Modulation of tocotrienol’s bone effects by osteocytes: a perspective
Nurul Nabihah Zahanordin, Pei Yuen Ng, Kok-Yong Chin

TL;DR
This paper explores how tocotrienols, a form of vitamin E, may protect bones by influencing osteocytes, which are key cells in bone health.
Contribution
The paper highlights the novel perspective that tocotrienols may act through osteocytes to improve bone quality.
Findings
Tocotrienols reduce oxidative stress and modulate osteocyte-secreted factors like sclerostin and DMP1.
TTs show protective effects on osteocytes in animal models of metabolic syndrome and ovariectomy.
Current research is limited by model specificity and the need for advanced techniques to study TT mechanisms.
Abstract
Osteocytes function as central regulators of skeletal health by acting as mechanosensors that control bone remodelling mediated by osteoblasts and osteoclasts. Disrupted osteocyte function, often driven by oxidative stress and linked to ageing and osteoporosis, contributes to pathological bone remodelling. Tocotrienols (TTs), a family of vitamin E, are intensively investigated for their bone-protective effects, with mechanisms that involve reducing intracellular reactive oxygen species, enhancing antioxidant defences, and modulating signalling pathways of bone remodelling. Preliminary studies suggest that TTs exert protective and anabolic effects by influencing osteocytes, including shielding them from oxidative damage. In vivo models using ovariectomised or metabolic syndrome rats demonstrated that TT supplementation modulated key osteocyte-secreted factors, including sclerostin,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
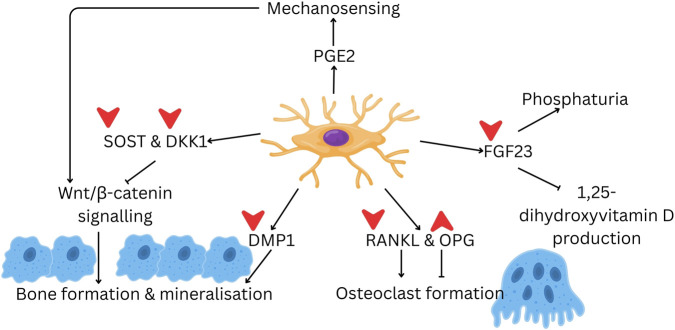 FIGURE 1
FIGURE 1| Study | Model | Treatment | Changes in osteocyte parameters (vs. osteoporosis control) | Remarks | ||
|---|---|---|---|---|---|---|
| Increased | Decreased | Unchanged | ||||
|
| MLO-Y4 cells exposed to t-BHP at 250 µM or 125 µM for 3 h | Delta-TT (1.25–20 μg/mL for 2 h) (pre-treatment) | Cell viability (1.25–10 μg/mL) | Apoptosis (5 μg/mL) | - | The redox signalling parameters were tested in osteoblasts only |
|
| Wistar rats fed with high-fat high-carbohydrate (HFHC) diet | Palm or annatto TT (60 or 100 mg/kg) (p.o.) | - | Skeletal protein expression of SOST, DKK-1, FGF23 and RANKL | Skeletal protein expression of OPG | Treatment was started 8 weeks after HFHC diet was started |
|
| Wistar rats fed with high-fat high-carbohydrate (HFHC) diet | Palm or annatto TT (60 or 100 mg/kg) (p.o.) | - | Ct empty lacunae number (AnTT at 100 mg/kg) | Tb and Ct osteocyte number | Treatment was started 8 weeks after HFHC diet was started |
|
| Ovariectomised female SD rats | Formulated or self-emulsified annatto TT (100 mg/kg) for 2 months (p.o.) | - | Skeletal SOST protein level | Tb osteocyte number | Treatment was started 2 months after ovariectomy |
|
| Ovariectomised female SD rats | Formulated (100 mg/kg) or self-emulsified palm TT (50 mg/kg) for 10 weeks (p.o.) | Tb osteocyte number | Skeletal | Tb and Ct empty lacunae number | n/a |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntioxidant Activity and Oxidative Stress · Vitamin K Research Studies · Bone Metabolism and Diseases
Introduction
1
Osteoporosis is a bone disease characterised by degenerative changes in bone quantity (bone mass) and quality (microstructure) associated with ageing, leading to a decline in bone strength and an increased risk of fractures (Chin et al., 2022). It is an increasingly important healthcare issue in the context of global societal ageing, given the healthcare burden associated with fragility fractures (GBD, 2019 Fracture collaborators, 2021). Both sexes are affected by osteoporosis; postmenopausal women are disproportionately affected by the disease (Liang et al., 2025), but men suffer from a higher mortality post-fracture (Larsen et al., 2025). Various pharmacology interventions have been designed to improve bone mass and reduce the risk of fractures, each with its advantages and side effects (Larsen et al., 2025).
