Phenotypic and genomic characterization of Roseomonas mucosa, an opportunistic pathogen with discrepancies among antimicrobial susceptibility testing methods
Camille Cotet, Sebastien Galopin, Anne-Emeline Creach, Marwan Tenouri, Charlotte Le Pont, Nathalie Laquay, Jean-Christophe Giard, Simon Le Hello, François Gravey

TL;DR
This study examines antibiotic resistance patterns and genetic factors in Roseomonas mucosa, a bacterium that causes infections in immunocompromised patients.
Contribution
The study identifies a novel regulatory mechanism of beta-lactamase expression in R. mucosa that explains discrepancies in susceptibility testing.
Findings
High susceptibility to aminoglycosides, fluoroquinolones, and carbapenems was observed in R. mucosa strains.
Discrepancies in susceptibility testing methods were linked to a class A beta-lactamase regulated by an LysR-type regulator.
Abstract
Roseomonas mucosa is an opportunistic bacterium found in clinical and environmental samples that primarily affects immunocompromised patients. Treatment is challenging owing to the lack of standardized susceptibility testing methods, breakpoints, and variable antimicrobial resistance profiles published. This study evaluated different antimicrobial susceptibility testing approaches and searched for new insights into resistance mechanisms, especially against third-generation cephalosporins. Antimicrobial susceptibility profiles of a panel of 17 R. mucosa strains were analyzed using disk diffusion method (DDM), broth microdilution (BMD) method, and MIC gradient strips. Discrepancies between susceptibility methods were further explored using beta-lactamase inhibitors, whole-genome sequencing, and transcriptomic analyses. Antimicrobial susceptibility testing revealed high susceptibility to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
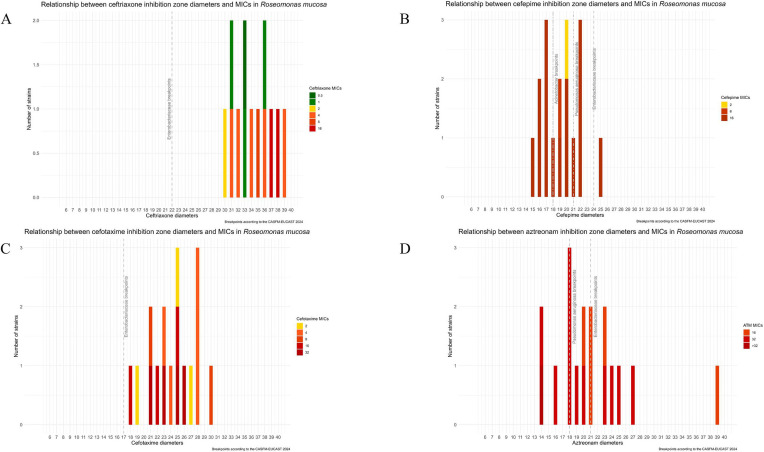 Fig 1
Fig 1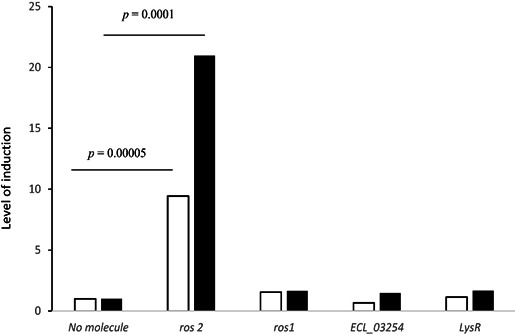 Fig 2
Fig 2| Antibiotic | MIC (mg/L) | Diameter (mm) | |||||
|---|---|---|---|---|---|---|---|
| MIC50 | MIC90 | % S | Minimum | Maximum | Median | % S | |
| Piperacillin-tazobactam | 1,024 | 1,024 | 0 | 6 | 7 | 6 | 0 |
| Temocillin | >32 | >32 | 0 | 6 | 7 | 6 | NA |
| Ceftriaxone | 4 | 16 | 40 | 30 | 40 | 36 | NA |
| Cefotaxime | 8 | 32 | 20 | 18 | 30 | 25 | NA |
| Cefepime | 16 | 16 | 10 | 15 | 25 | 19 | 100 |
| Ceftazidime | 32 | 32 | 0 | 6 | 7 | 6 | 0 |
| Ceftazidime-avibactam | >32 | >32 | 0 | NA | NA | NA | NA |
| Ceftolozane-tazobactam | >32 | >32 | 0 | NA | NA | NA | NA |
| Ertapenem | 0.125 | 0.25 | 100 | 36 | 40 | 38 | NA |
| Imipenem | ≤1 | ≤1 | 100 | 37 | 40 | 39 | 100 |
| Imipenem-relebactam | 0.5 | 1 | 100 | NA | NA | NA | NA |
| Meropenem | 0.5 | 0.5 | 100 | 28 | 40 | 40 | 100 |
| Meropenem-vaborbactam | 0.25 | 0.25 | 100 | NA | NA | NA | NA |
| Aztreonam | 32 | 32 | 0 | 14 | 39 | 20 | NA |
| Cefiderocol | 16 | 16 | 0 | NA | NA | NA | NA |
| Amikacin | ≤2 | ≤2 | NA | 22 | 40 | 40 | 100 |
| Gentamicin | 0.125 | 0.125 | 100 | 35 | 40 | 40 | 100 |
| Tobramycin | ≤0.5 | ≤0.5 | 100 | 34 | 40 | 40 | 100 |
| Ciprofloxacin | 0.25 | 0.5 | 1 | 21 | 40 | 40 | 100 |
| Tigecycline | ≤0.5 | ≤0.5 | 100 | 28 | 40 | 35 | NA |
