rhamnosus Hao9 and L. fermentum MCC2759 and MCC2760 lowered the serum concentration and gene expression levels of IL-1β, IL-6, and TNF-α and stimulated IL-10 expression [95, 96]. L. fermentum MCC2759 and MCC2760 also reversed hepatic inflammatory infiltration and steatosis formation and normalized glomerular and intestinal integrity [95]. The immunomodulatory function of these strains was postulated to be mediated by modulating the gut microbiota, which is associated with reduced lipopolysaccharide production and improved gut barrier function [95]. It is suggested that IL-10 suppresses inflammatory cytokine generation and improves insulin signalling and glucose metabolism in skeletal muscle [97].
Meanwhile, L. rhamnosus Hao9 restores intestinal barrier integrity, preventing the vascular leakage of lipopolysaccharides and pro-inflammatory cytokines [96]. Supplementing high fructose-fed rats with L. plantarum significantly decreased renal tissue levels of IL-1β, IL-6, IL-10, and TNF-α but not nuclear factor kappa B (NF-κB) [80]. On the other hand, the anti-inflammatory effect of Lactobacillus helveticus (L. helveticus) was limited to the decrease in renal TNF-α level [80].
Another study has shown that the heat-killed L. plantarum L-137 attenuated systemic inflammation by lowering cytokine expression and improved insulin signalling by increasing protein kinase B (Akt) phosphorylation in the adipose tissue of obese rats [98]. Besides this, L. plantarum LRCC5310 and LRCC5314 alleviated insulin resistance in T2DM mice under chronic cold stress by markedly lowering TNF-α, IL-6, and C-C motif chemokine ligand 2 (CCL2) expression, thereby suppressing the pro-inflammatory activity [99]. Meanwhile, L. paracasei HII01 significantly increased Akt phosphorylation and GLUT-4 expression and improved insulin signalling by reducing systemic inflammation biomarkers TNF-α and NF-kB in diabetic rats [100]. L. gasseri CKCC1913 increased glucose tolerance and reduced the expression of TNF-α and IL-6 [93].
On the other hands, L. acidophilus KLDS1.1003 and KLDS1.0901 ablated the levels of pro-inflammation cytokines TNF-α, IL-8, and IL-1β in the liver and colon of T2DM mice [77]. Another study demonstrated that L. fermentum MCC3216 significantly lowered IL-6 and elevated IL-10 levels in high fructose-fed T2DM mice [101]. Similarly, L. fermentum TKSN041 increased IL-10 and decreased IL-1β and IL-6 in STZ-induced T2DM rats [102]. The strain also upregulated NFκB-inhibitor-α (IκB-α) expression to prevent the activation of NFκB-p65, resulting in lower TNF-α levels [102]. L. casei CCFM0412 protected against the pathological inflammatory state induced in T2DM mice, as exhibited by the lowered TNF-α and higher IL-10 levels [103].
Hence, based on these findings as shown in Table 2, Lactobacillus spp. may have the potential to improve the pathological state of T2DM by modulating host’s immune response via regulation of pro- and anti-inflammatory cytokine production. Altering the balance of these anti-inflammatory profiles may help to enhance insulin signalling pathway and glucose metabolism, preserving β-cell integrity as well as ameliorate insulin resistance to better control of glucose levels. This immunomodulatory mechanism highlights the potential of strain-specific Lactobacillus formulations as adjunctive therapies in the personalized management of T2DM, which reduce chronic low-grade inflammation associated with metabolic dysfunction. However, these findings are based on small-scale animal experiments which require further research involving clinical studies in humans to confirm their relevance and effectiveness in human populations.Table 2. Summary of experimental studies investigate the role of Lactobacillus spp. In modulating inflammation in metabolic disease models relevant to T2DM. The table highlights strain-specific effects on inflammatory cytokines, signaling pathways and metabolic outcomes across in vivo and in vitro modelsStrainModelDose, duration and route of administrationCytokines AffectedKey FindingsReferences L. casei CCFM419HFD + STZ-induced mice1 × 10⁹CFU/mL;12 weeks; intragastric↓ TNF-α, ↓ IL-6, ↔ IL-10Increased SCFAs (esp. butyrate); anti-inflammatory effect via SCFA pathways [91] L. gasseri CKCC1913HFD-induced mice1 × 10⁹ CFU/mL;6 weeks;oral gavage↓ TNF-α, ↓ IL-6Improved glucose tolerance and reduced inflammation [93] L. rhamnosus GGT2DM zebrafishDose not reported;10 days;oral administration↓ TNF-α, ↓ IL-1βCytokine reduction correlated with lower blood glucose [94] L. fermentum MCC2759/2760STZ-induced rats10⁹ CFU/mL;4 weeks;intragastric↓ TNF-α, ↓ IL-6, ↓ IL-1β, ↑ IL-10Reversed hepatic inflammation, improved gut barrier integrity [95] L. plantarum, L. helveticusHigh fructose-fed rats10⁹ CFU per 100 g body weight;6 weeks;gastric gavage↓ IL-1β, ↓ IL-6, ↓ IL-10, ↓ TNF-α (limited for L. helveticus)Anti-inflammatory effects; L. helveticus effect limited to TNF-α [80] Heat-killed L. plantarum L-137Obese rats2 mg/kg/day (low dose) or 75 mg/kg/day (high dose);4 weeks;oral gavage↓ Systemic cytokine expressionImproved insulin signalling via ↑ Akt phosphorylation [98] L. plantarum LRCC5310/5314Cold-stressed T2DM miceL. plantarum LRCC5310: 1 × 10⁸ CFU or 1 × 10¹⁰ CFUL. plantarum LRCC5314: 1 × 10⁸ CFU or 1 × 10¹⁰ CFU;12 weeks;oral administration↓ TNF-α, ↓ IL-6, ↓ CCL2Relieved insulin resistance by suppressing inflammation [99] L. paracasei HII01Diabetic rats1 × 10^8^ CFU/day;12 weeks;oral administration↓ TNF-α, ↓ NF-κB↑ Akt phosphorylation and GLUT-4 expression, improved insulin signalling [100] L. acidophilus KLDS1.1003/0901T2DM mice1 × 10⁹ CFU/day;6 weeks;oral gavage↓ TNF-α, ↓ IL-1β, ↓ IL-8Reduced inflammation in liver and colon [77] L. fermentum MCC3216High fructose-fed mice2.0 × 10⁸ CFU/mL;8 weeks;oral↓ IL-6, ↑ IL-10Balanced inflammatory response [101] L. fermentum TKSN041STZ-induced rats1.0 × 10⁹ CFU/mL;13 weeks;oral gavage↓ IL-1β, ↓ IL-6, ↑ IL-10, ↓ TNF-αUpregulated IκB-α to block NF-κB activation [102] L. casei CCFM0412T2DM mice1 × 10⁹ CFU/day;12 weeks;oral administration↓ TNF-α, ↑ IL-10Protected against pathological inflammation [103]
Collectively, the studies summarized in Table 2 suggest that selected Lactobacillus strains can attenuate T2DM-related inflammation by shifting the balance away from pro-inflammatory mediators. This is crucial to glycaemic control because chronic low-grade inflammation disrupts insulin signalling and contributes to systemic insulin resistance. This immunomodulatory mechanism highlights the potential of strain-specific Lactobacillus formulations as adjunctive therapies in the personalized management of T2DM, highlighting the importance of precision microbiome modulation rather than generalized probiotic use.