Antiosteoporosis drugs primarily target the bone remodelling processes mediated by osteoblasts (bone-forming cells) and osteoclasts (bone-resorbing cells) (Elahmer et al., 2024). Recently, the most abundant bone cells, osteocytes, have also been identified as an avenue for intervention (Pathak et al., 2020). Osteocytes play a significant role in maintaining skeletal health. They are terminally differentiated from osteoblasts, which become embedded within the mineralised extracellular matrix of bone and assume regulatory functions. They have an extensive lacuno-canalicular network that acts as mechanosensor (Robling and Bonewald, 2020). They sense mechanical stress and regulate the function of osteoblasts and osteoclasts to maintain bone remodelling and structural stability (Yan et al., 2020).
Osteocytes secrete regulatory molecules, such as receptor activator of nuclear factor kappa-B ligand (RANKL), osteoprotegerin (OPG), dentin matrix protein 1 (DMP1), sclerostin (SOST), and fibroblast growth factor-23 (FGF23) (Kitaura et al., 2020). RANKL is involved in osteoclastogenesis, while OPG counterbalances this effect by preventing excessive bone resorption (Divieti Pajevic and Krause, 2019). SOST maintains bone formation within physiological limits by suppressing Wnt/β-catenin signalling in osteoblasts, limiting their survival and activities (Chen et al., 2025). Additionally, FGF23 regulates phosphate metabolism, linking osteocyte activity to systemic mineral homeostasis (Kurpas et al., 2021). The current antibody-based therapies for osteoporosis target SOST (e.g., romosozumab) and RANKL (e.g., denosumab) to achieve their therapeutic potential (Pang et al., 2020; Wu et al., 2023).
Tocotrienols (TTs) are a natural product that has been investigated intensively for its bone-protective effects through multiple mechanisms, potentially involving osteocytes (Chin, 2024a; Chin, 2024b). TTs are a family of vitamin E with double bonds on the carbon tail. They are found mainly in palm oil, annatto bean and rice bran, along with other sources, including grains and grapes (Pang and Chin, 2019). TTs contribute to the reduction of intracellular reactive oxygen species (ROS), enhance cellular antioxidant defences, and regulate the RANKL/OPG balance. Furthermore, they are involved in protective signalling mechanisms, notably the phosphoinositide 3-kinase/protein kinase B-nuclear factor erythroid 2-related factor 2 (NRF2) signalling pathway (Casati et al., 2020). TTs also play a significant role in suppressing the mevalonate pathway while also promoting the increased expression of osteoprotegerin (OPG) mRNA. This regulatory mechanism results in decreased production of RANKL. Consequently, this signalling cascade leads to diminished bone resorption, thereby enhancing protection against bone loss (Wong et al., 2019b; Ormsby et al., 2022).
Recent studies suggest that the therapeutic effects of TTs are mediated by osteocytes, but a structured review on this aspect has not been conducted. Thus, this review aims to summarise previous studies regarding the effects of TTs on osteocytes, focusing on their potential mechanisms of action, cellular targets, and effects on bone health. The limitations and prospects of this field are also put forward to guide future research.
