Further dissection of Gardnerella vaginalis: description of Gardnerella lacydonensis sp. nov. (formerly genomic species 2), Gardnerella bretellae sp. nov. (formerly genomic species 9), Gardnerella massiliensis sp. nov. (formerly genomic species 14) and Gardnerella phocaeensis sp. nov
Priscille Allini Ntiguemassa, Mamadou Beye, Nicolas Orain, Sara Bellali, Stephane Alibar, Claudia Andrieu, Alissa Hammoud, Grégory Dubourg, Oleg Mediannikov, Anthony Levasseur, Florence Fenollar

TL;DR
This paper describes four new species of Gardnerella bacteria identified from clinical samples using genomic and phenotypic analysis.
Contribution
The study introduces four newly proposed Gardnerella species based on comprehensive genomic and phenotypic data.
Findings
Four new Gardnerella species were identified using genomic and phenotypic analysis.
Digital DNA-DNA hybridization and OrthoANI values confirmed the novelty of these species.
Each new species was assigned a unique name and type strain.
Abstract
Four bacterial strains – Marseille-Q9181T, Marseille-QA0894T, Marseille-Q9179T and Marseille-Q2328T – were isolated, the first three from vaginal samples and the last from a blood sample. MALDI-TOF MS analysis failed to assign these isolates to any known species within the genus Gardnerella. Consequently, phenotypic and taxonogenomic characterizations were conducted. All four strains were Gram-negative, non-motile and non-spore-forming coccobacilli. Strains Marseille-Q9181T, Marseille-Q2328T and Marseille-Q9179T were facultative anaerobes with microaerophilic capabilities, whereas Marseille-QA0894T was strictly anaerobic and microaerophilic. The dominant fatty acids (>10%) identified in strains Marseille-Q9181T, Marseille-QA0894T and Marseille-Q2328T were C16 : 0, C18 : 1 ω9c and C18 : 0. In strain Marseille-Q9179T, C18 : 0 was also among the most abundant fatty acids, along with the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
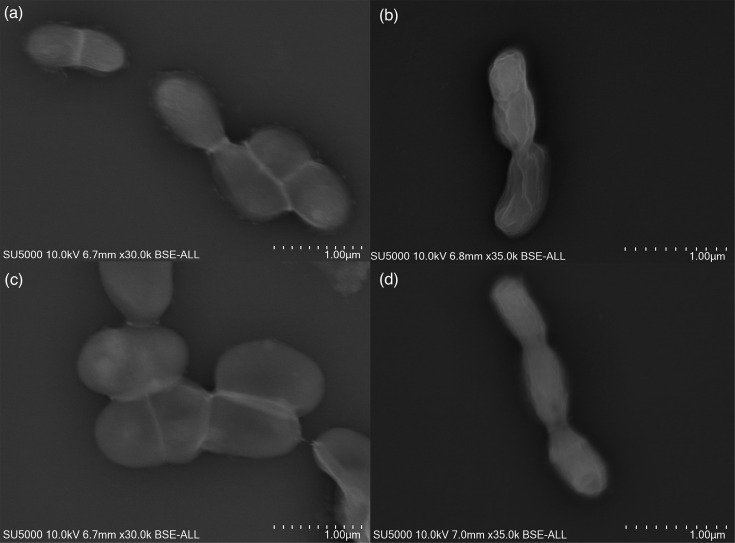 Fig. 1
Fig. 1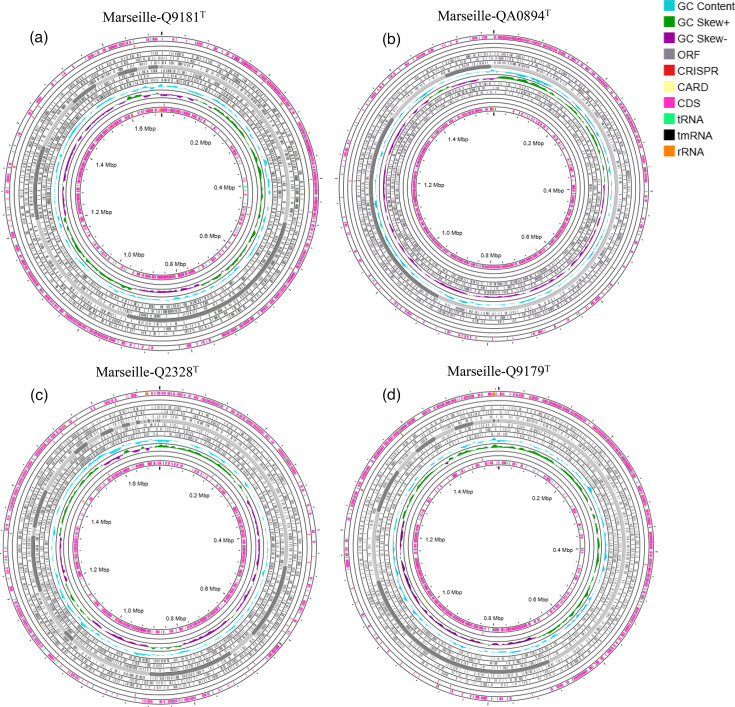 Fig. 2
Fig. 2 Fig. 3
Fig. 3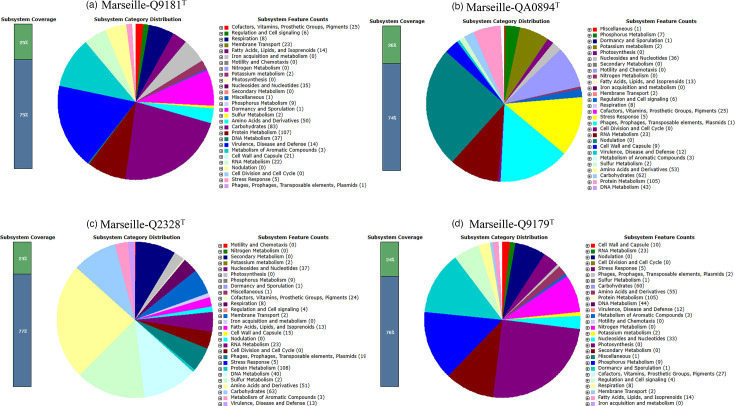 Fig. 4
Fig. 4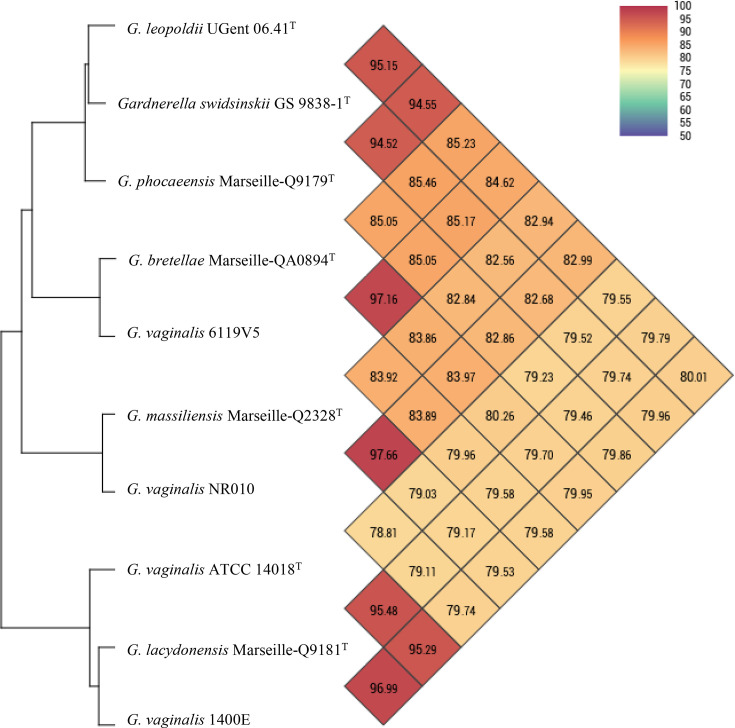 Fig. 5
Fig. 5|
| ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
| Vagina | Vagina | Blood | Vagina | Vagina | Vagina | Vagina | Vagina | Urine | Urine |
|
| 5% | 5% | 5% | 5% | 5% | 5% | 5% | 5% | 5% | 5% |
|
| Thick, whitish with regular edges | Rounded with irregular edges and whitish | Small, circular and flat, very transparent | Small, circular, whitish greyish with regular edges | Smooth surface from white to greyish | Smooth surface from white to greyish | Smooth surface from white to greyish | Smooth surface from white to greyish | Very small, smooth and circular, light grey, almost transparent | Small, circular and opaque white to greyish |
|
| Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli | Coccobacilli |
|
| 1.39×0.86 µm | 0.73×0.5 µm | 3×1.5 µm | 1×0.54 µm | 0.5×1.5 µm | 0.5×1.5 µm | 5×1.5 µm | 0.5×1.5 µm | 0.94 µm | 0.79 µm |
|
| Negative | Negative | Negative | Negative | Negative to Gram-variable | Negative to Gram-variable | Negative to Gram-variable | Negative to Gram-variable | Positive | Negative |
|
| This study | This study | This study | This study | [ | [ | [ | [ | [ | [ |
|
| ||||
|---|---|---|---|---|
|
| ||||
| Oxidase | − | − | − | − |
| Catalase | − | − | − | − |
|
| ||||
| + | + | + | + | |
| Ribose | − | − | − | − |
| + | + | + | + | |
| Lactose | + | + | + | + |
| + | + | + | + | |
| + | + | + | + | |
| Salicin | + | + | + | + |
| Aesculin | + | + | + | + |
| Iron citrate | + | + | + | + |
| Glycerol | + | + | + | + |
| + | + | + | + | |
| + | + | + | + | |
| + | + | + | + | |
| + | + | + | + | |
| − | − | − | + | |
| − | − | − | + | |
| Starch | + | + | + | + |
|
| ||||
| Acid phosphatase | + | + | + | + |
| Alkaline phosphatase | + | + | + | + |
| Valine arylamidase | + | + | + | + |
| Leucine arylamidase | + | + | + | + |
| Naphthol-AS-BI-phosphohydrolase | + | + | + | + |
| Esterase (C4) | − | − | − | + |
| Esterase lipase (C8) | − | − | − | + |
| Lipase (14) | − | − | − | + |
| ß-Galactosidase | + | − | − | − |
| α-Glucosidase | + | + | + | + |
| ß-Glucosidase | − | − | − | − |
| ß-Glucuronidase | − | − | − | + |
| + | − | − | − |
| Antibiotics | ||||||||
|---|---|---|---|---|---|---|---|---|
| MIC (µg ml−1) | S/R/I | MIC (µg ml−1) | S/R/I | MIC (µg ml−1) | S/R/I | MIC (µg ml−1) | S/R/I | |