Role of Lactobacillus spp. in Regulating Oxidative Stress
In T2DM, abnormal hyperglycaemia leads to reactive oxygen species (ROS) overproduction, causing oxidative stress via multiple signalling pathway, including the advanced glycation end products pathway, hexosamine pathway, polyol pathway, and protein kinase C pathway [104]. Increased oxidative stress contributes to impaired glucose metabolism, potentially contributing to the development of T2DM, due to the imbalance of ROS generation and scavenging [104, 105].
Notably, the T2DM patients show higher malondialdehyde (MDA) levels, a marker of lipid damage and superoxide dismutase (SOD) activity, coupled with diminished glutathione (GSH) content and other antioxidant enzyme activity [106, 107]. The elevated total SOD activity alone as an adaptive response does not sufficiently protect against the substantial increase in oxidative stress-induced ROS generation [106]. Pancreatic beta cells are particularly susceptible to oxidative damage due to their limited antioxidant capacity and low antioxidant enzyme content, which can lead to beta cell failure [108]. Furthermore, ROS also inhibits GLUT-4 translocation and hinders insulin-stimulated glucose uptake by activating c-Jun N-terminal kinases (JNKs), suppressing the phosphatidylinositol 3-kinase (PI3K)/Akt signalling pathway, or oxidizing proteins in the insulin-signalling pathway [109–111].
Numerous studies indicate that administering Lactobacillus spp. exerts antioxidant effects in animal models, thereby delaying diabetes progression. Recent studies showed that L. paracasei L14 and NL41 substantially increased the levels of ROS-scavenging enzymes catalase, glutathione peroxidase (GPX), and SOD as well as reduced MDA concentration, restoring most parameters to baseline levels [112, 113]. On the other hand, L. paracasei isolated from Malaysian water kefir grains suppressed the STZ-induced increase in MDA levels and improved glucose tolerance in T2DM mice [114].
Compared to other isolates, Lactobacillus isolates from these grains possessed higher levels of antioxidant indicators, including high free radical scavenging activity [115]. The antioxidant strength is primarily linked to the high total phenolic and flavonoid content [115]. Lactobacillus spp. can increase the bioavailability of phenolic and flavonoid molecules by acting as hydrogen donors to deactivate free radicals [116]. Another finding reported that L. casei CCFM0412 promoted hepatic GSH and SOD levels while reducing MDA and ROS [103]. GSH is a nonenzymatic antioxidant, acting directly to quench free radicals or indirectly as the reducing agent for GPX-catalyzed reactions [117].
L. rhamnosus Hao9 promoted the hepatic antioxidant capacity, exemplified by the elevated levels of catalase and SOD in streptozotocin-high fat diet-induced T2DM mice [96]. Meanwhile, L. rhamnosus LRa05 exerted comparable effects to metformin in increasing catalase and GSH levels while reducing MDA levels in the liver of T2DM mice [74]. However, unlike L. rhamnosus Hao9, L. rhamnosus LRa05 did not significantly increase hepatic SOD levels, which may be attributed to the differential antioxidant mechanisms between species [74]. Moreover, supplementation with L. fermentum MCC3216 exerted a more substantial reduction in lipid peroxidation and an increase in SOD and catalase protein levels, with comparable effects on GSH concentration in the pancreas of high fructose-fed diabetic Wistar rats as compared to the metformin-treated control group [101].
T2DM rats administered with L. plantarum SS18*-*5 increased SOD levels and activity and lowered MDA levels [118]. L. gasseri CKCC1913 supplementation yielded similar effects in T2DM mice [93]. L. plantarum SCS3 effectively recovered SOD, catalase, and GSH levels, providing antioxidant protection [119]. Similarly, the inactivated cellular content of L. plantarum SCS4 lowered ROS and MDA content and restored SOD, catalase, GPX, and GSH concentration to physiological levels [120]. In the kidneys of fructose-fed rats, L. plantarum and L. helveticus elevated SOD2 expression [121]. Supplementation of these Lactobacillus isolates improved glycaemic indices and enhanced insulin sensitivity in the diabetic models [93, 118–120].
Several studies suggested that Lactobacillus spp. exert their redox role through multiple mechanisms such as inherent generation of oxidative resistance products, regulation of nuclear factor erythroid 2 (NFE2)-related factor 2 (Nrf-2) signalling, modulation of NF-κB transcription factor activity, and uncoupling of endothelial nitric oxide synthase [122–125]. Numerous investigations revealed that the antioxidant properties of Lactobacillus protect peripheral organs from oxidative damage. For instance, L. paracasei L14 markedly increased beta-cell number, sustained the structural arrangement of pancreatic islets, and improved the hepatic histological morphology [112]. L. rhamnosus Hao9 and L. rhamnosus LRa05 improved hepatic injury by reducing oxidative stress-associated fibrosis and lowering vacuolar degeneration, respectively [74, 96]. Despite these promising results in the animal models, further clinical investigations are necessary to verify the therapeutic efficacy in humans.