Osteocytes
2
Osteocytes and bone remodelling
2.1
As stated earlier, osteocytes came from embedded osteoblasts within the lacuna-canalicular network. Osteocytes generate signals that are essential for controlling osteoclast formation and regulating phosphate metabolism, including proteins such as DMP1, SOST, and FGF23. Additionally, they serve as a significant source of RANKL (Wu et al., 2025). All these factors help osteocytes function as central regulators of bone remodelling (Figure 1).
The multifaceted role of osteocytes in regulating bone remodelling. Osteocytes secrete signalling molecules, including PGE2, DMP1, FGF23, SOST, DKK1, RANKL, and OPG, which regulate various processes of bone remodelling, such as mechanosensing, mineral metabolism, bone formation, and osteoclast formation. The red arrows indicate potential targets of tocotrienol; up arrows indicate upregulation, and down arrows indicate downregulation. Abbreviation: DKK1, dickkopf-related protein 1; DMP1, dentin matrix protein 1; FGF23, fibroblast growth factor 23; OPG, osteoprotegerin; PGE2, prostaglandin E2; RANKL, receptor activator of nuclear factor κB ligand; SOST, sclerostin. (Drawn using Canva).
The processes of mechanotransduction play a critical role in bone remodelling, which is regulated by osteocytes. Notably, the activation of Wnt/β-catenin signalling is partly mediated through Piezo one and integrin-linked pathways. This activation results in the suppression of SOST, thereby establishing a critical nexus between mechanical loading and the processes of bone formation and resorption (Wu et al., 2025). Furthermore, Wnt signalling in osteocytes increases the formation of osteoclasts by increasing RANKL expression through interactions with transforming growth factor-β (TGF-β) and Smad4 at the promoter region of RANKL (Ye et al., 2025).
Disruption of osteocyte function contributes to pathological remodelling. Processes such as apoptosis, ferroptosis or pyroptosis can disrupt the lacuna-canalicular network. This will lead to impairment of remodelling and repair capacity (Wu et al., 2025). Various osteocyte-targeted strategies show significant potential in restoring osteocyte function. These include anticytokine therapy, gene editing, stem cell interventions, and multi-omics approaches (Wu et al., 2025; Liu et al., 2025).
Pathophysiology: oxidative stress and inflammation in osteocytes
2.2
Oxidative stress is highly recognised as the central drive of skeletal deterioration. Elevated reactive oxygen species (ROS) will impair osteocyte homeostasis. Oxidative stress serves as a central mechanism in response to various insults, including glucocorticoids and homocysteine. These sources impaired osteocyte biology and activated maladaptive bone remodelling. These exposures upregulate the NOX family of NADPH oxidases, driving the cytokine expression and apoptosis in osteocyte-like cells (Notsu et al., 2019; Zhang et al., 2022). Mitochondrial dysfunction due to ageing or injury-related causes will also contribute to the upregulation of ROS, resulting from impaired mitophagy (Ardura et al., 2020).
The resulting reactive oxygen species activate a network of stress-responsive signalling pathways that reshape osteocyte survival, communication, and remodelling output. ROS stimulate the NLRP3 inflammasome, promoting caspase-1 activation, IL-1β maturation, and pyroptotic cell death (Kar et al., 2019). Activation of NF-κB and MAPK pathways leads to elevations in RANKL expression and the production of pro-resorptive cytokines (Marcucci et al., 2023). Oxidative stress also inhibits Wnt/β-catenin signalling by diverting β-catenin towards FOXO transcription factors, suppressing anabolic bone formation (Marcucci et al., 2023).
Tocotrienols
3
Tocotrienols: overview of skeletal protective mechanisms
3.1
Similar to the classic vitamin E, tocopherols (TPs), TTs can be categorised into four subtypes, such as alpha-, beta-, gamma-, and delta-homologues, differing by arrangement of the methyl group on the chromanol ring (Chin et al., 2013). TT and TF homologues exist in a mixture in natural sources (Aggarwal et al., 2010).