|
| 0.047 | S | 0.125 | S | 0.032 | S | <0.064 | S |
|
| <0.016 | S | 0.094 | S | <0.016 | S | 0.094 | S |
|
| 0.047 | S | 0.25 | S | 0.25 | S | 1 | S |
|
| 0.25 | S | 0.5 | S | <0.016 | S | 0.25 | S |
|
| 0.094 | S | 0.5 | S | 0.008 | S | 0.125 | S |
|
| 0.25 | S | 0.5 | S | 2 | S | 0.5 | S |
|
| <0.016 | S | 0.25 | S | 0.047 | S | <0.016 | S |
|
| ≥256 | R | 16 | R | ≥256 | R | ≥256 | R |
|
| ≥256 | R | ≥256 | R | ≥256 | R | ≥256 | R |
|
| 16 | R | ≥256 | R | ≥256 | R | ≥256 | R |
|
| <0.016 |
| 0.023 |
| 0.064 |
| 0.032 |
|
|
| 0.5 |
| 0.75 |
| 1.5 |
| 2 |
|
|
| 0.25 |
| 0.5 |
| 32 |
| 32 |
|
| Fatty acids | Names | ||||||
|---|---|---|---|---|---|---|---|
| Mean relative % (a) | |||||||
| 16 : 0 | Hexadecanoic acid | 50.2±1.4 | 39.2±0.8 | 43.7±1.2 | 40.3±0.1 | 36.6±2.9 | 23.5±2.2 |
| 18 : 1 n9 | 9-Octadecenoic acid | 15.3±1.1 | 20±1.2 | 28.5±0.4 | 18±1 | 35.2±2.2 | 44.3±3.3 |
| 18 : 0 | Octadecanoic acid | 12.2±0.2 | 15.1±0.9 | 12.8±0.4 | 13.9±2 | 9.2±1.4 | 3.8±0.5 |
| 14 : 0 | Tetradecanoic acid | 7.7±0.1 | 7.6±0.9 | 4.7±0.1 | 12.3±2.4 | 6.9±1.5 | 4.6±0.4 |
| 18 : 2 n6 | 9,12-Octadecadienoic acid | 5.4±0.5 | 10.1±0.1 | 5.1±0.6 | 7.7±0.8 | 4.8±1.3 | 8±1.5 |
| 10 : 0 | Decanoic acid | 2.3±0.4 | 1±0.3 |
| 1.2±0.1 |
|
|
| 18 : 1 n7 | 11-Octadecenoic acid | 1.5±0.2 | 1.1±0.1 |
|
| 2.5±2 | 4.4±2.3 |
| 17 : 0 | Heptadecanoic acid |
| 1±0.1 |
|
|
|
|
| 12 : 0 | Dodecanoic acid | 1.8±0.6 |
|
|
|
|
|
| 15 : 0 | Pentadecanoic acid | 1±0.2 |
| 1.1±0.1 |
|
|
|
| 16 : 1 n7 | 9-Hexadecenoic acid |
|
|
|
| 3±0.9 | 6.2±0.5 |
| 19 : 0 | Nonadecanoic acid |
|
|
|
|
| 1.1±0.6 |
| 18 : 1 n5 | 13-Octadecenoic acid |
|
|
|
|
|
|
| 17 : 0 anteiso | 14-Methyl-hexadecanoic acid |
|
|
|
|
|
|
| 20 : 4 n6 | 5,8,11,14-Eicosatetraenoic acid |
|
|
|
|
|
|
| 16 : 1 n9 | 7-Hexadecenoic acid |
|
|
|
|
|
|
| 17 : 1 n7 | 10-Heptadecenoic acid |
|
|
|
|
|
|
| 15 : 0 anteiso | 12-Methyl-tetradecanoic acid |
|
|
|
|
|
|
| 17 : 0 iso | 15-Methyl-hexadecanoic acid |
|
|
|
|
|
|
| 16 : 0 iso | 14-Methyl-pentadecanoic acid |
|
|
|
|
|
|
| 15 : 0 iso | 13-Methyl-tetradecanoic acid |
|
|
|
|
| |
| 19 : 01 | Nonadecenoic acid |
|
|
|
|
|
|
| 08 : 0 | Octanoic acid |
|
|
|
|
|
|
| 14 : 0 iso | 12-Methyl-tridecanoic acid |
|
|
|
|
|
|
| References | This study | This study | This study | This study | [ | [ | |
| Genome features |
| |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
| 1,702,271 | 1,505,165 | 1,720,610 | 1,565,522 | 1,667,406 | 1,563,545 | 1,622,089 | 1,514,270 | 1,511,834 | 1,509,345 |
|
| 12 | 12 | 32 | 12 | 1 | 1 | 9 | 5 | 11 | 3 |
|
| 41.4 | 43.2 | 45.3 | 42.3 | 41.4 | 42.1 | 41.9 | 42.5 | 43.3 | 42.5 |
|
| 1,374 | 1,186 | 1,380 | 1,227 | 1,320 | 1,239 | 1,327 | 1,210 | 1,176 | 1,204 |
|
| 1,323 | 1,138 | 1,331 | 1,178 | 1,268 | 1,187 | 1,277 | 1,159 | 1,127 | 1,155 |
|
| 51 | 48 | 49 | 49 | 52 | 52 | 50 | 51 | 49 | 49 |
|
| 5 | 3 | 3 | 3 | 6 | 6 | 4 | 5 | 3 | 3 |
|
| 45 | 44 | 45 | 45 | 45 | 45 | 45 | 45 | 45 | 45 |
|
| 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
|
| 0 | 0 | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 0 |
|
| 439,839 | 304,156 | 166,989 | 425,876 | 1,667,384 | 948,424 | 432,172 | 1,406,824 | 472,464 | 1,398,357 |
|
| 2 | 3 | 4 | 2 | 1 | 1 | 2 | 1 | 2 | 1 |
|
|
| Code | COG function | ||||
|---|---|---|---|---|---|
| [A] | RNA processing and modification | 1 | 1 | 1 | 1 |
| [B] | Chromatin structure and dynamics | 0 | 0 | 0 | 0 |
| [C] | Energy production and conversion | 24 | 23 | 23 | 23 |
| [D] | Cell cycle control, cell division, chromosome partitioning | 15 | 15 | 16 | 14 |
| [E] | Amino acid transport and metabolism | 62 | 65 | 64 | 65 |
| [F] | Nucleotide transport and metabolism | 49 | 50 | 52 | 47 |
| [G] | Carbohydrate transport and metabolism | 124 | 78 | 74 | 77 |
| [H] | Coenzyme transport and metabolism | 40 | 35 | 43 | 42 |
| [I] | Lipid transport and metabolism | 28 | 25 | 25 | 27 |
| [J] | Translation, ribosomal structure and biogenesis | 144 | 144 | 142 | 142 |
| [K] | Transcription | 49 | 39 | 42 | 39 |
| [L] | DNA replication, recombination and repair | 64 | 63 | 80 | 61 |
| [M] | Cell wall/membrane/envelope biogenesis | 76 | 72 | 72 | 69 |
| [N] | Cell motility | 3 | 2 | 2 | 2 |
| [O] | Posttranslational modification, protein turnover, chaperones | 51 | 49 | 53 | 46 |
| [P] | Inorganic ion transport and metabolism | 40 | 37 | 37 | 39 |
| [Q] | Secondary metabolite biosynthesis, transport and catabolism | 2 | 1 | 1 | 2 |