As summarized in Table 3, these in vivo findings demonstrate that Lactobacillus spp. supplementation ameliorates oxidative stress-induced insulin resistance and hyperglycaemia by upregulating ROS-scavenging enzymes or nonenzymatic antioxidants as well as diminishing ROS production. These results also highlight the antioxidant defences such as SOD, catalase, and GSH. Hence, it shows that oxidative stress as a modifiable target via microbiome-based approaches. Nevertheless, the antioxidant effects differ across strains whereby strain selection remains essential for a precision microbiome modulation.Table 3. Summary of experimental studies evaluating the effects of Lactobacillus spp. on oxidative stress regulation in animal models of T2DM and metabolic syndrome. The table outlines strain-specific influences on enzymatic and non-enzymatic antioxidant systems and reactive oxygen species (ROS) generation*Lactobacillus speciesAnimal model (disease state)Dose, duration and route of administrationEffect on enzymatic antioxidantEffect on non-enzymatic antioxidantEffect on ROS generationReferenceL. casei *CCFM0412C57BL/6 J mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/day;12 weeks;oral administrationIncrease SODIncrease GSHReduce MDA and ROS[10]L. helveticusWistar rats (high fructose-induced metabolic syndrome)1 × 10⁹ CFU/100 g body weight;6 weeks;gastric gavageIncrease SOD2 - -[121]L. fermentum MCC3216Wistar rats (high fructose-induced T2DM)2.0 × 10⁸ CFU/mL;8 weeks;oralIncrease SOD and catalaseIncrease GSHReduce lipid peroxidation[101]*L. paracasei *NL41Sprague–Dawley rats (HFD/STZ-induced T2DM)1 × 10¹⁰ CFU/day;12 weeks;oral gavageIncrease catalase and SOD contentIncrease GPXReduce MDA concentration[113]*L. rhamnosus *LRa05Kunming mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/day;6 weeks;oral gavageIncrease SOD (not significant), decrease catalaseDecrease GSHReduce MDA[74]*L. plantarum SS18-*5Sprague Dawley rats (HGFD/STZ-induced T2DM)1 × 10¹⁰ CFU/day;6 weeks;oral gavageIncrease SOD -Reduce MDA concentration[118]L. plantarum SCS4 (inactivated cellular content) Kunming mice (STZ-induced T2DM)2 × 10⁹ CFU/dose;10 weeks;oral gavageIncrease SOD and catalaseIncrease GPX and GSHReduce ROS and MDA concentration[120]L. plantarum SCS3Kunming mice (STZ-induced T2DM)5 × 10⁹ CFU/day;10 weeks;intragastric administrationIncrease SOD and catalaseIncrease GSH -[119]*L. rhamnosus *Hao9C57BL/6J mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/day;11 weeks;oral gavageIncrease catalase and SOD - -[96]L. gasseri CKCC1913C57BL/6J mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/mL;6 weeks;oral gavageIncrease SOD -Reduce MDA concentration[93]*L. paracasei *(Malaysian water kefir grains)C57BL/6 mice (HFD/STZ-induced T2DM)Low dose (LD): 1 × 10⁶ CFU/mL; High dose (HD): 1 × 10¹⁰ CFU/mL;4 weeks;oral gavage- -Reduce MDA concentration[114]*L. paracasei *L14Sprague–Dawley rats (HFD/STZ-induced T2DM)1 × 10¹⁰ CFU/day;12 weeks;oral gavageIncrease catalase and SOD contentIncrease GPXReduce MDA concentration[112]*Abbreviations:*HFD high-fat diet, STZ streptozotocin, SOD superoxide dismutase, GSH glutathione, MDA malondialdehyde, ROS reactive oxygen species, GPX glutathione peroxidase, HGFD high glucose fat diet
Role of Lactobacillus spp. in Remodelling Diabetic Gut Microbiota
Lactobacillus spp. augments microbial diversity and richness, as indicated by the increase in Shannon index and Chao1 scores [74, 77, 126]. At the phylum level, the Firmicutes/Bacteroidetes (F/B) ratio is significantly lower upon the administration of L. acidophilus KLDS1.0901 and KLDS1.1003, L. casei CCFM419, L. casei LC89, L. plantarum LRCC5314, L. plantarum SHY130, L. rhamnose us LRa05, and L. paracasei IMC 502 in T2DM rodent models [74, 75, 77, 91, 126–128]. In contrast, the F/B ratio of T2DM patients typically exhibit a higher F/B ratio, with Firmicutes associated with insulin resistance and Bacteroidetes with improved glucose tolerance [50, 129–131]. While the exact mechanism has not been elucidated, the altered F/B ratio may lead to T2DM development by modulating inflammation.
Interestingly, Firmicutes perform saccharolytic fermentation, which may increase the gastroepithelial barrier permeability and induce inflammation, as indicated by its positive correlation with inflammatory markers TNF-α and IL-1β in obese and T2DM patients, respectively [132–134]. However, the Firmicutes phylum also comprises most of the butyrate-producers [135]. This paradox could be due to a reduction in butyrate-producing bacteria, which are replaced by other Firmicutes, leading to lower levels of the anti-inflammatory SCFA [136]. Therefore, restoring the F/B ratio and composition may normalize the SCFA levels and alleviate inflammation and thus improve insulin sensitivity.
Increase abundance of Parasutterella has been observed in the gut microbiota of T2DM patients [137, 138]. The administration of L. paracasei strains L14 and NL41 to T2DM rodent models reduced Parasutterella levels [112, 139]. Various studies have linked higher blood glucose levels and improved insulin sensitivity with increased Parasutterella in diabetic animals, supporting its pro-diabetic role [140–142]. As a high L-cysteine consumer, the high Parasutterella abundance reduces serum levels of this metabolite [137]. The inverse relationship between Parasutterella and L-cysteine levels may contribute to the pathogenesis of T2DM. L-cysteine supplementation improves glucose utilization by promoting the secretion of insulin-sensitizer adiponectin and reduces oxidative stress by increasing GSH levels and lowering ROS [143]. Therefore, by reducing Parasutterella abundance, the L. paracasei strains increase serum L-cysteine levels, which decreases oxidative damage to pancreatic beta cells and alleviates insulin resistance [137].