TTs exert both oxidative and non-oxidative actions on the skeletal system to achieve their anti-osteoporotic effects. TTs stimulate bone growth by facilitating the differentiation of osteoblasts and protecting these cells from oxidative stress (Abd Manan et al., 2012; Casati et al., 2020). It upregulates osteoblastogenic transcription factors, such as Runt-related transcription factor 2 (Runx2) and Osterix, thereby promoting the differentiation of osteoblasts (Wan Hasan et al., 2018). This action is potentially modulated by 3-hydroxy-3-methylglutaryl-coenzyme A reductase and GTPase activities (Wan Hasan et al., 2020). TTs also stimulate the Wnt signalling pathway to achieve pro-osteoblastogenesis effects (Xu et al., 2022). The anti-osteoclastogenic effects of TTs are mediated by the RANKL/OPG pathway and its anti-inflammatory effects. TT supplementation has been shown to modify the RANKL/OPG ratio and lower inflammation in the bone, favouring an anti-osteoclastogenesis milieu (Mohamad et al., 2021; Wong et al., 2018b).
Effects of tocotrienols on osteocytes
3.2
There are some preliminary studies on the effects of TT on osteocytes. Delta-TT protects osteocytes (MLO-Y4) from oxidative stress induced by tert-butyl hydroperoxide, a commonly used in vitro inducer of oxidative damage (Casati et al., 2020). Its cytoprotective properties combat oxidative stress by activating signalling pathways, such as PI3K/Akt and Nrf2. Through PI3K/Akt activation, delta-TT enhances cell survival signalling and promotes the phosphorylation of downstream targets that inhibit apoptosis (Casati et al., 2020). Simultaneously, through Nrf2 activation, it induces the expression of antioxidant defence genes, leading to increased production of protective enzymes. These coordinated actions result in a reduction of reactive oxygen species (ROS) levels by improving the glutathione (GSH)/glutathione disulfide (GSSG) ratio. GSH is a primary cellular antioxidant molecule, and maintaining a high GSH/GSSG ratio indicates a healthy cellular redox state (Vázquez-Meza et al., 2023).
However, in this study, the redox signalling was established in the osteoblast cell line rather than the osteocyte cell line. Although osteoblasts and osteocytes originate from the same lineage, they have distinct physiological roles and metabolic demands (Karthik and Guntur, 2021; Shen et al., 2022). Therefore, direct confirmation of these signalling mechanisms specifically in osteocytes would strengthen the conclusions about delta-TT’s protective effects in these cells.
The osteoporosis-reversing effects of emulsified and non-emulsified palm TT mixtures were investigated in a unique experimental design using ovariectomised rats (Mohamad et al., 2021). This study differed significantly from previous studies using TTs by employing a therapeutic rather than preventive approach, in which supplementation was initiated 2 months post-ovariectomy in a group of older rats (12 months old by the end of treatment). This experimental design is clinically relevant because it mimics the real-world scenario where osteoporosis treatment typically begins after significant bone loss has already occurred, rather than as a preventive measure.
The results showed no significant changes in the number of osteocytes and the number of empty lacunae of trabecular bone after a 2-month treatment (Mohamad et al., 2021). The lack of change in osteocyte numbers and empty lacunae suggests that the relatively short treatment duration (2 months) in rats with established, advanced bone loss may not have been sufficient to demonstrate recovery of the osteocyte population. Osteocyte turnover is a slow process; thus, the restoration of osteocyte networks takes considerable time (Lerebours and Buenzli, 2016). Empty lacunae represent sites of osteocyte death, and their persistence indicates ongoing or incompletely reversed bone deterioration.