| [R] | General function prediction only | 43 | 37 | 37 | 39 |
| [S] | Function unknown | 24 | 24 | 24 | 21 |
| [T] | Signal transduction mechanisms | 28 | 27 | 27 | 27 |
| [U] | Intracellular trafficking, secretion and vesicular transport | 8 | 8 | 8 | 8 |
| [V] | Defence mechanisms | 47 | 43 | 39 | 44 |
| [X] | Mobilome: prophages, transposons | 9 | 18 | 29 | 7 |
| [Y] | Nuclear structure | 0 | 0 | 0 | 0 |
| [Z] | Cytoskeleton | 0 | 0 | 0 | 0 |
| – | Not in COGs | 128 | 119 | 126 | 113 |
| Query strains | dDDH (%) | OrthoANI (%) | Difference G+C% | |
|---|---|---|---|---|
|
| 61.5 | 95.48 | 0.04 | |
| 72.1 | 96.99 | 0.22 | ||
| 32.1 | 87.18 | 1.08 | ||
| 32.2 | 86.85 | 1.07 | ||
| 23 | 79.79 | 0.7 | ||
| 26.8 | 79.74 | 0.44 | ||
| 27.6 | 79.68 | 1.83 | ||
| 26.3 | 79.79 | 1.9 | ||
| 26 | 79.58 | 1.86 | ||
| 27 | 79.64 | 1.56 | ||
| 30.2 | 85.89 | 1.87 | ||
| 27.3 | 78.15 | 3.39 | ||
| 28.7 | 79.73 | 0.59 | ||
| 26.5 | 79.11 | 4.04 | ||
| 26.5 | 79.7 | 1.79 | ||
| 26.7 | 79.17 | 3.85 | ||
| 27.1 | 79.46 | 0.85 | ||
|
| 27.5 | 80.26 | 1.83 | |
| 27 | 79.95 | 2.01 | ||
| 26.7 | 79.42 | 0.71 | ||
| 26.7 | 79.62 | 0.75 | ||
| 28.2 | 85.23 | 1.1 | ||
| 29.5 | 85.46 | 1.35 | ||
| 30.8 | 85.22 | 0.26 | ||
| 58.9 | 95.02 | 0.11 | ||
| 72.4 | 97.16 | 0.07 | ||
| 60.2 | 95.31 | 0.24 | ||
| 26.9 | 79.68 | 0.08 | ||
| 26.2 | 78.45 | 5.18 | ||
| 28.4 | 81.12 | 2.38 | ||
| 29 | 83.97 | 2.24 | ||
| 29.3 | 83.92 | 2.06 | ||
| 29 | 85.05 | 0.94 | ||
|
| 26.8 | 79.03 | 0.58 | |
| 28.1 | 79.53 | 4.07 | ||
| 26.5 | 78.79 | 2.77 | ||
| 26.7 | 78.8 | 2.81 | ||
| 28.1 | 82.94 | 3.16 | ||
| 27.2 | 82.56 | 3.41 | ||
| 27.7 | 83.16 | 2.32 | ||
| 29.1 | 83.65 | 1.95 | ||
| 29.4 | 83.92 | 1.99 | ||
| 28.4 | 83.37 | 2.3 | ||
| 26.5 | 78.91 | 1.98 | ||
| 27.7 | 77.91 | 7.28 | ||
| 30.1 | 80.5 | 4.45 | ||
| 75.5 | 97.66 | 0.18 | ||
| 27.4 | 82.84 | 3 | ||
|
| 26.6 | 79.23 | 1.9 | |
| 26.9 | 79.86 | 1.07 | ||
| 26.3 | 79.09 | 0.23 | ||
| 26.2 | 78.93 | 0.18 | ||
| 57.3 | 94.55 | 0.16 | ||
| 57.6 | 94.52 | 0.42 | ||
| 28.5 | 84.01 | 0.68 | ||
| 28.9 | 85.09 | 1.05 | ||
| 28.6 | 85.05 | 1.01 | ||
| 28.7 | 84.9 | 0.7 | ||
| 26.1 | 78.89 | 1.02 | ||
| 26 | 78.42 | 4.25 | ||
| 31.1 | 82.35 | 1.45 | ||
| 27.2 | 82.86 | 3.18 |
- —http://dx.doi.org/10.13039/501100001665 Agence Nationale de la Recherche
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive tract infections research · Bacterial Identification and Susceptibility Testing · Genomics and Phylogenetic Studies
Introduction
Gardnerella vaginalis is a facultative anaerobic bacterium associated with bacterial vaginosis [12]. Different classification systems for G. vaginalis (biotyping, genotyping and sub-grouping) have been previously proposed [36]. However, some Gardnerella isolates have never been identified using clade-specific qPCR assays [4,79]. Additionally, analysis of the V4 hypervariable region of the 16S rRNA gene revealed that for over 98.5% of Gardnerella species, the V4 region was identical [10]. In order to resolve their taxonomic position, 81 genomes of G. vaginalis were analysed in 2019. Based on an average nucleotide identity of 96% and DNA–DNA hybridization of 70% or higher, strains of G. vaginalis were classified into several genomic species, with description of six species: G. vaginalis sensu stricto, and five ex-genomic species as Gardnerella leopoldii, Gardnerella piotii, Gardnerella swidsinskii, Gardnerella pickettii and Gardnerella greenwoodii [1011]. Other genomic studies, using a larger number of genomes, have revealed the presence of additional genotypes [12], including fourteenth genomic species (Gardnerella NR010 genomic species 14), with an average nucleotide identity value lower than or equal to 96% compared to other genomes, that is compatible to a new species [1314]. Despite the number of molecular and genomic differences [715] among genotypes, some have never been officially described as new species, notably genomic species 2, 7, 9, 10, 11, 12, 13 and 14. Here, we demonstrate genetic and biological differences and describe four species of the genus Gardnerella: Gardnerella lacydonensis, Gardnerella bretellae and Gardnerella massiliensis, known as genomic species 2, genomic species 9 and genomic species 14, respectively, as well as Gardnerella phocaeensis, a new potential species, not attributed previously to any genomic species. We used phenotypic, taxonomic and genomic approaches to characterize these species.
