X chromosome-encoded microRNAs in immune regulation: sex differences and clinical implications
Valeria Lodde, Valentina Margarita, Myriam Gorospe, Ilaria Campesi

TL;DR
This paper explores how X chromosome-encoded microRNAs influence immune responses differently in males and females, potentially explaining sex differences in autoimmune diseases and immune function.
Contribution
The paper highlights the role of X chromosome-encoded miRNAs in immune regulation and their potential as sex-aware biomarkers and therapeutic targets.
Findings
X chromosome-encoded miRNAs are key regulators of immune homeostasis and are enriched in immune-related pathways.
Some X-resident miRNAs escape inactivation, leading to female-biased expression that may enhance immune reactivity and autoimmunity.
These miRNAs are proposed as potential biomarkers and therapeutic targets for sex-aware precision medicine.
Abstract
Sex-based differences in immune function influence susceptibility to infections and predisposition to autoimmunity, with women showing both stronger immune responses and a higher burden of autoimmune and chronic inflammatory diseases. While sex hormones contribute to these differences, accumulating evidence highlights a central role for the X chromosome, which is enriched in immune-related genes and subject to complex regulatory mechanisms such as X-chromosome inactivation, skewing, escape from inactivation, and imprinting. Within this context, X chromosome–encoded microRNAs (miRNAs) have emerged as key post-transcriptional regulators of immune homeostasis. The X chromosome harbors the highest density of miRNAs in the human genome, many of which target pathways involved in immune activation, tolerance, and tumorigenesis. Notably, some X-resident miRNAs escape X-chromosome inactivation,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
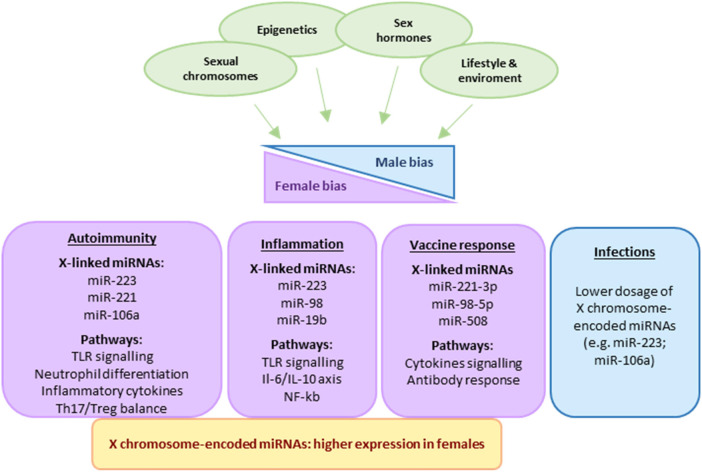 Figure 1
Figure 1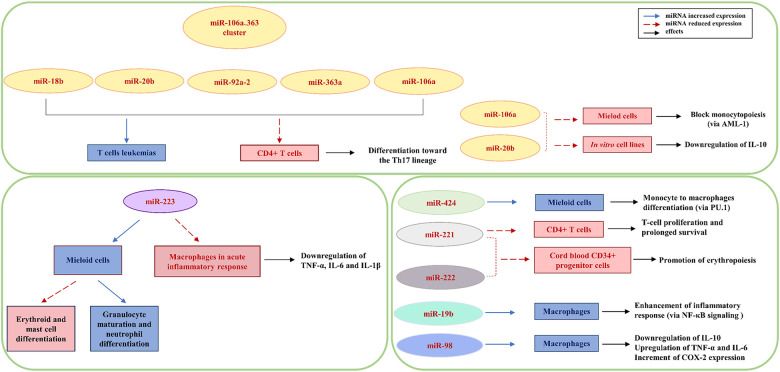 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Genetic Associations and Epidemiology · Reproductive System and Pregnancy
Introduction
Sex-based differences in immune function are among the most consistent and biologically significant forms of sexual dimorphism in humans, influencing both susceptibility to infectious diseases and predisposition to autoimmunity. Females generally exhibit stronger innate and adaptive immune responses compared to males, with enhanced antigen presentation, higher antibody titers following vaccination, and more robust activation of T and B lymphocytes (1). This heightened immune responsiveness provides superior protection against infectious agents but also contributes to a higher incidence and greater severity of autoimmune and chronic inflammatory diseases such as systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), multiple sclerosis (MS), and autoimmune thyroiditis, which affect women up to eight times more frequently than men (2–4).