| Cotrimoxazole | ≤1 | ≤1 | 100 | 20 | 40 | 39 | 100 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Infections and bacterial resistance · Aquaculture disease management and microbiota
INTRODUCTION
Described for the first time in 1993 (1), bacteria that belong to the Roseomonas genus are now recognized as emerging opportunistic pathogens. This gram-negative coccobacillus, which is aerobic, slow growing, and pink pigmented, has been isolated from both clinical and environmental samples (2, 3). The reservoirs include surfaces, water, soil, plants, and human skin.
Roseomonas mucosa is the most common species isolated from clinical samples, mainly among immunocompromised patients with comorbidities such as malignant tumors, diabetes, systemic lupus erythematosus, and acquired immunodeficiency syndrome (4). Clinical infections may vary and include bacteremia, respiratory and skin infections, and peritonitis (5, 6). Invasive infections in immunocompetent patients, including meningitis (7) or dental root canal infection (8), have also been described.
Antimicrobial strategies for the treatment of these infections can be challenging because studies have focused mainly on case reports (9). There is no standard laboratory method for drug susceptibility testing specific to this genera (EUCAST-CASFM –-Comité de l’Antibiogramme de la Société Française de Microbiologie 2024; CLSI). This bacterium is typically described as susceptible to quinolones, aminoglycosides, and carbapenems but is frequently resistant to beta-lactams (ampicillin, piperacillin, and cephalosporins) (10). Besides, antimicrobial susceptibility varies among species; R. mucosa has shown higher antibiotic resistance rates (2).
In April 2022, a 63-year-old woman was hospitalized in our teaching hospital for the excision of a meningioma of the right ventricular junction. One week later, an external ventricular bypass was introduced due to intracranial pressure. After 2 weeks, she developed a persistent fever despite antibiotic treatment. Cerebrospinal fluid (CSF) analysis identified R. mucosa, leading to the diagnosis of cerebral ventriculitis. Empirical antimicrobial therapy was initiated with the combination of meropenem with intrathecal injections of amikacin. First-line antibiotics were switched for cefepime, which was categorized as “susceptible” regarding the diameters and EUCAST-CA-SFM guidelines (https://www.sfm-microbiologie.org/). Ten days later, the electroencephalogram revealed diffuse brain damage attributed to toxic encephalopathy due to the cefepime iatrogenic effect, confirmed by CSF analysis, which revealed an overdose of the antibiotic. Following a collegiate decision, treatment was changed to cotrimoxazole and ciprofloxacin for 3 weeks, leading to recovery.
In this study, we investigated the antimicrobial susceptibility profiles of a cohort of 17 R. mucosa strains alongside an in-depth genomic analysis. We combined phenotypic testing, whole-genome sequencing, and transcriptomics to understand discrepancies between antimicrobial susceptibility methods, leading to the description of three beta-lactamases in the genomes of R. mucosa species.