Lactobacillus species play important roles in managing diabetes by enhancing glucose uptake, reducing inflammation, lowering oxidative stress as well as supporting gut microbiota balance as shown in Fig. 2. Specific strains such as L. paracasei L14 and NL41 increased the abundance of the Lachnospiraceae NK4A136 group [112, 139]. L. rhamnosus LRa05 increased the abundance of Akkermansia [74]. Microbiota analyses of T2DM subjects and animal models have shown reduced abundance in both genera [142, 144, 145]. Lachnospiraceae NK4A136 group and Akkermansia are negatively associated with fasting blood glucose and HbA1c (glycated haemoglobin), respectively [142, 146]. The established correlation suggests that restoration of the genera abundance may mitigate the pathological state of T2DM.Fig. 2. Antidiabetic properties of lactobacillus spp. which play important roles in managing diabetes by enhancing glucose uptake, reducing inflammation, supporting gut microbiota balance and lowering oxidative stress. (Created with BioRender.com)
Intervention with metformin, the gold standard drug for T2DM, also induced similar effects by increasing their abundance in high-fat diet mice, further supporting their beneficial roles [147]. Lachnospiraceae NK4A136 group and Akkermansia are butyrate-producers [148, 149]. Butyrate promotes intestinal barrier integrity by increasing the expression of claudins, the main constituents of tight junctions [150]. Additionally, as mucin-degrading bacteria, Akkermansia increases the number of goblet cells, stimulating mucus synthesis [151]. Hence, an enriched abundance of Akkermansia and Lachnospiraceae NK4A136 group reduces intestinal permeability, prevents lipopolysaccharide entry to the vascular system, and reduces inflammation, ultimately alleviating glucose intolerance.
Table 4 summarizes the microbiota changes induced by various Lactobacillus spp. and the corresponding roles. Many studies reported that increased SCFA-producing taxa, improved in gut barrier function and reduced inflammatory leakage. These microbiota shifts are associated with lower inflammation and improve insulin sensitivity. While the consensus agrees that Lactobacillus spp. alter the microbiome composition to improve glycaemic control, the magnitude of the microbial community shifting may vary based on the specific strain and dose of probiotic administered. These strain-specific and ecosystem-dependent microbiota shifts supports the importance of precision microbiome modulation, in which targeted Lactobacillus interventions are aligned with host microbiome structure and functional capacity.Table 4. Effects of Lactobacillus spp. on gut microbiota composition and functional profiles in animal models. This table summarizes strain-dependent alterations in key microbial taxa and community structure, as well as the associated metabolic and immunomodulatory roles, demonstrating how Lactobacillus contributes to microbiota remodeling as a strategy for precision intervention in T2DMLactobacillus spp.Animal model (disease state)Dose, duration and route of administrationChanges in microbiotaRolesReferencesL. casei CCFM419C57BL/6J mice (HFD/STZ-induced T2DM)1 × 10⁹CFU/mL;12 weeks;intragastricLower Firmicutes/Bacteroidetes (F/B) ratio; increased total SCFAsModulates gut flora–SCFA–inflammation/GLP-1 axis, reduces inflammatory cytokines (TNF-α, IL-6) and improves insulin resistance[91]L. acidophilus KLDS1.1003 and KLDS1.0901C57BL/6J mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/day;6 weeks;oral gavageReduced F/B ratio; enrichment of SCFA-producing taxaRebalances gut microbiota, normalizes SCFA levels, improves glucose and lipid metabolism[77]L. rhamnosus LRa05Kunming mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/day;6 weeks;oral gavageIncreased SCFA-producing genera (Alloprevotella, Bacteroides); decreased proinflammatory genera (Odoribacter, Mucispirillum).Ameliorates hyperglycaemia and insulin resistance by modulating glucagon-mediated signalling, reducing hepatic oxidative stress and LPS-related inflammation, and partially reshaping gut microbiota toward more SCFA-producing taxa[74]L. casei LC89HFD/STZ-induced T2DM mice1 × 10⁹ CFU/day;6 weeks;oral gavageReduced F/B ratio; modulation of gut community structure towards SCFA-producing generaImproves hepatic glucagon response and insulin sensitivity via microbiota remodelling[75]Heat-killed* L. plantarum LRCC5314 (HK-LRCC5314*C57BL/6 mice (cold stress and HFD-induced T2DM)1 × 10⁸ CFU/day;12 weeks;oral administrationAltered gut community structure; enrichment of beneficial and butyrate-producing genera (Barnesiella, Alistipes, Akkermansia) and reduction of potentially pro-inflammatory genera (Ruminococcus, Dorea, Clostridium).Relieves stress-related hyperglycaemia and insulin resistance and lowers inflammatory markers via gut microbiome modulation[126]L. plantarum SHY130C57BL/6J mice (HFD/STZ-induced T2DM)10¹⁰ CFU/kg body weight per day;10 weeks;oral gavageReadjusts intestinal flora structure; enriches SCFA-producing genera (Faecalibaculum, Odoribacter, Alistipes); increases colonic SCFA levels.Supports enteroinsular axis regulation and glycaemic control by increasing SCFA-producing bacteria, upregulating colonic SCFA receptors GPR41/GPR43, and protecting islet α/β-cell balance[127]L. paracasei IMC 502C57BL/6J mice (HFD/STZ-induced T2DM)1 × 10⁹ CFU/day;15 weeks;oral administrationReduced F/B ratio; enrichment of SCFA-producing generaMediates the gut microbiota–SCFA–hormone/inflammation pathway to ameliorate T2DM[12]L. paracasei NL41Sprague–Dawley rats (HFD/STZ-induced T2DM)1 × 10¹⁰ CFU/day;12 weeks;oral gavageRegulates intestinal microbiota and inflammatory response (unpublished data)Decreases insulin resistance and oxidative stress, protects β-cells, and improves glycaemic and lipid metabolism[113]*L. paracasei L14L. paracasei *L141 × 10¹⁰ CFU/day;12 weeks;oral gavageSprague–Dawley rats (HFD/STZ-induced T2DM)Decreased Parasutterella; increased *Lachnospiraceae *NK4A136 group[112]*Abbreviations:*HFD high-fat diet, STZ streptozotocin, T2DM type 2 diabetes mellitus, SCFA short-chain fatty acid
Precision Nutrition Development Using Lactobacillus spp. for T2DMz
Leveraging advancements in precision medicine, which focuses on personalized healthcare based on individual variability in genes, environment and lifestyle, has driven the recent precision and personalized nutrition strategies [152, 153]. Numerous studies reported that different probiotic strains of Lactobacillus may play a pivotal role in modulating host glycaemic responses, oxidative stress pathways and inflammatory states.