Methods
Ethical considerations and sampling
The four strains were isolated from four specimens (three vaginal swabs and one blood culture) taken during patient care and sent for diagnostic purposes to the clinical microbiology laboratory at our university hospital (AP-HM, at the Institut Hospitalo-Universitaire Méditerranée Infection, Marseille, France). As permitted by French law (Article L.1211-2 of the Public Health Code), patients have been informed of the possible re-use of their samples and personal data collected during treatment for research purposes. They had the option of objecting by notifying the AP-HM Data Protection Officer, but none of the four patients indicated any objections.
Bacterial colony identification
Bacterial colonies from vaginal swabs and blood samples were isolated on Columbia agar enriched with 5% sheep blood (bioMérieux, Marcy-l'Etoile, France) in an anaerobic atmosphere using a GasPak^™^ EZ Anaerobe gas generating system (Becton Dickinson, Franklin Lakes, NJ, USA) for 48 h. Prior to this, vaginal samples were incubated at 5% CO_2_ for 48 h at 37 °C on three culture media: PolyViteX chocolate agar, PolyViteX VCAT3 chocolate agar and Columbia CNA agar, which was enriched with 5% sheep blood (bioMérieux), whereas the blood sample was incubated in an anaerobic blood culture bottle for 28 h. Subsequently, all bacterial colonies were identified by MALDI-TOF MS using the Microflex LT tool instrument (Bruker Daltonics, Bremen, Germany). A score of >2 indicated correct species identification, while a score of ≥1.7 but <2 indicated correct genus identification to identify colonies of the genus Gardnerella [1617].
Phenotypic characteristics
Growth characteristics
Gram staining of cell was performed using a Color Gram 2 kit (bioMérieux). The staining was evaluated under a Motic Panthera C2 trinocular microscope with a 100X oil immersion lens. The images were produced using Mosaic software version 3.0.7. Cell morphology of the four strains was evaluated by scanning electron microscopy (SEM) using an SU5000 microscope (Hitachi High-Tech Corporation, Tokyo, Japan) [18]. Sporulation and motility were tested as previously described [18]. Moreover, optimal growth conditions were determined on Columbia agar enriched with 5% sheep blood (bioMérieux) for 72 h at different growth temperatures (28, 37, 45 and 56 °C) and different atmospheres: aerobic, anaerobic using the GasPak^™^ EZ Anaerobe gas generation system (Becton Dickinson) and microaerophilic using the GasPak^™^ Campy pocket system (Becton Dickinson). Salt tolerance and ability to grow under different pH conditions were assessed after 24, 48 and 72 h of incubation at 37 °C under anaerobic conditions, using 37 g brain heart infusion broth (Becton Dickinson) supplemented with 5 g yeast extract, 1 g starch, 1 g peptone and 21 g Columbia agar (Thermo Fisher Scientific, Waltham, MA, USA) for 1 l. NaCl was added to obtain salt concentrations of 5%, 7%, 10%, 15%, 20%, 25% and 30%. NaOH or HCl was adjusted to 4.5, 5.5, 6.5, 7.5 and 8.5 to achieve the pH values tested.
Biochemical tests
Catalase and oxidase activity were assayed using a hydrogen peroxide solution (bioMérieux) and oxidase discs (Becton Dickinson), respectively, as previously described [19]. Carbohydrate metabolism and enzymatic activities of all strains were analysed using API^®^ ZYM, API^®^ 50 CH and API^®^ 20 A strips (bioMérieux). Cellular fatty acid methyl esters were analysed by GC/MS as already reported [20].
Antibiotic susceptibility
Amoxicillin, amoxicillin/clavulanic acid, ticarcillin/clavulanic acid, ceftriaxone, imipenem, vancomycin, colistin, tobramycin, azithromycin, ciprofloxacin, clindamycin, doxycycline and metronidazole were tested for antibiotic susceptibility using E-test strips (bioMérieux). Mueller–Hinton agar supplemented with 5% sheep blood (bioMérieux) was used to test antibiotic susceptibility. The manufacturer’s instructions were followed to read the MIC, which was then interpreted in accordance with the guidelines of the European Committee on Antimicrobial Susceptibility (https://www.eucast.org/) consulted in January 2025 and the CLSI M100 guidelines. When breakpoint values were available, antibiotic susceptibility was interpreted according to the following specific categories: susceptible (S), susceptible, increased exposure (I) and resistant (R). In the absence of breakpoint values and when MICs were high, as for colistin and tobramycin, strains were considered resistant. In other cases (azithromycin, ciprofloxacin and doxycycline), MICs could not be interpreted due to the absence of breakpoint values.
Whole-genome sequencing and bioinformatics analysis
Genomic DNA from the four strains was extracted with the EZ1 instrument (Qiagen, Hilden, Germany) using the EZ1 DNA Tissue kit (Qiagen) and was sequenced on the MiSeq sequencer (Illumina Inc, San Diego, CA, USA) using a Nextera XT Paired End sample preparation kit (Illumina) [21]. Assembled reads were obtained using SPAdes 3.11.1 software [22]. Scaffolds of less than 800 bp and depth values of less than 25% of the median depth were excluded during assembly, and CheckM was used to evaluate the quality of genome assemblies [23]. Genomic sequences for the four strains were annotated using Prokka version 1.14.6 and Rapid Annotation Using Subsystem Technology (RAST) [2425]. In addition, we performed a BLASTp analysis against the Clusters of Orthologous Genes (COGs) database [26]. Genome maps were constructed using the CGview/Proksee web server [27]. The genomes of the four Gardnerella strains were compared to the closely related Gardnerella genomic species described in previous studies [1014].