MATERIALS AND METHODS
Strains and growth conditions
The 17 strains were isolated between 1997 and 2022 in the teaching hospital of Caen Normandy, France. They were distributed among 14 R. mucosa samples (including the strain of the clinical case, id: 2733), one Roseomonas giliardii sample, and two Roseomonas spp. Species were identified via MALDI-TOF Biotyper sirius Brucker (Billerica, MA, USA). Most of the strains were isolated from human samples (n=13), notably from blood cultures (n=5), meconium (n=5), cerebrospinal fluid (n=2), and finger swabs (n=1). The four environmental isolates were collected from surfaces or gloves by the hospital hygienist department (Table S1).
Optimal growth conditions were assessed by incubating cultures at 35°C for 24 and 48 h under aerobic and anaerobic conditions using Mueller–Hinton medium or Muller–Hinton fastidious agar.
Antimicrobial susceptibility testing and determination of the minimum inhibitory concentrations
Twenty-eight antibiotics were tested via the disk diffusion method (DDM). Inhibition diameters were interpreted using EUCAST-CASFM 2024 breakpoints for Acinetobacter spp. Minimum inhibitory concentrations (MICs) of 24 molecules were determined via the broth microdilution (BMD) method with FRAM2GN and EUMDROXF Sensititre plates (Thermo Fisher Scientific, Waltham, MA, USA) or manually. For 16 molecules, the susceptibility profiles were determined via both DDM and BMD methods (Table S2). MICs of cefiderocol were determined using UMIC test kits (Bruker, Germany) according to the manufacturer’s instructions. Considering the absence of breakpoints for Roseomonas species, in both EUCAST and CA-SFM referential, MICs were interpreted according to breakpoints available in the “nonrelated species PK/PD breakpoints” section of the EUCAST-CASFM 2024 guidance document. In case of discrepancies between methods only, results obtained with BMD were considered.
MIC gradient strips were used to compare the MICs of cefotaxime and ceftriaxone with those obtained with BMD approaches. MIC gradient MICs from two manufacturers, Biomerieux (Marcy l’étoile, France) and Liofilchem (Roseto degli Abruzzi, Italy), were measured according to their recommendations.
Plates were incubated at 35°C for 48 h for all the antimicrobial susceptibility approaches.
The associations between MICs and diameters were visualized using R version 4.4.2 on RStudio software version 4.2.1 and ggplot2 version 3.5.1 (11).
Genomic analyses
Genomes of the seventeen strains were sequenced using the NextSeq 500 platform, generating 150 pb paired-end reads (Illumina Technology, San Diego, CA, USA) after DNA extraction via the Magna Pure system (Roche, Bate, Suisse) and library preparation by the Nextera Kit according to the manufacturer’s instructions (Illumina, San Diego, CA, USA) at the “plateforme de microbiologie mutualisée” (p2m, Institut Pasteur, Paris, France). Quality control of the reads was performed with FastQC https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ and MultiQC (12). De novo assembly was carried out via SPAdes v3.12 (13). The quality of the genomes was assessed using QUAST (14). Species determination was performed using the rMLST tool (15). Genome annotation was performed with Prokka (16). Antimicrobial resistance genes were looked for in both Resfinder (17) and CARD (18) databases; all questioned in December 2024.
Sixty-four additional genomes of Roseomonas spp. were downloaded from NCBI to realize an average nucleotide identity (ANI) approach between the genomes from this study and genomes available in RefSeq. Details are available in Table S3. ANI was performed using the mash dist algorithm (19); the distance matrix obtained was converted into a neighbor-joining tree using the Phylogeny Inference Package (20) and visualized using iTOL version 7 (21).
Sequences of the three beta-lactamases as well as the lysR regulator were in silico translated to assess the diversity of these proteins within the genomes collection.
Exploration of the effects of beta-lactamase inhibitors on the MICs of third-generation cephalosporins
MICs of cefotaxime and ceftriaxone were assessed by adding clavulanic acid in the Mueller-Hinton (MH) medium using clavulanic acid/amoxicillin MIC gradient strips Biomerieux (Marcy l’étoile, France), or via a specific MH plus cloxacillin (250 mg/L) Biomerieux (Marcy l’étoile, France). MICs were measured using MIC gradient strips; plates were incubated aerobically at 35°C for 48 h.