Preclinical Evidence
At the preclinical level, multiple studies demonstrate that fermented foods containing Lactobacillus spp. exert antioxidant and glucose-regulating effects in animal models. A study has shown that T2DM mice treated with defatted rice bran fermented by L. fermentum MF423 exhibited upregulated levels of SOD, total antioxidant capacity (T-AOC) and GPX and diminution of MDA levels [154]. These effects were non-significant upon treatment with unfermented rice bran, demonstrating that Lactobacillus fermentation enhanced the antioxidant capacity to confer hepatic protection and improve glucose and lipid metabolism [154]. Similarly, fermentation of bitter melon juice with L. fermentum LLB3 exhibited more potent antioxidant capacity in T2DM rats, with a significant increase in SOD levels compared to the nonfermented juice [155]. Comparable findings were observed in T2DM rats treated with L. plantarum NCU116-fermented carrot juice, resulted in a greater increase in CAT, GPX, and SOD activities and T-AOC levels than its non-fermented form [156].
Meanwhile, various Lactobacillus spp. were found to exhibit regulatory effects on glucose metabolism pathways. The hypoglycaemic effects of L. plantarum-fermented rice germ extracts in C57BL/6KsJ-db/+ mice were attributed to the modulation of hepatic glucose metabolism [10]. The fermented extract upregulated the expression of glucokinase to stimulate glucose storage and utilization and downregulated the expression levels of G6Pase and PEPCK to reduce gluconeogenesis [10]. Based on* in vitro* assays, fermentation with L. plantarum FNCC 0027 elevated the inhibitory activities of α-amylase, α-glucosidase, and amyloglucosidase in Jamaican cherry juice, which lowers glucose absorption [157]. Similarly, yoghurt fermented with L. plantarum KU985438 and L. rhamnosus KU985439 significantly reduced α-amylase concentrations in STZ-induced diabetic rats [158]. Black goji berry juice fermented with L. rhamnosus GG exhibited more potent DPP-IV inhibitory activity in vitro [159]. DPP-IV inhibition prevents the degradation of GLP-1 and increases its endogenous levels, resulting in elevated insulin secretion and lower glucose production to enhance glycaemic control [159]. However, the therapeutic superiority of fermented matrices over stand-alone probiotic supplementation remains inconsistent. A study showed that L. casei Q14 fermented yoghurt improved glucose tolerance and insulin levels, downregulated the expression of gluconeogenesis-associated enzymes, and re-established a protective microbiota [160]. The antidiabetic effects of the fermented yoghurt, while more effective than plain yoghurt, were comparable to that of the probiotic-only group, suggesting the strain itself was the primary driver of bioactivity [160]. L. gasseri SBT2055-fermented skim milk powder increased the expression of insulin genes and transcription factor Pdx1, which promoted glycogen levels in diabetic Goto-Kakizaki rats [161]. The fermented milk powder also alleviated inflammation by suppressing serum levels of pro-inflammatory cytokines TNF-α, macrophage inflammatory protein-1α (MIP-1α), and IL-18 and pancreatic levels of granulocyte colony-stimulating factor (G-CSF) [161]. Nevertheless, these findings suggest that the food matrix may not always enhance the intrinsic metabolic benefits of the Lactobacillus strain. Furthermore, some studies have not shown clear advantages of fermented products over probiotics alone, raising questions about the specific role of fermentation in mediating these benefits.
However, its effects on the diabetic indices, such as postprandial glucose response and plasma insulin levels, were similar to the unfermented equivalent [161]. In diabetic Wistar rats, L. plantarum NCU116-fermented Momordica charantia juice ameliorated oxidative stress and induced significant hypoglycaemic, hypoinsulinemic, and hypolipidemic effects as compared to the untreated controls [162]. While the fermented juice exhibited more potent antidiabetic effects than the nonfermented counterpart, the differences were also non-significant [162].
Clinical Evidence
Clinical evidence also supports the potential role of Lactobacillus spp. in metabolic management. For instance, current clinical evidences support the usage of Lactobacillus strains in improving glycaemic control. A recent randomized clinical trial reported that probiotic Lactobacillus paracasei HII01 improved glycaemic control in T2DM patients throughout the 12 weeks of intervention with no serious adverse effects or symptoms among the participants [1]. This is in accordance with another study by Khalili and colleagues who demonstrated that Lactobacillus casei could improve glycemic response in patients with T2DM [29]. Despite the different strains of Lactobacillus employed in these studies, the findings revealed the potential of utilising Lactobacillus spp as an adjunct treatment for T2DM. Furthermore, the effectiveness of Lactobacillus spp. in managing T2DM extends to augmenting the properties of functional food, which is associated with enhanced antioxidant effects and enzymatic expression and activity. However, the relative contribution of fermentation versus strain-specific bioactivity remains unclear.
From a practical consumer health perspective, current findings suggest that selected Lactobacillus-containing foods or supplements may be utilised as beneficial adjunct therapies to support glycaemic control and attenuate oxidative stress in T2DM. However, it is worth noting that not all the fermented products or probiotic formulations labelled with Lactobacillus necessarily exert clinically relevant antidiabetic effects. Therefore, these products should be considered as adjuncts, not substitutes, for standard dietary and pharmacological T2DM management. Their use is best justified when supported by robust clinical evidence for the specific strain, dose and formulation along with ongoing well-controlled studies which are essential to validate and optimise these recommendations.