Although sex hormones, particularly estrogens, play a central role in modulating inflammatory processes and promoting their resolution, sexual dimorphism in immune function emerges as early as during fetal development (5). In this regard, accumulating evidence indicates that genetic and epigenetic mechanisms linked to the X chromosome play an equally critical role in determining immune dimorphism (6). The X chromosome carries the highest density of immune-related genes in the human genome, including TLR7, TLR8, IL2Rγ, FOXP3, CXCR3, and CD40L, which encode key regulators of immune activation, tolerance, and inflammatory signaling (7, 8). In females, the presence of two X chromosomes introduces additional layers of genetic and epigenetic complexity. To ensure dosage compensation of genes residing on the X chromosome, one X chromosome in females undergoes random inactivation (X-chromosome inactivation, XCI) during early embryogenesis (9). However, an estimated 15% of X-linked genes escape XCI, potentially contributing to sexual dimorphism in X-linked gene expression. In addition to XCI escape, other regulatory mechanisms, including skewed XCI, characterized by nonrandom silencing of the X chromosome, and parent-of-origin-specific imprinting of genes located on the X chromosome, further influence sex-dependent differences in X-linked gene expression (10).
Within this chromosomal landscape, a considerable number of microRNAs (miRNAs) located on the X chromosome have been identified as critical modulators of immune homeostasis, further reinforcing the concept of a female immunological advantage (11). MiRNAs are small non-coding RNAs spanning 21–24 nucleotides that regulate gene expression post-transcriptionally. MiRNAs are transcribed as primary transcripts (pri-miRNAs) and processed by the microprocessor complex composed of Drosha and DGCR8, exported to the cytoplasm via exportin 5 (XPO5)/RanGTP, and finally cleaved by Dicer into mature miRNA duplexes that integrate into the RNA-induced silencing complex (RISC); upon integration, miRNAs suppress target protein production by either degrading the target mRNAs and/or suppressing its translation (12–17).
Through their impact on protein expression patterns, miRNAs regulate a broad range of biological processes, including cell proliferation, differentiation, apoptosis, and immune signaling, and their dysregulation contributes to the pathogenesis of numerous diseases, including cancer and autoimmune disorders (18). Notably, comprehensive genomic distribution analyses have demonstrated that the X chromosome exhibits the highest density of miRNA sequences among all chromosomes (19), with approximately 10% of known human miRNAs residing on the X chromosome (20). According to the miRBase database (https://www.miRBase.org), 118 miRNAs are mapped to the X chromosome, and 62 of them are validated as high-confidence entries as (Supplementary Table 1). A substantial proportion of these X-residing miRNAs are involved in immune regulation and tumorigenesis, two biologically interconnected processes that share common molecular pathways as well as in other essential cellular pathways. Using in silico miRNA target prediction algorithms, the possible functional roles of miRNAs residing on the X chromosome were assessed by identifying the pathways implicating proteins encoded by their target mRNAs. Supplementary Table 2 summarizes the pathway analysis of X-resident miRNAs. From this analysis, 75 signaling pathways were identified, of which approximately 23% are immune-related and 28% are associated with tumorigenesis. Among the pathways identified, representative immune- and tumor-related pathways include proteoglycans in cancer*,* viral carcinogenesis, endocytosis, bacterial invasion of epithelial cells, renal cell carcinoma and TGF-beta signaling pathway. Importantly, some X chromosome-resident miRNAs appear to escape XCI, resulting in female-biased expression patterns that may amplify immune reactivity and influence the balance between protection and pathology (21). Although a comprehensive and high-resolution map of miRNAs escaping XCI is still lacking, and direct quantitative evidence remains limited, accumulating data support the notion that a subset of X-resident miRNAs may evade silencing (21). Notably, several miRNAs are embedded within host genes that have been demonstrated to escape XCI, suggesting that these miRNAs may share the same epigenetic status (21). Examples include miR-548am, hosted within the CTPS2 gene, and miR-374 and miR-421, located within the long non-coding RNA FTX, both reported to escape XCI (22, 23).