Transcriptomic response of R. mucosa to cefotaxime exposure
Strain 2733 was grown on MH plates with ceftriaxone and cefotaxime MIC gradient strips. After 48 h at 37°C, cells near the strips (with antibiotics) and those from the plate periphery (without antibiotics) were harvested. Total RNA was extracted using the Maxwell RSC miRNA Tissue Kit (Promega, Madison, WI, USA) and quantified via NanoDrop (Thermo Fisher Scientific, Waltham, MA, USA). cDNA synthesis (0.1 μg) and RT-qPCR were performed using the Platinum SYBR Green One-Step qRT-PCR Kit (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. The expression of beta-lactamase encoding genes (ros1, ros2, and ecl_03254) as well as the lysR was calculated by comparing transcription levels with and without antibiotics, using gyrA as an internal control. Experiments were repeated at least three times. Ratios were compared using the ΔΔCt method (22); statistical significance was assessed using Student’s t-test (P < 0.05). Sequences of the primers used are available in Table S4. Ros2 gene, for which its expression was associated with ceftriaxone and cefotaxime exposure, was cloned with its promoter and expressed in Escherichia coli to further explore its contribution to the observed phenotype.
Construction of multicopy plasmid containing ros2 beta-lactamase
The ros2 gene from R. mucosa strain 2733, with or without its own promoters, was amplified by PCR using primers listed in Table S5. Each PCR product was then cloned into the low-copy number overexpression plasmid pBAD202 Directional TOPO (Invitrogen, Carlsbad, CA, USA) and transformed into E. coli Top10 recipient cells (Invitrogen) according to the manufacturer’s instructions. E. coli cells carrying pBAD202 recombinants containing correctly oriented fragments were selected on LB plates with 50 mg/L of kanamycin. Inhibition diameters of amoxicillin, amoxicillin + clavulanic acid, cefotaxime, and ceftriaxone were measured using E. coli cells carrying pBAD202 with and without ros-2 cloned with its promoter.
RESULTS
Species determination
Based on the mass spectrometry results, 14 strains belonged to the R. mucosa species, one to R. gilardii species (1793), and two (3994, 11153) were only identified at the genus level. Genomic analyses revealed that all the strains belonged to the R. mucosa species (Table S1). ANI approaches were in accordance with the rMLST taxonomic assignation, Fig. S1.
Antimicrobial profile determination
The optimal growth condition was an aerobic atmosphere on MH agar and was used during all the following experimentations.
Considering the DDM results, several antibiotics were active including aminoglycosides: amikacin, gentamicin, and tobramycin (percentage of sensitivity [PS] 100%); fluoroquinolones: ciprofloxacin and levofloxacin (PS 100%), carbapenems imipenem and meropenem (PS 100%); ticarcillin-clavulanic acid (PS 100%), cefepime (PS:100%), and cotrimoxazole (SP 100%; Table S6). In contrast, all the strains were resistant to piperacillin-tazobactam and ceftazidime (Table S6). BMD confirmed the efficacy of aminoglycosides (MIC_50/90_ of gentamicin 0.125 mg/L, MIC_50/90_ of tobramycin ≤0.5 mg/L; and MIC_50/90_ of amikacin ≤2 mg/L) and cotrimoxazole (MIC_50/90_ ≤ 1 mg/L; Table 1; Table S7). Notably, MICs of colistin were high (MIC_50/90_ 16 mg/L), whereas the MICs of eravacycline were low (MIC_50/90_ 0.06 mg/L; Table S7). For both antibiotics, no clinical interpretations were made due to the absence of PK/PD breakpoints in the CA-SFM 2024 reference.
Among the beta-lactams, the efficacies of new beta-lactam–beta-lactamase inhibitor combinations as well as new molecules were variable. All the strains were resistant to ceftazidime-avibactam (MIC_50/90_ > 32 mg/L) or ceftolozan-tazobactam (MIC_50/90_ > 32 mg/L; Table 1; Table S7). The combinations of imipenem-relabactam and meropenem-vaborbactam were effective (MIC_50/90_ of 1 mg/L and MIC_50/90_ of 0.25/1 mg/L, respectively; Table 1; Table S7). In contrast, temocillin, ceftazidime, piperacillin-tazobactam, and cefiderocol were ineffective (MIC_50/90_ > 32 mg/L, MIC_50/90_ > 32 mg/L, MIC_50/90_ 1,024 mg/L, and MIC_50/90_ 16mg/L, respectively; Table 1; Table S7).