Overall, advancing precision nutrition with Lactobacillus spp. for T2DM requires further exploration on strain–substrate interactions, effectiveness of fermentation and patient-specific responses. In particular, the fermentation-induced bioactivity-enhancing effect may be associated with discrete strains or food types and vary according to the inoculation concentration, treatment dose and treatment duration. The fermentation protocol should also be optimized for each strain as changes in incubation conditions, including time, pH, temperature, and salt concentration, can alter SCFA production [163, 164]. Moreover, prospective studies should investigate the individual and combined effects of the food and specific Lactobacillus strain to accurately determine the beneficial effects of Lactobacillus fermentation.
Additionally, the antidiabetic effects of fermented food products should be explicitly distinguished between their inherent antihyperglycemic properties and the fermentation-induced enhancements by comparing the metabolite content pre- and post-fermentation. Furthermore, to ensure result consistency, a consensus should be drawn to clearly define the primary endpoint as a measure of therapeutic effectiveness. Ultimately, application of strain-specific strategy which integrates both host and microbial factors is crucial to harness the potential of Lactobacillus-based precision nutrition in T2DM management as summarised in Fig. 3.Fig. 3. The schematic illustrates how host factors (e.g., genetics and baseline metabolic phenotype) and lifestyle factors (diet, physical activity, smoking, sleep and medication use) interact with gut microbiome features to influence Lactobacillus spp. modulation of* Lactobacillus* through targeted functional foods is integrated into a precision-nutrition workflow aimed at optimizing glycemic control and improving T2DM management (Created with BioRender.com)
Challenges and Future Perspectives
Safety and the Risk of Antimicrobial Resistance
One of the primary challenges in the application of Lactobacillus-based probiotics for therapeutic purposes lies in ensuring bacterial viability and stability during gastrointestinal transit. Probiotic treatment involves the consumption of large quantities of bacteria: a minimum dose of 106 CFU bacteria is needed to exert a beneficial effect, but higher administration doses of at least 109 CFU are usually required to account for the ablated bacteria viability due to harsh gastric environments [91]. Several Lactobacillus strains resist low pH and bile acids [165, 166].
However, not all strains demonstrate such tolerance and exhibit impaired stability and survivability in internal gastric conditions, thus reducing their therapeutic efficacy. Various methods can account for the variability in strain tolerance, including probiotic encapsulation and bioengineering. Lactobacillus spp. encapsulation with alginate, pectinate, and spray drying retains high viability [167–170]. A recent study reported that advanced encapsulation approaches using alginate–basil seed mucilage–prebiotic microencapsulation showed markedly improve probiotic Leuconostoc mesenteroides ABRIINW.N18 viability in yoghurt during storage and simulated gastrointestinal passage [171]. Hence, these technologies could be adapted for Lactobacillus strains used in T2DM interventions to enhance their delivery to distal regions of the gut.
In addition, safety and regulatory challenges pose significant obstacles to the broader clinical implementation of Lactobacillus spp. in T2DM management. Although their therapeutic benefits are indisputable, several publications have raised safety concerns about probiotic consumption. Long-term supplementation of a multi-strain Lactobacillus mixture in healthy rats induced systemic pro-inflammatory responses, elevated cardiovascular risk, and enlarged colonic lymphoid aggregates [172]. Lactobacillus probiotic supplementation has also been associated with sepsis, Lactobacillus endocarditis, and vertebral osteomyelitis [173–175].
Due to poor regulation, the vague probiotics formulations marketed are especially detrimental in managing adverse health conditions such as inflammatory bowel diseases or immunosuppressive disorders [176]. Therefore, it is vital to verify the adverse effect profile of various preparations and ensure that the product adheres to the stringent quality requirements and is contamination-free [177]. An innovative method is to extract the intracellular content of Lactobacillus probiotics and subsequently inactivate it to prevent septic infection while still retaining efficacy in T2DM amelioration [70].
Nevertheless, navigating the clinical translation of Lactobacillus spp. requires heightened regulatory scrutiny following recent updates from the Food and Drug Administration (FDA) and The European Food Safety Authority (EFSA), which increasingly differentiate therapeutic live biotherapeutic products (LBPs) from dietary supplements [178, 179]. Furthermore, a recent large‑scale whole‑genome sequencing analysis of 579 isolates from 12 commonly used probiotic species identified mobile antibiotic‑resistance genes (ARGs) in commercial Lactobacillus strains, highlighting potential safety risks and the need for careful safety assessments [180].
Furthermore, probiotics pose a risk as potential reservoirs ARGs. Unlike intrinsic ARGs, the acquired ARGs may be disseminated via horizontal gene transfer in the environment, conferring the resistant trait to potentially pathogenic microbes. Several studies have affirmed the presence of ARG among probiotic Lactobacillus strains. The blaTEM resistance gene, encoding extended-spectrum beta-lactamases against β-lactams antibiotics, has been detected in various probiotics-derived Lactobacillus isolates [181].
Similarly, at least one ARG has been detected in L. plantarum, Lactobacillus delbrueckii, and L. helveticus obtained from probiotics and food products [182]. While the ARG detection frequency is relatively lower than other bacterial species, ARGs are potentially transferable since they are plasmid-associated, with those in L. plantarum being exceptionally diverse [182]. Therefore, the commercialization of probiotics may invoke selective pressure and favour highly resistant Lactobacillus species, contributing to the emergence and spread of antimicrobial resistance.
Precision Probiotics: Multi-omics Profiling and Personalized Interventions
Despite the promising antidiabetic potential of Lactobacillus spp., the strain-specific mechanisms by which Lactobacillus spp. modulate glycaemic control remain poorly understood [15]. This impedes the development of precise and targeted probiotic interventions which require further research to comprehend the relationship between Lactobacillus spp. and individuals with diabetes. Consequently, it is crucial to identify selection of strains based on their distinct mechanisms of action or biomarkers for response prediction. Thus, integrating comprehensive clinical and multi-omics profiling, including personalized microbiome profiling could identify T2DM subgroups with specific Lactobacillus targets and enable personalized probiotic interventions [183, 184]. The integration of diverse high-throughput multi-omics datasets enabling pharmacomicrobiomics encompasses of different modules including amplicon sequencing, metagenomics, metatranscriptomics, metaproteomics and metabolomics/metabonomics [50]. These modules enable researchers to investigate a wide range of gene variation and modulation datasets obtained from multi-omics sequencing.