The 16S rRNA sequences of the 4 strains analysed and the genomes of 14 Gardnerella species (Table S1, available in the online Supplementary Material) were aligned using muscle [28]. mega version 11 software was used to construct the phylogenetic tree with 1,000 bootstrap replications using the neighbour-joining method and the Tamura 3-parameter method [2932]. According to digital DNA–DNA hybridization (dDDH) and the average nucleotide identity (OrthoANI), the degree of pairwise similarity between the genomes was compared with the 14 genomic species of the genus Gardnerella [3334]. The dDDH was calculated by downloading the genome sequences from the Genome-to-Genome Distance Calculator 2.1 (GGDC) web server (http://ggdc.dsmz.de/) consulted in January 2025 [35]. OrthoANI was analysed using OAT version 0.93.1 software, and a heat map was generated [36].
CRISPR loci and CRISPR-Cas clusters with an evidence level of 3 or 4 were analysed using regularly spaced short palindromic repeats (CRISPR). Prophage sequences were identified using the PHASTEST [37]. The insertion sequences and replicon of the plasmids were determined using the ISfinder and PlasmidFinder [3839]. BAGEL software was used to predict bacteriocins [40], while secondary metabolites were estimated using antiSMASH 5.0 [41]. Genes coding for vaginolysin and sialidase were identified via BLASTp with an identity threshold of 40% and a minimum length of 30% against two databases created with protein sequences of different types of vaginolysin (Type 1A, Type 1B, Type 1C, Type 2 and Type 3) [42] and different sequences of sialidase (nanH1, nanH2 and nanH3) [43]. The vaginolysin protein sequences were aligned using muscle [28]. Structural prediction of the candidate proteins vaginolysin and sialidase was performed using PHYRE 2.2 [44].
Results
Strain identification by MALDI-TOF MS Biotyper
Strains Marseille-Q9181^T^, Marseille-QA0894^T^ and Marseille-Q9179^T^ were isolated from vaginal samples, while strain Marseille-Q2328^T^ was isolated from a blood sample. MALDI-TOF MS analysis failed to identify the Gardnerella strains to the species level. Strains Marseille-Q9181^T^ and Marseille-Q2328^T^ were identified as G. vaginalis with scores of 2.01–2.1 and 2.29–2.23, respectively. Strains Marseille-QA0894^T^ and Marseille-Q9179^T^ were identified as G. leopoldii_swidsinskii with scores of 2.18–2.33 and 2.18–2.46, respectively.
Phenotypic characterization
The phenotypic characteristics of the four Gardnerella strains were compared with those of other described Gardnerella species with validly published names, namely G. vaginalis, G. leopoldii, G. piotii, G. swidsinskii [10], G. pickettii and G. greenwoodii [11].
Bacterial growth and morphological observations
Growth of the four strains was observed on Columbia agar enriched with 5% sheep blood (bioMérieux), after 48 h of incubation at an optimum temperature of 37 °C in microaerophilic and anaerobic atmospheres. Aerobic growth was only observed for strains Marseille-Q2328^T^ and Marseille-Q9179^T^. Strains Marseille-Q2328^T^ and Marseille-QA0894^T^ grew at a pH ranging from 6.5 to 8.5 (optimum 7.5), while strains Marseille-Q9181^T^ and Marseille-Q9179^T^ grew at a pH ranging from 7.5 to 8.5 (optimum 7.5). Growth was observed at 0.5% NaCl for all four isolates. Regarding morphological description, strain Marseille-Q9181^T^ exhibited thick, whitish bacterial colonies with regular edges. Strain Marseille-QA0894^T^ featured domed colonies with irregular edges and a whitish colour. Strain Marseille-Q2328^T^ exhibited small, circular, flat and transparent colonies. Strain Marseille-Q9179^T^ showed small, circular, whitish-greyish colonies with regular margins.
Gram staining and SEM
All four strains stained consistently as Gram-negative coccobacilli (Fig. S1), as well as non-spore-forming and non-motile. SEM micrographs revealed that the cells of strain Marseille-Q9181^T^ measured 1.39±0.4 µm in length and 0.86±0.07 µm in width (Fig. 1a); strain Marseille-QA0894^T^ measured 0.73±0.13 µm in length and 0.5±0.06 µm in width (Fig. 1b); strain Marseille-Q2328^T^ measured 3±0.64 µm in length and 1.5±0.09 µm in width (Fig. 1c); and strain Marseille-Q9179^T^ measured 1±0.05 µm in length and 0.54±0.04 µm in width (Fig. 1d). The growth characteristics of strains Marseille-Q9179^T^, Marseille-Q2328^T^, Marseille-QA0894^T^ and Marseille-Q9179^T^, compared with the most closely related species of the genus Gardnerella, are listed in Table 1.
SEM micrographs of (a) G. lacydonensis Marseille-Q9181T, (b) G. bretellae Marseille-QA0894T, (c) G. massiliensis Marseille-Q2328T and (d) G. phocaeensis Marseille-Q9179T. SEM acquisition settings appear on each micrograph as follows: instrument, accelerating voltage, working distance, magnification, detector and scale.
Biochemical characterization, antibiotic susceptibility and fatty acids
Strains Marseille-Q2328^T^, Marseille-QA0894^T^, Marseille-Q9181^T^ and Marseille-Q9179^T^ were negative for catalase and oxidase. Using the API^®^ 20 A strip, positive reactions were observed for d-glucose, d-mannitol, lactose, d-sucrose, d-maltose, salicin, aesculin, iron citrate, glycerol, d-cellobiose, d-mannose, d-melezitose and d-sorbitol for all four strains. l-Rhamnose and d-trehalose were positive only for strain Marseille-Q9179^T^. Using the API^®^ 50 CH strip, all four strains were positive for starch. Using API^®^ ZYM strips, all four strains were positive for alkaline phosphatase, leucine arylamidase, valine arylamidase, acid phosphatase, naphthol-AS-BI-phosphohydrolase and α-glucosidase. Strain Marseille-Q9179^T^ was positive for esterase (C4), esterase lipase (C8), lipase (C14) and ß-glucuronidase. Strain Marseille-Q9181^T^ was also positive for ß-galactosidase and d-mannosidase. These results are summarized in Table 2.
MICs for the four strains are shown in Table 3. Briefly, strains Marseille-Q9181^T^, Marseille-QA0894^T^, Marseille-Q2328^T^ and Marseille-Q9179^T^ were susceptible to clindamycin but resistant to metronidazole. The fatty acid composition of the four strains is presented in Table 4 and compared with G. vaginalis strains [45]. The most abundant fatty acids (representing >20% of the total) for strain Marseille-Q2328^T^ were C_16 : 0_ (43.7%) and C_18 : 1_ ω9c (28.5%). The most prevalent fatty acid for strain Marseille-Q9181^T^ was C_16 : 0_ (50.2%). For strain Marseille-QA0894^T^, the major fatty acids were C_16 : 0_ (39.2%) and C_18 : 1_ ω9c (20%). For strain Marseille-Q9179^T^, the most abundant fatty acid was C_16 : 0_ (40.3%).