Whereas all the strains were categorized as sensitive to cefepime according to the DDM, the susceptibility rate dropped to 10% regarding the BMD results (Table 1). Furthermore, the sensitivity rates of ceftriaxone and cefotaxime were 40% and 20%, respectively (Table 1; Table S8). These low susceptibility rates were unexpected considering the inhibition diameters observed for both molecules: 36 mm for ceftriaxone and 25 mm for cefotaxime (Fig. S2). The MICs of ceftriaxone and cefotaxime were confirmed via MIC gradient strips from two different manufacturers. For cefotaxime, the MIC_50/90_ were higher with the MIC gradient strips rather than with the BMD test (>32 mg/L; Table S8). For ceftriaxone, the MICs obtained with the MIC gradient were slightly lower than those obtained with BMD (MIC_50/90_ 2/4 mg/L with Biomerieux and MIC_50/90_ 4/8 mg/L with Liofilchem; Table S8). Notably, colonies that were grown at relatively high MICs were tested again by DDM and presented the same original diameter, suggesting that the strain adapted to elevated concentrations of cephalosporins rather than being resistant mutants. According to these results and for the 16 molecules tested with both DDM and BMD, the distribution of diameters and MICs was studied (Fig. S3). For ceftriaxone (Fig. 1A), cefepime (Fig. 1B), cefotaxime (Fig. 1C), and aztreonam (Fig. 1D), the diameter measured was not predictive of the MICs.
Distribution between DDM and BMD for the R. mucosa cohort according to the CASFM-EUCAST 2024 breakpoints. (A) Ceftriaxone. (B) Cefepime. (C) Cefotaxime. (D) Aztreonam.
Identification of two putative class A and one class C beta-lactamase
No gene encoding for a beta-lactamase was found in the genomes according to ResFinder and Card databases. More detailed explorations of the genome annotation of the 17 strains revealed the presence of three proteins that harbored a putative beta-lactamase motif (Table S9). The first gene, PRJNA1137846: AB3X21_00595, encoded for a serine hydrolase with beta-lactamase activity and will be hereafter named ROS-1. The corresponding gene was located between a metalloendopeptidase and a voltage-gated chloride channel-encoding gene. The best match in the card database of this protein was OKP-C-1 beta-lactamase, with an identity rate of 30% and coverage of 44%, followed by several variants of SHV beta-lactamase from Klebsiella pneumoniae, namely, SHV-156, SHV-157, and SHV-62, with identity rates between 30% and 31% and coverage between 49% and 50%. The second protein, PRJNA1137846: AB3X21_17530, was annotated as a class A beta-lactamase and will hereafter be named ROS-2. The corresponding gene was located between a gluconate 2-dehydrogenase subunit 3 family and a LysR family transcriptional regulator. This putative beta-lactamase shares approximately 50% identity with several class A beta-lactamases, including the PEN beta-lactamase recovered from Burkholderia multivorans strains, CARB from Acinetobacter baumannii, Psychobacter maritimus, or Oligella urethralis, and BOR-1 from Bordetella bronchiseptica. The protein sequence of the LysR regulator (PRJNA1137846: AB3X21_17535) shares more than 80% identity with the AmpR/LysR transcriptional activator identified in Pseudoroseomonas coralli, Acetobacteraceae bacterium, and Aliidongia dinghuensis. A class C beta-lactamase-encoding gene was also found. This gene was next close to the ECL_03254 gene (between 99% and 100% identity and 100% coverage), described among Enterobacter cloacae complex strains.
To experimentally verify the beta-lactamase activity of the rso2 gene product, we cloned this locus into a strain of E. coli. This heterologous complemented strain proved to be more susceptible to amoxicillin than the wild-type strain when clavulanic acid was added, showing that Ros2 belonged to a class A enzyme (Table S10).