In addition to genomic context, functional evidence also supports a role for specific X chromosome-resident miRNAs in XCI regulation (24). Lou et al. identified six miRNAs, miR-106a, miR-363, miR-340, miR-34b, miR-30e, and miR-181a, as regulators of XCI (24).
Overexpression of specific X chromosome-encoded miRNAs can potentiate pro-inflammatory signaling cascades or impair immune tolerance mechanisms, thereby contributing to the female predominance observed in autoimmune diseases (3). The interplay among hormonal factors, immune-related genes, and X-encoded miRNAs thus represents a critical axis in the regulation of sex-dependent immune responses (Figure 1).
Sex differences in immune responses arise from the combined influence of sex hormones, sex chromosomes, epigenetic mechanisms, and environmental factors. X-chromosome–encoded miRNAs, which are more highly expressed in females, contribute to female-biased immune activation and differential susceptibility to autoimmunity, inflammation, vaccine responses, and infections.
This minireview aims to provide a comprehensive overview of the emerging role of X chromosome-encoded miRNAs in the regulation of immune function, highlighting how their differential expression contributes to sex-based disparities in the incidence and severity of inflammatory and autoimmune diseases, and discussing their potential implications for the development of sex-specific diagnostic and therapeutic strategies.
Search strategy
A comprehensive literature search was performed using MEDLINE/PubMed and Google Scholar as the primary databases, supplemented by manual searches of additional scientific sources and the reference lists of key meta-analyses and review articles to ensure completeness. The search strategy targeted the following thematic domains: sex differences in inflammation, sex differences in immune responses, X chromosome genes and inflammation, X chromosome genes and immunity, miRNAs and immunity, miRNAs and inflammation, and X chromosome genes and miRNAs, vaccines, vaccination. A combination of Medical Subject Headings (MeSH) and free-text keywords was employed, applying Boolean operators (“AND” and “OR”) to optimize the sensitivity and specificity of the search. All identified records were initially screened based on titles and abstracts to assess relevance. Full-text versions of potentially eligible articles were retrieved and evaluated for inclusion according to their methodological quality and pertinence to the review topic. No restrictions were applied regarding publication year, while only articles written in English were considered. The literature search was last updated in January 2026, and the list of X-chromosome-encoded miRNAs was identified using miRBase (https://www.miRBase.org). The miRNA pathway interactions were explored using the DIANA miRPath v.3 web-based computational tool (http://snf-515788.vm.okeanos.grnet.gr/) (25). This software performs enrichment analysis of miRNA targets associated with each set of miRNA target genes in relation to KEGG pathways. KEGG pathway enrichment analysis enables the investigation of biological pathways and molecular interactions linked to target genes.
X chromosome-encoded miRNAs in immune regulation
Several X-linked miRNAs have been shown to exert critical functions in immunity and inflammation (Figure 2), potentially contributing to sex-dependent differences in immune responses as well as to variations in the dynamics and magnitude of the inflammatory reaction (11, 26).
Role of X-linked miRNAs in immunity and inflammation. The figure summarizes the involvement of different miRNAs in the regulation of immune cell differentiation, and inflammatory responses. Solid blue arrows indicate miRNAs increased expression, while dashed red arrows indicate reduced expression of selected miRNAs.