With the recent advancement in artificial intelligence, machine learning models and bioinformatics tools can be utlised to individual responses to probiotics and support personalized therapeutic decision-making. These technologies facilitate the development of tailored microbiome interventions by integrating host-specific data such as genetic background, existing gut microbiota composition and metabolic status for the T2DM patients. Moving away from traditional “one-size-fits-all” treatment to precision-based medicine, the current technology leverages the big data from patients’ electronic health records. This includes genomic information, biomarkers, laboratory and radiological investigations data [11, 185, 186].
Hence, precision medicine strategies which assess the patient's genetic profile, lifestyle habits and environmental factors are vital in guiding individualized treatment decisions and predicting therapeutic outcomes. Nevertheless, longitudinal clinical trials are critical to establish causal relationships between specific* Lactobacillus *strains and glycaemic control in T2DM patients [187]. Such trials are essential to translate the promising potential of targeted probiotic therapies into clinical practice by identifying strain-specific indications, optimal dosages, delivery routes as well as safety monitoring under a precision medicine framework. Figure 4 illustrates the shift from traditional probiotic approaches to a precision medicine framework.Fig. 4. Integrated workflow for developing precision Lactobacillus probiotic interventions in T2DM. The process begins with the acquisition of "big data," integrating comprehensive host clinical profiles (genetics, lifestyle) with diverse high-throughput multi-omics datasets. Artificial intelligence (AI), machine learning (ML) and bioinformatics tools process this integrated data to overcome current knowledge gaps regarding strain-specific mechanisms. This analytical step identifies distinct T2DM subgroups and matches them with specific Lactobacillus targets based on predicted response biomarkers. The final outcome is the implementation of tailored microbiome interventions, guiding individualized therapeutic decision-making away from a "one-size-fits-all" model. (Created with BioRender.com)
Alternative Microbiome-targeted Strategies: Paraprobiotics, Postbiotics and Fecal Microbiota Transplantation
While the direct administration of live Lactobacillus spp. represents a primary strategy for precision microbiome modulation in T2DM management, alternative approaches are rapidly gaining traction to mitigate infection risks while retaining beneficial effects. The current therapeutic landscape is shifting to include non-viable microbial components and metabolites. This has led to the emergence of paraprobiotics (inactivated microbial cells or cell fractions) and postbiotics (bioactive microbial metabolites or cell-free components), representing the next generation of microbiome-based interventions, which offer advantages such as greater stability, safety and a more focused mechanism of action.
Paraprobiotics also referred to as inactivated or heat-killed probiotics, represent non-viable microbial cells or fractions that confer a health benefit to the host [188]. Findings revealed that heat-killed strains of Lactobacillus and Bifidobacterium, exert their anti-diabetic and metabolic effects primarily through direct immune modulation and gut barrier fortification via their structural component [189]. Various studies showed that paraprobiotics are able to modulate inflammatory and oxidative pathways which helps in improving glucose homeostasis [189]. A current study reported that the purified extracellular polysaccharide (EPS) derived from the novel strain Lactobacillus acidophilus YL01 possess significant insulin-sensitizing and anti-obesity properties in high-fat mice via modulating intestinal specific bacterial groups and AMPK/ACC signaling pathway [190]. Overall, non-viable approaches offer therapeutic benefits while reducing the risks associated with live bacteria, providing a more favorable safety profile.
Meanwhile, postbiotics are defined as “a preparation of inanimate microorganisms and/or their components that confers a health benefit on the host”, according to the International Scientific Association for Probiotics and Prebiotics (ISAPP) in year 2021 [191]. It has shown postbiotics has pharmacological advantages over live probiotics, including superior safety profiles for immunocompromised individuals, enhanced thermal and storage stability as well as precise dosing [192]. A current randomized controlled trial (RCT) has demonstrated that supplementation with a specific Lactobacillus rhamnosus-derived postbiotic preparation for three months led to a significant reduction HbA1c and fasting plasma glucose in T2DM patients, primarily attributed to an improvement in beta cells functional activity. Furthermore, postbiotics are being explored as emerging treatment against diabetic microvascular complications like diabetic retinopathy.
Next, is the fecal microbiota transplantation (FMT), which may consider as a comprehensive microbiome-targeted therapy by transferring stool from a healthy donor into a T2DM recipient. Recent systematic review and meta-analysis evaluated the efficacy of FMT in T2DM patients found that FMT primarily impacts indices like HbA1c, fasting plasma glucose and insulin resistance [193]. Study reported that FMT from metabolically healthy donors can transiently improve peripheral insulin sensitivity, accompanied by shifts in gut microbiota composition and SCFA profiles [194, 195]. While multiple RCTs and animal studies show significant metabolic benefits, individual responses vary and data in established T2DM are limited. Therefore, it is crucial to conduct strict donor screening and standardized protocols for the safe use of FMT in the future.
Engineered Probiotics and Synthetic Biology Approaches
Next, the synthetic biology and next-generation probiotic technologies enable the engineering of Lactobacillus spp. with tailored functionalities for glycaemic control in T2DM for enhanced therapeutic applications. For example, CRISPR-based engineering of tailored probiotics allows precise genome editing and transcriptional regulation in these bacteria, aiming to enhance stability and biodelivery for targeted health interventions [196, 197]. For instance, the next-generation probiotic L. plantarum-pMG36e-GLP-1, expressing glucagon-like peptide-1 (GLP-1) demonstrated improvements in glycemic control, pancreatic function and liver metabolism in diabetic mouse models [198].
In a recent study, a genetically engineered strain of Escherichia coli Nissle 1917 (EcN)-GLP-1 demonstrated protective effects on the islet β-cells, reduced key inflammatory markers including TLR-4, p-NF-κB/NF-κB, and Bax/Bcl-2 besides helped restore balanced gut microbial diversity in diabetic mice models [199]. Overall, advancement of engineered bacterial therapies to produce therapeutic molecules or modulate the gut microbiota brings healthcare closer to personalized medicine by reducing patient outcomes and minimizing side effects.