The sequence of the LysR transcriptional factor was similar among all the strains. Two different sequences of the protein ECL_03254 were found, distant by only one amino acid substitution p.Ala85Thr (Table S9). Three sequences of ROS-2 were found with a maximum of three amino acid substitutions p.Ser42Gly, p.Val252Ala, and p.Gly282Ser (Table S9). On the opposite, ROS-1 was very variable with a total of 10 different sequences including six proteins with a deletion of the C-terminal extremity (Table S9). Considering the MICs measured and the integrity of the proteins, it seems that ROS-1 did not have an impact on the phenotype observed as the MICs were uniformly distributed regardless of the integrity of this protein (Table S9). Those three beta-lactamases were found in all the 41 genomes of R. mucosa (NCBI + this study; Table S11). The sequence of ROS-2 was conserved, and all the genes presented the same length with an identity rate from 99.2% to 100%. The sequence of ROS-1 was less conserved, with gene length between 887 and 1,266 nucleotides and identity percentages from 97.5% to 100%. Considering the ECL_03254_betalactamase, all the genes presented the same length with an identity rate from 99.5% to 100%.
Exploration of discrepancies between susceptibility testing methods
Considering the presence of both a Class-C and two Class-A beta-lactamases, the efficiency of beta-lactamase inhibitors on the MICs was assessed. Cefotaxime MICs were measured on an MH agar associated with 250 mg/L cloxacillin to counteract the effect of a putative beta-lactamase with a cephalosporinase activity, but MICs did not change (data not shown). The combination of acid clavulanic/amoxicillin plus cefotaxime presented an MIC of 0.75 mg/L versus greater than 32 mg/L without clavulanic acid, which indicated a reduction of more than five dilutions of the MICs of cefotaxime alone (Fig. S4). The MICs of ceftriaxone decreased from 2 mg/L to 1 mg/L after the addition of clavulanic acid/amoxicillin. These results suggest the involvement of a type A-beta lactamase in the observed phenotype. This fact was also suspected based on the inhibition diameters of amoxicillin-clavulanic acid and ticarcillin-clavulanic acid, which were all larger than those of amoxicillin and ticarcillin alone (Table S6).
Transcriptional level of beta-lactamases in the presence of cefotaxime or ceftriaxone
As shown in Fig. 2, the expression of the ros2 gene increased 9- and 20-fold in the presence of the third-generation cephalosporins ceftriaxone and cefotaxime, respectively. At the same time, the transcription of the two other beta-lactamases (ros1 and ECL_03254 like) as well as the lysR genes was not affected by the presence of these molecules.
Levels of induction of ros2, ros1, ECL_03254, and lysR gene expression when cells were cultured in the presence of ceftriaxone (white bars) and cefotaxime (black bars; see Materials and Methods section) compared with the expression level in the absence of antibiotics (normalized to 1).
Altogether, (i) the fact that truncated ROS-1 had no effect on the MICs measured, (ii) the absence of cloxacillin effect on the MICs, and (iii) the transcriptomics results demonstrated that ROS-2 was responsible for the phenotype observed.
DISCUSSION
Little is known about the appropriate antimicrobial therapy and which molecule should be tested in microbiology laboratories to respond to R. mucosa infections, whereas this opportunistic pathogen has been associated with various community- and hospital-acquired infections (23).
Our results mostly agree with those of Han et al., who analyzed 36 strains (and 2 ATCC) of Roseomonas species (2). Where all the strains were susceptible to amikacin and ciprofloxacin, most of them were also susceptible to ticarcillin-clavulanate (31/38), whereas susceptibility rates to cotrimoxazole were lower (29% [11/38]). Han et al. also warned against the poor efficacy of third-generation cephalosporins (ceftriaxone S = 18/38; ceftazidime S = 0/38). Moreover, the results from Nariyama and coworkers are consistent with our findings, except for the use of cotrimoxazole, which was inefficient toward the strain causing peritonitis infection (5). The overall susceptibility patterns of several previously reported cases are in agreement with our findings (4, 6, 24, 25). Nevertheless, none of the published studies address the discrepancies regarding cefotaxime and ceftriaxone DDM/BDM results despite their common use in antimicrobial therapy strategies.
Despite the absence of results within the resfinder (26) and card databases (18), in silico analyses revealed the presence of three putative beta-lactamases in the R. mucosa genomes. Among them, ROS-2 was responsible for the discrepancies between inhibition diameters and MICs for cefotaxime and ceftriaxone. Interestingly, a class D beta-lactamase was found among Roseomonas fluvialis genomes (27). This oxacillinase was not found in any genomes of R. mucosa, from this study either from NCBI.