Among the X chromosome-resident miRNAs implicated in immune regulation, miR-223 is one of the most extensively studied. It is predominantly expressed in cells of the granulocytic lineage, with expression levels progressively increasing during granulocyte maturation and decreasing during erythroid and mast cell differentiation (27, 28). Functionally, miR-223 acts as a key regulator of neutrophil differentiation from myeloid precursors and serves as a negative modulator of the inflammatory response, particularly by fine-tuning the acute inflammatory response following pathogen recognition through Toll-like receptors (TLRs) (29). Four additional X-linked miRNAs, miR-106a, miR-424, miR-542, and miR-503, are implicated in hematopoietic lineage differentiation (11). MiR-106a has been shown to negatively regulate monocytopoiesis by targeting AML-1, a key inducer of monocyte differentiation and maturation (30). MiR-106a belongs to the miR-106a∼363 cluster, which also includes miR-18b, miR-20b, miR-19b-2, miR-92a-2, and miR-363, all residing on the X chromosome and involved in fine-tuning T helper cell differentiation toward the Th17 lineage (31). Notably, overexpression of these miRNAs has been reported in human T-cell leukemias (32). Furthermore, the X-resident microRNAs miR-424, miR-503 and miR-222, promote monocytic differentiation (33). In particular, miR-424 regulates the myeloid-specific transcription factor SPI1 (PU.1) and promoting monocyte to macrophage differentiation by targeting NFI-A (Nuclear Factor I A) (33).
MicroRNAs miR-221 and miR-222 are downregulated during erythropoiesis, and this reduction may promote erythropoiesis by the regulation of key functional proteins (34). MiR-221 suppresses genes involved in T-cell proliferation and survival, while inhibition of miR-221 results in enhanced T-cell proliferation and prolonged survival, suggesting that miR-221 functions as a negative feedback regulator to maintain immune homeostasis (35). In addition, miR-106a and miR-20b have been shown to modulate the expression of the anti-inflammatory cytokine IL-10 (36, 37). Furthermore, miR-19b promotes nuclear factor kappa B (NF-κB) signaling and is implicated in the enhancement of inflammatory responses (38). MiR-98 directly targets the 3′ untranslated region of IL-10 mRNA; accordingly, its overexpression suppresses TLR4-induced IL-10 production, enhancing COX-2 expression and increasing expression of pro-inflammatory cytokines, including TNF-α and IL-6, thereby amplifying the inflammatory response (39).
Taken together, these findings highlight the pivotal role of X chromosome-encoded miRNAs in regulating inflammatory pathways, myeloid cell differentiation, and innate immune responses. It is therefore plausible that dysregulation of these X-linked miRNAs contributes to the observed sex-based differences in susceptibility and severity of immune-mediated diseases.
X chromosome-encoded miRNAs in autoimmunity
Autoimmunity diseases (ADs) comprise a group of disorders in which the immune system aberrantly targets self-antigens, leading to chronic inflammation and subsequent tissue damage (40). Common ADs include RA, SLE, MS, type 1 diabetes mellitus (T1D), psoriasis, Graves’ disease, inflammatory bowel disease, and Sjögren's syndrome. The majority of ADs show a marked female predominance, with female-to-male ratios ranging from 2:1 to 25:1 in certain conditions (41). However, not all ADs occur exclusively or preferentially in females (42). MiRNAs play a key role in the pathogenesis of several ADs (43–48). Altered miRNA levels are observed in most ADs and have negative effects on B and T cells differentiation, homeostasis and activation, and differentiation of dendritic cells and macrophages via Toll-like receptors (49–52).
Rheumatoid arthritis
The abnormal expression of X-encoded miRNAs in RA patients has been analyzed mainly in the context of T cell differentiation, production of inflammatory cytokines, and B cell activation (53, 54). Imbalances in miR-223 have been reported in the immune cells of RA patients (55). MiR-223 is intensely expressed in RA synovium, regulates the differentiation of osteoclasts, and has important roles in the state of RA joints (56). Furthermore, Khalifa et al. reported that the expression levels of five X-encoded miRNAs (miR-221, miR-222, miR-532, miR-106a, and miR-98) were significantly different between peripheral blood mononuclear cells (PBMCs) of RA patients and healthy controls when stratifying by sex, and the expression levels of four miRNAs (miR-222, miR-532, miR-98, and miR-92a) were significantly different between RA females and males (57).