Genomic characterization
Genome properties
The genome length of strain Marseille-Q9181^T^ is 1,702,271 bp, assembled into 12 contigs, with a G+C content of 41.4 mol%. The gene prediction analysis of strain Marseille-Q9181^T^ reports 1,374 predicted genes, including 1,323 protein-coding genes, 51 RNA genes (5 rRNA, 45 tRNA and 1 tmRNA) and 7 pseudogenes (Table 5 and Fig. 2a). Strain Marseille-QA0894^T^ has a genome length of 1,505,165 bp, assembled into 12 contigs, with a G+C content of 43.2 mol%. Strain Marseille-QA0894^T^ comprises 1,138 predicted genes, including 1,207 protein-coding genes, 49 RNA genes (3 rRNA, 45 tRNA and 1 tmRNA) and 5 pseudogenes (Table 5 and Fig. 2b). The genome length of strain Marseille-Q2328^T^ is 1,720,610 bp, assembled into 32 contigs, with a G+C content of 45.3 mol%. Strain Marseille-Q2328^T^ has 1,380 predicted genes, with 1,331 protein-coding genes, 49 RNA sequences (3 rRNA, 45 tRNA and 1 tmRNA), 7 pseudogenes and 1 repeat region (Table 5 and Fig. 2c). Strain Marseille-Q9179^T^ has a genome length of 1,565,522 bp, assembled into 12 contigs, with a G+C content of 42.3 mol%. Strain Marseille-Q9179^T^ has 1,227 predicted genes, including 1,178 protein-coding genes, 49 RNA sequences (3 rRNA, 45 tRNA and 1 mRNA), 4 pseudogenes and 1 repeat region (Table 5 and Fig. 2d).
Circular genome map of strains (a) G. lacydonensis Marseille-Q9181T, (b) G. bretellae Marseille-QA0894T, (c) G. massiliensis Marseille-Q2328T and (d) G. phocaeensis Marseille-Q9179T generated by the CGView server.
Phylogenetic tree
The phylogenetic 16S rRNA tree illustrated the relationship between strains Marseille-Q9181^T^, Marseille-QA0894^T^, Marseille-Q2328^T^ and Marseille-Q9179^T^ and closely related species within the genus Gardnerella (Fig. 3). Comparative analysis of 16S rRNA sequences using the blast tool revealed that strains Marseille-Q9181^T^, Marseille-QA0984^T^, Marseille-Q2328^T^ and Marseille-Q9179^T^ showed sequence homologies of 99.78%, 100%, 100% and 99.80% with strains G. vaginalis ATCC 14018^T^, G. leopoldii UGent 06.41^T^, G. vaginalis NR010 and G. vaginalis 1500E, respectively.
Phylogenetic tree based on 16S rRNA gene sequences showing the evolutionary relationships of G. lacydonensis Marseille-Q9181T, G. bretellae Marseille-QA0894T, G. massiliensis Marseille-Q2328T and G. phocaeensis Marseille-Q9179T with officially described and closely related Gardnerella strains. The tree was deduced using the neighbour-joining method. Evolutionary distances were calculated using the Tamura 3-parameter method. Bootstrap percentages (based on 1,000 replications) are indicated at the nodes.
Functional annotation
The characteristics of the protein-coding gene subsystems detected by RAST in the four species studied were mainly related to subsystems involved in the metabolism of proteins, carbohydrates, amino acids and their derivatives, DNA, nucleosides and nucleotides metabolism, membrane transport, cell wall, capsule, cofactors, vitamins, prosthetic group, pigments and RNA metabolism. However, there were fewer genes encoding cell wall and capsule in strains Marseille-Q2328^T^ and Marseille-Q9179^T^. Genes encoding membrane transport were less prevalent in strains Marseille-QA0894^T^ and Marseille-Q9179^T^. In all four strains, genes encoding secondary metabolism, iron acquisition and metabolism, cell division, the cell cycle, photosynthesis, motility and chemotaxis and nitrogen metabolism were absent. The distribution of subsystem categories and the number of subsystem features for each bacterial strain are shown in Fig. 4. The distribution of COGs, shown in Table 6, is similar for all four strains. The number of genes encoding carbohydrate transport and metabolism was more predominant in strain Marseille-Q9181^T^. The number of genes encoding for mobilome was prevalent to Marseille-QA0894^T^ and Marseille-Q2328^T^. A high number of COG functional genes involved in replication, recombination and repair were found in Marseille-Q2328^T^. No genes encoding chromatin structure and dynamics or nuclear structure were found in any of the four strains.
Genome subsystem coverage, distribution of subsystem categories and number of subsystem features of strains (a) G. lacydonensis Marseille-Q9181T, (b) G. bretellae Marseille-QA0894T, (c) G. massiliensis Marseille-Q2328T and (d) G. phocaeensis Marseille-Q9179T by RAST.
Genome comparison of the four strains
The genomic characteristics of the 4 strains were compared with the genomes of the 14 Gardnerella genomic species available on NCBI (Table S1). dDDH and OrthoANI values are summarized in Table 7. OrthoANI data are also reported in a heat map (Fig. 5). A dDDH value of 72.1% was observed between strain Marseille-Q9181^T^ and strain G. vaginalis 1400E (genomic species 2). Furthermore, an OrthoANI value (96.99%) was observed for strain Marseille-Q9181^T^ and strain G. vaginalis 1400E (genomic species 2). A dDDH value of 72.4% was observed between strain Marseille-QA0894^T^ and strain G. vaginalis 6119V5 (genomic species 9). Furthermore, the highest OrthoANI value (97.16%) was observed for strain Marseille-QA0894^T^ and strain G. vaginalis 6119V5 (genomic species 9). A dDDH value of 75.5% was observed between strain Marseille-Q2328^T^ and strain G. vaginalis NR010 (genomic species 14). Furthermore, the highest OrthoANI value (97.66%) was observed for strain Marseille-Q2328^T^ and strain G. vaginalis NR010 (genomic species 14). The highest dDDH values for strain Marseille-Q9179^T^ were 57.6% with G. swidsinskii GS 9838-1^T^ and 57.3% with G. leopoldii UGent 06.41^T^. OrthoANI values below 95% were observed between strain Marseille-Q9179^T^ and G. swidsinskii GS 9838-1^T^, as well as strain G. leopoldii UGent 06.4^T^.
Heat map showing the average nucleotide identity based on calculated orthology (OrthoANI) of strains G. lacydonensis Marseille-Q9181T, G. bretellae Marseille-QA0894T, G. massiliensis Marseille-Q2328T and G. phocaeensis Marseille-Q9179T relative to the closest Gardnerella species.
Only the strain Marseille QA0894^T^ had one CRISPR sequence and one CRISPR-Cas type IE. The polyketosynthase-related Type III gene cluster was predicted in Marseille-Q9181^T^. Bacteriocin, plasmid, phage and insert sequence were not identified. The gene coding for nanH1 was present in Marseille-Q9181^T^, Marseille-QA0894^T^ and Marseille-Q2328^T^. No gene coding for sialidase was found in Marseille-Q9179^T^. Vaginolysin type 1B was found in Marseille-Q9181^T^ and Marseille-QA0894^T^. Vaginolysin type 1A was found in Marseille-Q2328^T^. Marseille-Q9179^T^ was aligned with vaginolysin type 2 possessing glutamic acid at position −1 and a substitution of valine by alanine at position 6 in the undecapeptide region (Fig. S2). Structural modelling of the candidate proteins vaginolysin and sialidase using PHYRE 2.2 enabled functional confirmation, with a confidence level of 100%.
Discussion
G. vaginalis species is categorized in the phylum Actinomycetota, class Actinomycetes, order Bifidobacteriales, family Bifidobacteriaceae and genus Gardnerella [46]. Until recently, the genus Gardnerella comprised 14 genomic species, 6 of which have got subsequently validly published names [1011]. Gardner and Dukes were the first to demonstrate the presence of G. vaginalis in women with bacterial vaginosis, formerly known as non-specific bacterial vaginitis [47]. Later, several studies showed a significant association between G. vaginalis and bacterial vaginosis [24849]. Bacterial vaginosis is a dysbiosis characterized by a decrease in lactobacilli and the proliferation of several anaerobic bacteria, such as G. vaginalis, Fannyhessea vaginae, Prevotella bivia, Mobiluncus curtisii and many others [5052].