Conclusion
This review summarized the evidence from various studies, suggesting the potential therapeutic mechanisms of Lactobacillus spp. for T2DM treatment. These include modulation of glucose metabolism and transport, anti-inflammatory effects, anti-oxidative effects, and restructuring of the gut microbiota composition. Besides that, they may also exert its antidiabetic actions by enhancing functional foods, but the extent of the effects may vary. Despite promising findings, clinical translation is hindered by multiple challenges, including strain-specific survivability in the gastrointestinal tract, safety concerns and risk of antibiotic resistance gene transfer. Nevertheless, from the perspective of consumer-health, selected Lactobacillus-containing foods or supplements may offer supportive benefits but not replacement for established pharmacological therapy.
The future of Lactobacillus-based interventions in T2DM lies in adopting precision medicine frameworks. The integration of precision medicine, supported by multi-omics technologies and artificial intelligence enable personalized microbiome-based therapies. Personalized microbiome profiling and electronic health data can be leveraged to tailor probiotic strain selection, dosage and delivery methods, moving beyond the traditional "one-size-fits-all" model. Furthermore, advances in synthetic biology and next-generation probiotic engineering offer exciting opportunities to enhance the functionality of Lactobacillus strains. In a nutshell, a multidisciplinary approach with the combination of microbiology, genomics, artificial intelligence and clinical sciences are essential to unlock the full potential of targeted Lactobacillus-based therapies for personalized T2DM treatment.
Key References
- Zhang C, Jiang J, Wang C, Li S, Yu L, Tian F, Zhao J, Zhang H, Chen W, Zhai Q. Meta-analysis of randomized controlled trials of the effects of probiotics on type 2 diabetes in adults. Clin Nutr. 2022 Feb;41(2):365–373. 10.1016/j.clnu.2021.11.037. Epub 2021 Dec 10. PMID: 34999331.
- ○ This large-scale meta-analysis consolidates evidence from randomized controlled trials on probiotics in T2DM, including Lactobacillus strains, demonstrating their glycemic benefits.
- Zhao Q, Chen Y, Huang W, Zhou H, Zhang W. Drug-microbiota interactions: an emerging priority for precision medicine. Signal Transduct Target Ther. 2023 Oct 9;8(1):386. 10.1038/s41392-023-01619-w. PMID: 37806986; PMCID: PMC10560686.
- ○ This article provides a cutting-edge overview of how gut microbiota influences drug response, directly supporting the rationale for pharmacomicrobiomics and potential of tailoring* Lactobacillus*-based interventions for T2DM treatment.
- Jiang S, Liu A, Ma W, Liu X, Luo P, Zhan M, Zhou X, Chen L, Zhang J. Lactobacillus gasseri CKCC1913 mediated modulation of the gut-liver axis alleviated insulin resistance and liver damage induced by type 2 diabetes. Food Funct. 2023 Sep 19;14(18):8504-8520. 10.1039/d3fo01701j. PMID: 37655696
- ○ This article reported the strain-specific effects of *Lactobacillus *on the gut–liver axis, linking probiotic modulation to improved insulin resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lê K-A et al. Alterations in fecal Lactobacillus and Bifidobacterium species in type 2 diabetic patients in Southern China population, (in English). Front Physiol Original Res 2013;3 2013-January-31. 10.3389/fphys.2012.00496.10.3389/fphys.2012.00496 PMC 356036223386831 · doi ↗ · pubmed ↗
- 2Bhute SS, Suryavanshi MV, Joshi SM, Yajnik CS, Shouche YS, Ghaskadbi SS. Gut Microbial Diversity Assessment of Indian Type-2-Diabetics Reveals Alterations in Eubacteria, Archaea, and Eukaryotes, (in English). Frontiers in Microbiology, Original Research 2017;8 2017-February-14. 10.3389/fmicb.2017.00214.10.3389/fmicb.2017.00214 PMC 530621128261173 · doi ↗ · pubmed ↗
- 3Nam Y et al. Heat-killed Lactiplantibacillus plantarum LRCC 5314 mitigates the effects of stress-related type 2 diabetes in mice via gut microbiome modulation. 2022;32(3):324-332. 10.4014/jmb.2111.11008.10.4014/jmb.2111.11008 PMC 962885234949748 · doi ↗ · pubmed ↗
- 4Bahar-Tokman H et al. Firmicutes/Bacteroidetes ratio in the gut microbiota and IL-1β, IL-6, IL-8, TLR 2, TLR 4, TLR 5 gene expressions in type 2 diabetes. Clin Lab. 2022;68(9). 10.7754/Clin.Lab.2022.211244.10.7754/Clin.Lab.2022.21124436125161 · doi ↗ · pubmed ↗
- 5Rahman A, Alqaisi S, Nath J. A Case of Lactobacillus casei Endocarditis Associated With Probiotic Intake in an Immunocompromised Patient. Cureus. 2023;15(4).10.7759/cureus.38049 PMC 1020784337228522 · doi ↗ · pubmed ↗
- 6Food US, Drug A. Early Clinical Trials With Live Biotherapeutic Products: Chemistry, Manufacturing, and Control Information; Guidance for Industry, U.S. Food and Drug Administration, 2016. [Online]. Available: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/early-clinical-trials-live-biotherapeutic-products-chemistry-manufacturing-and-control-information
- 7Tóth AG, Judge MF, Nagy S, Papp M, Solymosi N. A survey on antimicrobial resistance genes of frequently used probiotic bacteria, 1901 to 2022, (in eng). Euro Surveill. 2023;28(14). 10.2807/1560-7917.Es.2023.28.14.220027210.2807/1560-7917.ES.2023.28.14.2200272 PMC 1028346137022212 · doi ↗ · pubmed ↗
- 8Dhieb D et al. Harnessing pharmacomultiomics for precision medicine in diabetes: a comprehensive review, (in eng). Biomedicines. 2025;13(2). 10.3390/biomedicines 13020447.10.3390/biomedicines 13020447 PMC 1185302140002860 · doi ↗ · pubmed ↗