Other miRNAs encoded in the X chromosome were found to have an important role in RA. MiR-19a/b regulates the expression of TLR2 in RA synovial fibroblasts (RASFs), which are key effector cells in RA pathogenesis and contribute significantly to joint inflammation and destruction (58). Through the downregulation of TLR2, miR-19a/b reduces inflammatory responses by decreasing the production of IL-6 and MMP-3 (58). The expression levels of miR-92a were markedly reduced in RA synovial tissue and RASFs, suggesting its potential involvement in RA pathogenesis (59, 60). Restoration of miR-92a levels suppresses the proliferation and migration of RASFs, suggesting an inhibitory role of this miRNA in RA progression (61). Mechanistically, miR-92a was shown to inhibit cell proliferation and migration through the downregulation of AKT2, a member of the serine/threonine kinase family that is involved in key cellular processes such as metabolism, survival, and proliferation. Collectively, the miR-92a/AKT2 signaling axis plays a critical role in the pathogenesis of RA and may represent a promising therapeutic target for disease prevention and treatment (61). MiR-188-5p has been shown to target cell migration-inducing and hyaluronan-binding protein (CEMIP) and to indirectly regulate the expression of collagen type I alpha genes (COL1A1 and COL12A1) in RASFs. Reduced levels of miR-188-5p lead to enhanced activation and migration of RASFs (62). MiR-221 and miR-222 play an important role in the pathogenesis of RA. Elatta et al. reported that the expression levels of miR-221 and miR-222 were significantly upregulated in patients with RA, showing a positive correlation with disease activity (63). In particular, the overexpression of miR-221 has been shown to enhance the production of pro-inflammatory cytokines, promote the activation and migration of RASFs, and confer resistance to apoptosis (64).
Systemic lupus erythematosus
Alterations in miRNAs expression have been observed in various immune cells of patients with SLE. The first analysis of differentially expressed miRNAs in SLE was conducted by Dai et al., who identified distinct expression patterns in several miRNAs in PBMCs from SLE patients compared to healthy controls (65). Over the years, multiple studies have confirmed that SLE patients exhibit unique miRNA expression profiles, including circulating miRNAs (66, 67). Several X-encoded miRNAs are dysregulated in SLE. Notably, lower levels of miR-20, miR-106a, miR-92a, and miR-203 have been detected in the plasma of SLE patients (68). Moreover, the expression of miR-223 and miR-20 is significantly decreased in SLE patients with active nephritis (68). MiR-20 and miR-92a regulate apoptosis by repressing PTEN and, together with miR-106a, contribute to the control of monocytopoiesis and the regulation of regulatory T cells (30, 69).
MiR-221-5p has been identified as a potential biomarker of renal damage in SLE patients (70). The X-encoded microRNA miR-548m may target PTEN and contribute to SLE pathogenesis (71). Increased levels of miR-224 enhance STAT-1 expression in SLE T cells, promoting lupus nephritis (72). Additionally, Ming Zhao et al. reported that miR-505 is downregulated in T cells of SLE patients (73). All of these studies revealed an important role of X chromosome-encoded miRNAs in various molecular processes involved in disease development.
Multiple sclerosis
MS is an immune-mediated neuroinflammatory and demyelinating disease in which the role of miRNAs has been associated with disease onset, activity and duration (74–76). Indeed, miRNAs are highly expressed in cells of the immune system and central nervous system, where they regulate the expression of target genes in different cells. In this regard, the possible effects of miRNAs have been analysed for the MS-associated T cells, B cells, and macrophages (52, 77, 78).
Numerous investigations have detected dysregulated X chromosome-encoded miRNAs in MS patients. For instance, miR-18 and miR-20b are less abundant in relapse remitting MS patients when compared to healthy controls (79). Circulating X-encoded miRNAs such as miR-92a, miR-223 and miR-500, where detected in plasma, serum and cerebrospinal fluid (CSF) and correlate with disease type and severity (80–82). The increased levels of miR-424 in MS patients were associated with a reduction in the levels of CXCL10 and IRF1, as miR-424 is predicted to directly target their respective encoding mRNAs. The decrease in IRF1 and CXCL10 may contribute to a pro-survival phenotype in B cells in MS (83). Furthermore, the abundance of miR-448 also increased significantly in MS, and promoted Th17-mediated immune responses by targeting protein tyrosine phosphatase non-receptor type 2 (PTPN2) (84). In other examples, miR-92a levels negatively correlate with the number of lesions in the posterior inferior lobule and lateral temporal cortex (85), and miR-223 showed decreased serum levels in MS patients compared to healthy controls (86).