Despite the lack of changes in osteocyte parameters, both emulsified and non-emulsified palm TT formulations significantly reduced skeletal SOST protein expression levels (Mohamad et al., 2021). This reduction in SOST expression is mechanistically important because a lower level would allow activation of Wnt/beta-catenin signalling in osteoblasts, potentially promoting bone formation (Delgado-Calle et al., 2017). This molecular change could represent an early adaptive response that precedes observable changes in bone structure and cell populations. This observation suggests that TTs can modulate osteocyte gene expression even in established osteoporosis, which may translate into improved bone formation with longer treatment duration.
The observation that both emulsified and non-emulsified palm TT produced similar effects on SOST expression suggests that the bioactive TT compounds themselves, rather than the delivery vehicle, are responsible for this molecular effect. The reduction in SOST represents a favourable shift in the bone remodelling environment that could potentially lead to increased bone formation if treatment were continued for a longer period.
In a subsequent study using a preventive model, ovariectomised rats were supplemented with emulsified and non-emulsified palm TT mixtures for 10 weeks, with treatment initiated 1 week after ovariectomy (Ekeuku et al., 2025). This preventive approach differs from the therapeutic approach described in the previous study, as treatment begins before substantial bone loss occurs, allowing assessment of TTs’ ability to prevent rather than reverse osteoporosis.
The results showed a significantly higher number of trabecular and cortical osteocytes than in both the palm TT-treated groups (Ekeuku et al., 2025). The preservation of osteocyte numbers is significant because osteocyte loss is a hallmark of postmenopausal osteoporosis, where oestrogen deficiency increases osteocyte apoptosis (McNamara, 2021).
A significantly lower Dmp1 gene expression was noted in the TT-treated group, but no significant changes in Sost and Dkk1 were observed (Ekeuku et al., 2025). The interpretation of reduced DMP1 expression is complex. The lower expression might indicate a more mature, stable osteocyte population, as DMP1 is highly expressed during osteocyte differentiation but may decrease in fully mature osteocytes (Kamiya and Takagi, 2004). The lack of changes in SOST and DKK1 expression in this study contrasts with the previous therapeutic study that showed reduced SOST expression. These observations, combined with preserved osteocyte numbers, suggest that in the preventive model, TTs primarily work by protecting osteocytes from oestrogen deficiency-induced apoptosis rather than by modulating Wnt inhibitor expression.
Metabolic syndrome has been shown to exert degenerative effects on bone health, particularly on the trabecular structures, by elevating inflammation (Wong et al., 2018a). In a study using rats with metabolic syndrome induced by high-fat high-carbohydrate diet, palm and annatto TT supplementation reduced the expression of several key osteocyte-derived factors, namely, SOST, DKK1, FGF23, and RANKL (Wong et al., 2019a).
The concurrent reduction in both SOST and DKK1 represents a coordinated decrease in Wnt pathway inhibition, which would synergistically enhance osteoblast differentiation and activity (Hu et al., 2024). FGF23 is often elevated in metabolic syndrome and obesity (Kawai, 2016). Its reduction suggests improved osteocyte metabolic health and reduced inflammatory stress. RANKL is the primary regulator of osteoclastogenesis (Weitzmann, 2013). Its reduction indicates a shift toward an anti-resorptive profile, with decreased osteoclast formation and activity. This would be expected to reduce bone resorption and help preserve bone mass.
These changes collectively enhance osteocyte-mediated signalling and shift towards an anti-resorptive profile, with reduced bone resorption (via decreased RANKL) and potentially increased bone formation (via decreased SOST and DKK1).
A subsequent study using the same metabolic syndrome model found decreased cortical osteocyte numbers, skeletal DMP1 and PHEX levels and increased empty lacunae numbers in rats with metabolic syndrome (Wong et al., 2022). These findings suggest that metabolic syndrome leads to osteocyte death and dysfunction, characterised by impaired expression of proteins crucial for bone mineralisation and phosphate metabolism.