Recent studies revealed that the complex of strains previously attributed to a single species, G. vaginalis, and associated with bacterial vaginosis could represent multiple similar but distinct and sometimes neighbour species [471053]. Several genetically distinct groups have already been described as new species (G. picketti, formerly genomic species 3, G. piotii, formerly genomic species 4, * G. leopoldii*, formerly genomic species 5, G. swidsinskii, formerly genomic species 6, and G. greenwoodii, formerly genomic species 8). Other genomic species are not yet officially described. Here, we continue to dissect former genomic species of G. vaginalis that may represent different species according to modern criteria. We analysed four Gardnerella strains isolated from vagina and blood in Marseille, France, and characterized them.
Strains Marseille-Q9181^T^, Marseille-QA0894^T^ and Marseille-Q9179^T^ have a whitish to greyish morphology almost identical to that of G. vaginalis, G. piotii, G. leopoldii and G. swidsinskii as described by Vaneechoutte et al. [10], which limits the potential for identifying Gardnerella species based solely on the appearance of colonies isolated in culture. Previous studies have used β-galactosidase as a biochemical test to differentiate Gardnerella species [31011]. Our results showed that strains Marseille-QA0894^T^, Marseille-Q2328^T^ and Marseille-Q9179^T^ were negative for β-galactosidase, while strain Marseille-Q9181^T^ exhibited positive β-galactosidase activity. These data support the conclusion that the β-galactosidase test is no longer suitable for identifying Gardnerella species. The strains Marseille-Q9181^T^, Marseille-QA0894^T^, Marseille-Q2328^T^ and Marseille-Q9179^T^ appear as Gram-negative coccobacilli. They can grow at 5% CO_2_ and show no activity for catalase, oxidase and β-glucosidase. These phenotypic characteristics are similar to those observed in other species of the genus Gardnerella [104754].
A recent study updated the MALDI Biotyper database to distinguish between the G. vaginalis/G. piotii and G. leopoldii/G. swidsinskii pairs for routine diagnosis [55]. However, our results showed that even with an improved MALDI Biotyper database, it was not possible to accurately distinguish Gardnerella species.
Phylogenetic analysis based on the comparison of the 16S rRNA gene revealed that strains Marseille-Q9181^T^, Marseille QA0894^T^, Marseille-Q2328^T^ and Marseille-Q9179^T^ were more than 98.5% similar to their closest phylogenetic neighbour. However, this criterion is insufficient to establish a new species in Gardnerella as mentioned in previous studies [101456]. According to the generally accepted criteria for bacterial species delimitation, namely dDDH 70% and ANI 96% [3457], strains Marseille-Q9181^T^, Marseille-QA0894^T^ and Marseille-Q2328^T^ belong to Gardnerella genomic species 2, 9 and 14, respectively (dDDH ≥70% and ANI ≥96%). However, the strain Marseille-Q9179^T^ exhibits dDDH and ANI values below the thresholds established for species delimitation, confirming that this strain is a new species of the genus Gardnerella. Based on phenotypic, phylogenetic and genomic analyses, we describe four species of the genus Gardnerella: G. lacydonensis sp. nov., strain Marseille-Q9181^T^, belongs to the previously reported Gardnerella genomic species 2; G. bretellae sp. nov., strain Marseille-QA0894^T^, belongs to the previously reported Gardnerella genomic species 9 [10]; G. massiliensis sp. nov., strain Marseille-Q2328^T^, belongs to the previously reported Gardnerella genomic species 14 [14]; and G. phocaeensis sp. nov., strain Marseille-Q9179^T^, is a new species of the genus Gardnerella that may also be represented as Gardnerella genomic species 15.
The genome analyses show that the investigated strains have a genome size of 1.491 to 1.716 Mb and a G+C content from 41.1 mol% to 43.4 mol% that is generally comparable to the values described for the genus Gardnerella [58]. However, the strain Marseille-Q2328^T^ is distinguished by a significantly higher G+C content (45.3 mol%) than the officially described strains [1011]. In the genus Gardnerella, variation in G+C content has already been correlated with isolated genomic clades specifically adapted to different ecological niches, suggesting a role in adaptive divergence [58]. It should be noted that the strain with the highest G+C content (Marseille-Q2328^T^) was isolated from blood, in contrast to the others, which were isolated from the vagina. Furthermore, evidence of mobile subsystems and a high number of genes associated with DNA replication, recombination and repair in the strains Marseille-QA0894^T^ and Marseille-Q2328^T^ highlights a diverse and highly adaptable pangenome within these strains, characterized by frequent horizontal gene acquisition, genetic mobility and genome expansion [58].
Sialidase has harmful effects on the vaginal mucosa, facilitating the adhesion of bacteria to the vaginal epithelium and the formation of biofilms [5960]. Previously, three sialidase enzymes (nanH1, nanH2 and nanH3) have been identified in strains of the genus Gardnerella [43]. The nanH1 gene, encoding sialidase A, was detected; however, none of the strains possessed a gene encoding the active enzyme (nanH2 and nanH3) [43]. Thus, these results show that the four strains of Gardnerella do not possess sialidase activity. The four strains also possess the vaginolysin gene, as observed in other Gardnerella species [42]. The species of the genus Gardnerella produces vaginolysin, a toxin that is a cholesterol-dependent cytolysin [61]. According to a recent study, Gardnerella species have five different forms of vaginolysin: type 1A, type 1B, type 1C, type 2 and type 3 [42]. However, the strain Marseille-Q9179^T^ exhibits unique variability in the type 2 undecapeptide region of vaginolysin, which could indicate the existence of a new subclass of vaginolysin.
Description of Gardnerella lacydonensis sp. nov. (formerly known as Gardnerella genomic species 2)
Gardnerella lacydonensis (la.cy.don.en’sis. sis, N.L. fem. adj. lacydonensis, from Lacydon, the name of the ancient port of Marseille, the French city where the strain was first described).
Cells are facultative anaerobic and microaerophilic, non-motile, non-spore-forming and Gram-negative coccobacilli. The cells are 1.39±0.4 µm long and 0.86±0.07 µm wide. Catalase and oxidase activities are negative. Colonies are round, greyish to whitish and opaque on PolyViteX chocolate agar, Columbia CNA agar supplemented with 5% sheep blood and Columbia agar enriched with 5% sheep blood after 48 h of incubation. Growth occurs in a temperature range of 28–37 °C (optimum 37 °C), at a pH of 7.5 to 8.5 (optimum 7.5) and 0.5% NaCl. Using API strips, positive reactions are observed for d-glucose, d-mannitol, lactose, d-sucrose, d-maltose, salicin, aesculin, iron citrate, glycerol, d-cellobiose, d-mannose, d-melezitose, d-sorbitol, starch, alkaline phosphatase, leucine arylamidase, valine arylamidase, acid phosphatase, naphthol-AS-BI-phosphohydrolase, α-glucosidase, ß-galactosidase and d-mannosidase. The predominant fatty acids are C_16 : 0_, C_18 : 1_ ω9c and C_18 : 0_. The genome size is 1.7 Mbp, with a G+C content of 41.4 mol%. The type strain Marseille-Q9181^T^ (=CSUR Q9181^T^=CECT 31121^T^) was isolated from a vaginal sample. The 16S rRNA and genome sequences were deposited in GenBank under accession numbers PQ636763 and JBKQXT000000000, respectively.
Description of Gardnerella bretellae sp. nov. (formerly known as Gardnerella genomic species 9)
Gardnerella bretellae (bre.tel’lae, N.L. gen. n. bretellae, honouring Florence Bretelle for her contribution to the description of vaginal flora).