These data support the involvement of X-encoded miRNAs in the pathogenesis of ADs, suggesting that sex-linked differences in miRNA expression and regulation may contribute to the heightened susceptibility of women to autoimmunity. In particular, dysregulation of X chromosome-encoded miRNAs appears to influence key immune pathways implicated in RA, SLE, and MS.
MiRNAs and responses to vaccination
It is widely established that biological sexes differ in their responses to vaccination, although the mechanisms and clinical significance remain uncertain (1, 87). Females exhibit a higher magnitude of immune responses than males from birth to old age for a variety of vaccine candidates, such as inactivated diphtheria, hepatitis B, influenza, rabies, pertussis and pneumococcal vaccines, as well as the live measles vaccine (87). However, more recent evidence shows that sex differences in antibody titres are strongly age-dependent (88, 89), and systems-immunology studies show that these differences extend beyond humoral immunity to include innate and adaptive cell function, cytokine activity, and genetic–epigenetic regulation (90). Females tend to develop more frequent and sometimes more severe vaccine reactions than males, likely reflecting higher inflammatory response (87). They also report more local reactogenicity after influenza and COVID-19 vaccination, while systemic reactions are similar between sexes (87, 91). Conversely, post-authorization data indicate that serious adverse events, including life-threatening conditions and deaths after COVID-19 vaccination, are reported more often in males (91, 92), though these associations do not prove causality.
Sex difference in responses to vaccines could be influenced by multiple factors, including the X chromosome, sex hormones, and other biological mechanisms. In particular, some genes residing on the X chromosome escape XCI and are therefore expressed at higher levels in females, and miRNAs encoded on X chromosome may also contribute to these differences (10, 87). Some X-chromosome miRNAs are expressed more frequently in women due to incomplete XCI, which could contribute to sex differences in immune response to vaccines (21).
Although evidence directly linking X chromosome-residing miRNAs to vaccine responsiveness remains limited, recent studies have begun to uncover sex-dependent patterns of circulating miRNAs expression following immunization. In a cohort of healthcare workers receiving an mRNA-based COVID-19 vaccine, Anticoli et al. reported a sex-dependent miRNA signature, with miR-221-3p, an X-encoded miRNA, targeting molecules of the inflammatory pathways. Interestingly, vaccinated females showed higher miR-221-3p levels than unvaccinated females and vaccinated males, indicating that this upregulation may modulate the innate immune response to vaccine antigens (93). Similarly, the levels of miR-92a-2-5p in serum extracellular vesicles were shown to correlate with both antibody production and post-vaccination inflammatory responses in recipients of vaccines encoding the SARS-CoV-2 spike mRNA (94). Moreover, a recent study demonstrated that X-residing miR-508 and miR-98-5p showed the strongest positive correlation with vaccine-induced IgG antibody response against recombinant Ebola Zaire vaccine (rVSV-ZEBOV), although sex-stratified analyses were not conducted (95). MiR-508 is implicated in different diseases via PI3 K/AKT pathway regulation, affecting cell metabolism, growth, proliferation and survival, while miR-98-5p has been shown to regulate cytokine response via the NF-kB pathway and IL-6 signaling (95).
Collectively, these findings indicate that miRNAs may act as post-transcriptional regulators of vaccine-induced immunity and, interestingly, some of them have been identified as X chromosome resident miRNAs, potentially contributing to the observed sex differences in antibody and T-cell memory responses. However, the current evidence linking miRNAs to vaccine responsiveness remains largely preliminary. Available studies are limited by the smaller proportion of women in vaccine cohorts, the absence of extensive meta-analysis on sex differences, and insufficient statistical power due to limited group size, which together preclude definitive conclusions. In particular, investigations focusing on miRNAs encoded by the X-chromosome and their potential contribution to sex-related differences in long-term vaccine immunity remain scarce. Future research should therefore prioritize longitudinal, well-powered studies integrating sex, age, hormonal status, and functional immune parameters to better define the role of X-linked miRNAs in shaping vaccine-induced immune responses, and to explain individual variability in vaccine efficacy.