Only annatto TT reversed the decreased empty lacunae, but palm TT did not (Wong et al., 2022). Despite these changes, DMP1 and PHEX remained unchanged with annatto or palm TT treatments (Wong et al., 2022). The differential effects between annatto and palm TTs on empty lacunae but not on DMP1/PHEX expression suggest that annatto TT may have superior antioxidant or anti-apoptotic properties that better protect osteocytes from metabolic syndrome-induced death. The composition of TT isomers differs between palm (rich in γ and α-TT) and annatto (predominantly δ-TT), which may account for functional differences. Additionally, preventing osteocyte death (by reducing empty lacunae) may not immediately restore DMP1 and PHEX expression, which might require a longer treatment duration or additional interventions.
Collectively, these studies suggest dual protective actions of TTs on bone health through osteocytes. Firstly, TTs function as potent cytoprotective agents by activating the PI3K/Akt and Nrf2 pathways, thereby preserving the integrity of the osteocyte lacunocanalicular network (Casati et al., 2020). This is particularly evident in preventive models, where early supplementation forestalls the apoptosis typically induced by oestrogen deficiency (Ekeuku et al., 2025). Secondly, TTs appear to modulate the osteocyte secretome by downregulating key Wnt inhibitors, specifically SOST and DKK1, across both established osteoporosis and metabolic syndrome models (Mohamad et al., 2021; Wong et al., 2019a). By downregulating these inhibitors, TTs facilitate the activation of Wnt/β-catenin signalling, thereby promoting an environment conducive to bone formation. Ultimately, these findings indicate that the primary bone-sparing effect of TTs may not rely solely on maintaining osteocyte numbers, but also on driving the remaining osteocytes toward a pro-anabolic and anti-resorptive profile by reducing RANKL and Wnt antagonists. The findings of these studies are summarised in Table 1.
These limited studies have their shortcomings. Firstly, most in vivo studies also use only one or two doses (60 mg/kg or 100 mg/kg), previously established to prevent bone loss in animal models (Chin, 2024b). A dose-dependent effect on osteocytes could not be established by these studies. Secondly, most studies used young and growing rats as samples. Despite being sexually mature, the skeleton of rats continues to grow until an advanced age (Fukuda and Iida, 2004). Any assault at this age (ovariectomy or diet modification) may produce stunted growth rather than degenerative changes as in osteoporosis. Thus, researchers continue to debate the relevance and representativeness of these models, despite their widespread use. Lastly, some outcomes were only measurable at the end of the experiment (e.g., osteocyte number and skeletal expression of critical markers), preventing the tracking of changes over time. Therefore, missing the window may explain the lack of changes between treatments and the osteoporosis control.
Perspectives
4
Given the limitations in the field, we propose several limitations for future researchers. Many studies rely on simple in vitro models that examine osteocytes and other bone cells separately, which can differ from the complex, coordinated bone environment in an organism. Overcoming this limitation would require a three-dimensional co-culture system or a bone explant, with mechanical loading platforms, which can help determine osteoblast migration more accurately by mimicking real bone environments. Other advancements in osteocyte culture, including osteocyte-in-chip and spheroid culture, would better mimic skeletal micro-environments and help in future investigations (Kim and Adachi, 2019; Nasello et al., 2020).
The barriers of direct observation of bone cells in vivo have recently been overcome with advances in microscopy. Multiphoton microscopy, when combined with genetically encoded fluorescent probes or locally injected nanoparticles (e.g., Cornell Prime Dots), enables deep tissue imaging of osteocytes in live animals, overcoming the optical scattering associated with mineralised bone (Matthews et al., 2023). Third harmonic generation microscopy, a label-free and non-invasive technique, enables high-resolution imaging of the osteocyte lacunar-canalicular network in live mice. It can distinguish osteocyte boundaries and canaliculi without the use of dyes or sectioning, and has been used to analyse osteocyte density and morphology in vivo (Tokarz et al., 2017). These techniques would enable the real-time tracking of TT’s action on osteocytes.