Our study highlighted the necessity of considering MICs and not using the DDM to determine the antimicrobial susceptibility of this bacterium, especially for cephalosporins. The discrepancy between three susceptibility methods, namely, the DDM, BMD, and MIC gradient strips, is a crucial issue in clinical microbiology. DDM and MIC gradients are commonly used to assess the antimicrobial susceptibility profiles of bacteria. In this context, they yielded unreliable results, as demonstrated in this study, posing challenges for the treatment of infections. Discrepancies between DDM and the MIC gradient (both based on an antibiotic gradient in the agar plate) could be correlated with the concentration required for the new ROS-2 beta-lactamase induction. Moreover, the impact of the breakpoints used to interpret MICs is crucial, mainly for cefepime; with a MIC50/90 at 16mg/L, all strains were categorized as resistant considering PK/PD values, but could have been considered “sensible” with the Acinetobacter spp. breakpoints.
With the incorporation of ros-2 into the pBAD plasmid, the E. coli TOP-10 strain had a phenotypic class A penicillinase phenotype. Unfortunately, no modification of the MICs of third-generation cephalosporins was observed. This observation could be explained by the fact that the transcriptional regulator lys-R was not included in the plasmid, whereas this protein is crucial for ros-2 hyper-expression, as has been demonstrated by the transcriptomic analyses.
Conclusion
Significant discrepancies between susceptibility methods performed by DDM and those based on the determination of MICs were demonstrated for R. mucosa. As a consequence, in line with the official guidelines CA-SFM and EUCAST, MICs must be used instead of diffusion methods to interpret the susceptibility of strains to avoid treatment failure.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rihs JD, Brenner DJ, Weaver RE, Steigerwalt AG, Hollis DG, Yu VL. 1993. Roseomonas, a new genus associated with bacteremia and other human infections. J Clin Microbiol 31:3275–3283. doi:10.1128/jcm.31.12.3275-3283.19938308122 PMC 266400 · doi ↗ · pubmed ↗
- 2Han XY, Pham AS, Tarrand JJ, Rolston KV, Helsel LO, Levett PN. 2003. Bacteriologic characterization of 36 strains of Roseomonas species and proposal of Roseomonas mucosa sp nov and Roseomonas gilardii subsp rosea subsp nov. Am J Clin Pathol 120:256–264. doi:10.1309/731V-VGVC-KK 35-1Y 4J 12931556 · doi ↗ · pubmed ↗
- 3Romano-Bertrand S, Bourdier A, Aujoulat F, Michon A-L, Masnou A, Parer S, Marchandin H, Jumas-Bilak E. 2016. Skin microbiota is the main reservoir of Roseomonas mucosa, an emerging opportunistic pathogen so far assumed to be environmental. Clin Microbiol Infect 22:737. doi:10.1016/j.cmi.2016.05.02427269884 · doi ↗ · pubmed ↗
- 4Shao S, Guo X, Guo P, Cui Y, Chen Y. 2019. Roseomonas mucosa infective endocarditis in patient with systemic lupus erythematosus: case report and review of literature. BMC Infect Dis 19:140. doi:10.1186/s 12879-019-3774-030755159 PMC 6373168 · doi ↗ · pubmed ↗
- 5Nariyama T, Ito Y, Fujita K, Ito T, Terawaki H. 2024. Successfully treated Roseomonas mucosa-induced peritonitis diagnosed by mass spectrometry. Intern Med 63:2311–2315. doi:10.2169/internalmedicine.2998-2338220190 PMC 11414375 · doi ↗ · pubmed ↗
- 6Beucler N, Meyer M, Choucha A, Seng P, Dufour H. 2020. Peritonitis caused by Roseomonas mucosa after ventriculoperitoneal shunt revision: a case report. Acta Neurochir (Wien) 162:2459–2462. doi:10.1007/s 00701-020-04449-932535796 · doi ↗ · pubmed ↗
- 7Waris RS, Ballard M, Hong D, Seddik TB. 2021. Meningitis due to Roseomonas in an immunocompetent adolescent. Access Microbiol 3:000213. doi:10.1099/acmi.0.00021334151165 PMC 8209696 · doi ↗ · pubmed ↗
- 8Diesendorf N, Köhler S, Geißdörfer W, Grobecker-Karl T, Karl M, Burkovski A. 2017. Characterisation of Roseomonas mucosa isolated from the root canal of an infected tooth. BMC Res Notes 10:212. doi:10.1186/s 13104-017-2538-428615078 PMC 5471672 · doi ↗ · pubmed ↗