Conclusions
Chromosome X-resident miRNAs represent a pivotal axis in sex-dependent immunity, shaping inflammatory response, susceptibility to ADs, and vaccine responses. The emerging evidence linking X-encoded miRNAs to immune regulation highlights their potential as biomarkers of sex-dependent immune responsiveness and disease susceptibility. Circulating miRNA signatures could serve as minimally invasive indicators of vaccine efficacy and sex-dependent immune responses, shedding light on mechanisms underlying individual differences after vaccination or immune activation.
Beyond their value as biomarkers, miRNAs are being explored as direct therapeutic targets. Several miRNA-based strategies, including mimics and antagomiRs, have entered preclinical and clinical development (96), providing proof-of-concept for their translational potential. Notably, a miR-29 mimic has advanced into clinical trials for fibrotic diseases, based on its ability to limit tissue fibrosis (97, 98), and inhibitors of miR-155 have shown efficacy in preclinical models (99) demonstrating that modulating miRNA activity in vivo is feasible and can yield clinically meaningful outcomes. Moreover, they could, in principle, modulate immune reactivity in a sex-tailored manner.
However, it should be noted that research on miRNAs in vaccine responses remains relatively preliminary, with most studies being observational and limited in size or scope. Future research is required to validate the predictive and mechanistic roles of X chromosome-encoded miRNAs in vaccine-induced immunity. Such investigations could clarify the potential of miRNAs as modulators of sex-dependent vaccine efficacy and immune reactivity, while addressing current limitations in evidence.
Finally, translating these findings into clinical practice will require longitudinal, sex- and age-stratified studies that integrate genomic, hormonal, and epigenetic variables to validate the predictive and mechanistic value of X chromosome-encoded miRNAs. Critically, evaluations must consider the influence of age and distinct life stages in women, from infancy to menopause and older age, to avoid methodological biases and accurately capture sex-dependent immune dynamics (100, 101). Understanding the regulation of X chromosome-encoded miRNAs in this context promises to advance precision medicine approaches for vaccination, inflammatory disorders, and autoimmune diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Klein SL Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. (2016) 16(10):626–38. 10.1038/nri.2016.9027546235 · doi ↗ · pubmed ↗
- 2Migeon BR. Why females are mosaics, x-chromosome inactivation, and sex differences in disease. Gend Med. (2007) 4(2):97–105. 10.1016/S 1550-8579(07)80024-617707844 · doi ↗ · pubmed ↗
- 3Rio P Caldarelli M Miccoli E Guazzarotti G Gasbarrini A Gambassi G Sex differences in immune responses to infectious diseases: the role of genetics, hormones, and aging. Diseases. (2025) 13(6):1–24. 10.3390/diseases 13060179 PMC 1219188740558589 · doi ↗ · pubmed ↗
- 4Miquel CH Faz-Lopez B Guéry JC. Influence of X chromosome in sex-biased autoimmune diseases. J Autoimmun. (2023) 137:102992. 10.1016/j.jaut.2023.10299236641351 · doi ↗ · pubmed ↗
- 5Margarita V Lodde V Rappelli P Doro L Montella A Fiori PL The different innate immune response to infections in males and females emerges before birth. Life Sci. (2025) 369:123521. 10.1016/j.lfs.2025.12352140044031 · doi ↗ · pubmed ↗
- 6Migeon BR. X-linked diseases: susceptible females. Genet Med. (2020) 22(7):1156–74. 10.1038/s 41436-020-0779-432284538 PMC 7332419 · doi ↗ · pubmed ↗
- 7Bianchi I Lleo A Gershwin ME Invernizzi P. The X chromosome and immune associated genes. J Autoimmun. (2012) 38(2–3):J 187–92. 10.1016/j.jaut.2011.11.01222178198 · doi ↗ · pubmed ↗
- 8Mousavi MJ Mahmoudi M Ghotloo S. Escape from X chromosome inactivation and female bias of autoimmune diseases. Mol Med. (2020) 26(1):1–20. 10.1186/s 10020-020-00256-1PMC 772719833297945 · doi ↗ · pubmed ↗