Future studies should also consider ablating the function of mature osteocytes to assess whether the bone-protective effects of TTs are retained. One example of such models is the DMP1-diphtheria toxin receptor (DMP1-DTR) mouse model. In this model, transgenic mice express the human DTR under the DMP1 promoter in osteocytes. Administration of diphtheria toxin (DT) selectively ablates 70%–80% of osteocytes in vivo, sparing osteoblasts (Tatsumi et al., 2007). Such a model can help to explain the role of osteocytes in TT’s skeletal action.
Furthermore, further studies should also explore how TTs coordinate multiple signalling pathways, rather than acting through a single molecular route. Fundamentally, connecting these findings to functional outcomes, such as mineralisation, perilacunar remodelling, bone strength, and phosphate regulation, will help us understand the therapeutic potential of TTs in enhancing overall bone quality.
Conclusion
5
In conclusion, studies have demonstrated that TTs exert protective and anabolic effects on bone health by influencing osteocytes and osteoblasts. TTs play a role in protecting osteocytes from oxidative damage and expression of their markers, but functional evidence remains lacking. Despite the established bone-protective effects of TTs on the skeletal system, further research is needed to elucidate their roles in modulating the activities of osteocytes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abd Manan N. Mohamed N. Shuid A. N. (2012). Effects of low-dose versus high-dose γ-Tocotrienol on the bone cells exposed to the hydrogen peroxide-induced oxidative stress and apoptosis. Evid. Based Complement. Altern. Med. 2012, 680834. 10.1155/2012/680834 22956976 PMC 3432387 · doi ↗ · pubmed ↗
- 2Aggarwal B. B. Sundaram C. Prasad S. Kannappan R. (2010). Tocotrienols, the vitamin E of the 21st century: its potential against cancer and other chronic diseases. Biochem. Pharmacol. 80, 1613–1631. 10.1016/j.bcp.2010.07.043 20696139 PMC 2956867 · doi ↗ · pubmed ↗
- 3Ardura J. A. Álvarez-Carrión L. Gortázar A. R. Alonso V. (2020). “Chapter 6 - linking bone cells, aging, and oxidative stress: osteoblasts, osteoclasts, osteocytes, and bone marrow cells,” in Aging. Editors PREEDYV. R. PATELV. B. , Second Edition (Academic Press).
- 4Casati L. Pagani F. Limonta P. Vanetti C. Stancari G. Sibilia V. (2020). Beneficial effects of δ-tocotrienol against oxidative stress in osteoblastic cells: studies on the mechanisms of action. Eur. J. Nutr. 59, 1975–1987. 10.1007/s 00394-019-02047-9 31280345 PMC 7351870 · doi ↗ · pubmed ↗
- 5Chen M. Li W. Lei L. Zhang L. (2025). Role of SOST in response to mechanical stimulation in bone and extraosseous organs. Biomolecules 15, 856. 10.3390/biom 15060856 40563496 PMC 12190277 · doi ↗ · pubmed ↗
- 6Chin K.-Y. (2024 a). “Recent progress on the skeletal research of tocotrienol,” in Lipophilic vitamins in health and disease. Editors TAPPIAP. S. SHAHA. K. DHALLAN. S. (Cham: Springer International Publishing).
- 7Chin K. Y. (2024 b). Updates in the skeletal and joint protective effects of tocotrienol: a mini review. Front. Endocrinol. (Lausanne) 15, 1417191. 10.3389/fendo.2024.1417191 38974581 PMC 11224474 · doi ↗ · pubmed ↗
- 8Chin K. Y. Mo H. Soelaiman I. N. (2013). A review of the possible mechanisms of action of tocotrienol - a potential antiosteoporotic agent. Curr. Drug Targets 14, 1533–1541. 10.2174/13894501113149990178 23859472 · doi ↗ · pubmed ↗