Cells are strictly anaerobic and microaerophilic, non-motile, non-spore-forming and Gram-negative coccobacilli. Cells are 0.73±0.13 µm long and 0.5±0.06 µm wide. Catalase and oxidase activities are negative. Growth is observed at 37–42 °C (optimum 37 °C), pH 6.5–8.5 (optimum 7.5) and 0.5% NaCl. Bacterial colonies are round, greyish to whitish and opaque on PolyViteX chocolate agar, Columbia CNA agar supplemented with 5% sheep blood and Columbia agar enriched with 5% sheep blood after 48 h of incubation at 37 °C. Using API strips, positive reactions are observed for d-glucose, d-mannitol, lactose, d-sucrose, d-maltose, salicin, aesculin, iron citrate, glycerol, d-cellobiose, d-mannose, d-melezitose and d-sorbitol, starch, alkaline phosphatase, leucine arylamidase, valine arylamidase, acid phosphatase, naphthol-AS-BI-phosphohydrolase and α-glucosidase. The predominant fatty acids are C_16 : 0_, C_18 : 1_ ω9c and C_18 : 0_. The genome size is 1.5 Mbp, with a G+C content of 43.2 mol%. The type strain Marseille-QA0894^T^ (=CSUR QA0894^T^=CECT 31122^T^) was isolated from a vaginal sample. The 16S rRNA and genome sequences were deposited in GenBank under accession numbers PQ636764 and JBKQXU000000000, respectively.
Description of Gardnerella massiliensis sp. nov. (formerly known as Gardnerella genomic species 14)
Gardnerella massiliensis (mas.si.li.en’sis. N.L. fem. adj. massiliensis, from Massilia, the Latin name of Marseille in France, where the bacterium was first described).
Cells are facultative anaerobic and microaerophilic, non-motile, non-spore-forming and Gram-negative coccobacilli. Cells are 3±0.64 µm long and 1.5±0.09 µm wide. Catalase and oxidase activities are negative. Growth occurs in a temperature range of 28–42 °C (optimum 37 °C) at a pH of 6.5 to 8.5 (optimum 7.5) and 0.5% NaCl. Bacterial colonies are small, translucent, almost transparent and circular on Columbia agar enriched with 5% sheep’s blood and Columbia CNA agar supplemented with 5% sheep’s blood, after 48 h of incubation at 37 °C. Using API strips, positive reactions are observed for d-glucose, d-mannitol, lactose, d-sucrose, d-maltose, salicin, aesculin, iron citrate, glycerol, d-cellobiose, d-mannose, d-melezitose, d-sorbitol, starch, alkaline phosphatase, leucine arylamidase, valine arylamidase, acid phosphatase, naphthol-AS-BI-phosphohydrolase and α-glucosidase. The most abundant fatty acids are C_16 : 0_, C_18 : 1_ ω9c and C_18 : 0_. The genome size is 1.7 Mbp, with a G+C content of 45.3 mol%. The type strain Marseille-Q2328^T^ (=CSUR Q2328^T^=CECT 30239^T^) was isolated from blood. The 16S rRNA and genome sequences were deposited in GenBank under accession numbers MT822479 and CALNWW000000000, respectively.
Description of Gardnerella phocaeensis sp. nov.
Gardnerella phocaeensis (pho.cae.en’sis, N.L. fem. adj. phocaeensis, referring to Phocaea, the name of the Ionian Greek city where the founders of Marseille came from. The strain was isolated in Marseille).
Cells are facultative anaerobic and microaerophilic, non-motile, non-spore-forming and Gram-negative coccobacilli. Cells are 1±0.05 µm long and 0.54±0.04 µm wide. Catalase and oxidase activities are negative. Colonies are greyish to whitish in appearance and circular on Columbia agar enriched with 5% sheep’s blood, PolyViteX chocolate agar and Columbia CNA agar enriched with 5% sheep’s blood, after 48 h of incubation at 37 °C.
Growth occurs in a temperature range of 28–42 °C (optimum 37 °C), at a pH of 7.5 to 8.5 (optimum 7.5) and 0.5% NaCl. Using API strips, positive reactions are observed for d-glucose, d-mannitol, lactose, d-sucrose, d-maltose, salicin, aesculin, iron citrate, glycerol, d-cellobiose, d-mannose, d-melezitose, d-sorbitol, starch, l-rhamnose, d-trehalose, alkaline phosphatase, leucine arylamidase, valine arylamidase, acid phosphatase, naphthol-AS-BI-phosphohydrolase, α-glucosidase, esterase (C4), esterase-lipase (C8), lipase (C14) and ß-glucuronidase. The most abundant fatty acids are C_16 : 0_, C_18 : 1_ ω9c, C_18 : 0_ and C_14 : 0_. The genome size is 1.5 Mbp, with a G+C content of 42.3 mol%. The type strain Marseille-Q9179^T^ (=CSUR Q9179^T^=CECT 31120^T^) was isolated from a vaginal sample. The 16S rRNA and genome sequences were deposited in GenBank under accession numbers PQ636765 and JBKQXS000000000, respectively.
Supplementary material
10.1099/ijsem.0.007028Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Schwebke JR Muzny CA Josey WE Role of Gardnerella vaginalis in the pathogenesis of bacterial vaginosis: a conceptual model J Infect Dis 201421033834310.1093/infdis/jiu 08924511102 · doi ↗ · pubmed ↗
- 2Morrill S Gilbert NM Lewis AL Gardnerella vaginalis as a cause of bacterial vaginosis: appraisal of the evidence from in vivo models Front Cell Infect Microbiol 20201016810.3389/fcimb.2020.0016832391287 PMC 7193744 · doi ↗ · pubmed ↗
- 3Piot P Van Dyck E Peeters M Hale J Totten PA et al Biotypes of Gardnerella vaginalis J Clin Microbiol 19842067767910.1128/jcm.20.4.677-679.19846333436 PMC 271409 · doi ↗ · pubmed ↗
- 4Janulaitiene M Paliulyte V Grinceviciene S Zakareviciene J Vladisauskiene A et al Prevalence and distribution of Gardnerella vaginalis subgroups in women with and without bacterial vaginosis BMC Infect Dis 20171739410.1186/s 12879-017-2501-y 28583109 PMC 5460423 · doi ↗ · pubmed ↗
- 5Ingianni A Petruzzelli S Morandotti G Pompei R Genotypic differentiation of Gardnerella vaginalis by amplified ribosomal DNA restriction analysis (ARDRA)FEMS Immunol Med Microbiol 199718616610.1111/j.1574-695X.1997.tb 01028.x 9215588 · doi ↗ · pubmed ↗
- 6Balashov SV Mordechai E Adelson ME Gygax SE Identification, quantification and subtyping of Gardnerella vaginalis in noncultured clinical vaginal samples by quantitative PCRJ Med Microbiol 20146316217510.1099/jmm.0.066407-024200640 · doi ↗ · pubmed ↗
- 7Munch MM Strenk SM Srinivasan S Fiedler TL Proll S et al Gardnerella species and their association with bacterial vaginosis J Infect Dis 2024230 e 171e 18110.1093/infdis/jiae 02639052736 PMC 11272073 · doi ↗ · pubmed ↗
- 8Schellenberg JJ Paramel Jayaprakash T Withana Gamage N Patterson MH Vaneechoutte M et al Gardnerella vaginalis subgroups defined by cpn 60 sequencing and sialidase activity in isolates from Canada, Belgium and Kenya P Lo S One 201611 e 014651010.1371/journal.pone.014651026751374 PMC 4709144 · doi ↗ · pubmed ↗