In vivo selection of CMY-219 conferring resistance to ceftazidime-avibactam in an OXA-484-producing E. coli ST410
Agnès B. Jousset, Delphine Girlich, Saoussen Oueslati, Aurélien Birer, Anne Delaval, Caroline Guyot, Ines Rezzoug, Cécile Emeraud, Thierry Naas, Rémy A. Bonnin, Laurent Dortet

TL;DR
This study shows how a variant of the CMY-42 enzyme can cause resistance to a key antibiotic in a specific strain of E. coli.
Contribution
The study identifies CMY-219 as a novel variant linked to ceftazidime-avibactam resistance in an OXA-484-producing E. coli ST410.
Findings
Ceftazidime-avibactam resistance was conferred by the CMY-219 variant in an OXA-484-producing E. coli ST410.
ST410 and related clones are at risk of selecting CMY variants under ceftazidime-avibactam exposure.
These clones commonly carry CMY-42 and modified PBP3, increasing their potential for carbapenemase acquisition.
Abstract
Resistance to ceftazidime-avibactam (CAZ-AVI) is a growing problem. This study describes the selection of CMY-219, a CMY-42 variant (G156D), conferring resistance to CAZ-AVI in an OXA-484-producing Escherichia coli ST410 after treatment. It raises concern about the risk of selection of CMY variants under CAZ-AVI exposure in ST410 and related clones, which commonly carry CMY-42, are prone to carbapenemase acquisition, and harbor modified PBP3.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
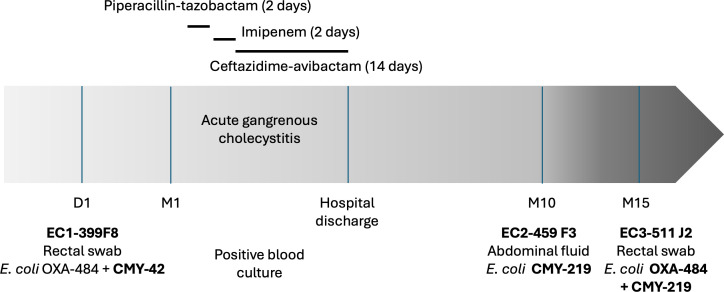 Fig 1
Fig 1| Clinical isolates | Transformants | |||||
|---|---|---|---|---|---|---|
| Amoxicillin | >256 | >256 | >256 | >256 | >256 | 32 |
| Amoxicillin-clavulanate | >256 | >256 | >256 | >256 | >256 | 24 |
| Piperacillin | >256 | >256 | >256 | 64 | 128 | 4 |
| Piperacillin-tazobactam | >256 | 32 | >256 | 4 | 4 | 1.5 |
| Cefoxitin | >256 | >256 | >256 | >256 | >256 | 24 |
| Cefotaxime | >32 | >32 | >32 | 8 | >32 | 2 |
| Ceftazidime | >256 | >256 | >256 | 64 | >256 | 64 |
| Ceftazidime-avibactam | 1 | 16 | 16 | 0.25 | 0.5 | 4 |
| Ceftazidime | NR | NR | NR | 0.38 | 1 | 24 |
| Ceftazidime-avibactam | NR | NR | NR | 0.19 | 0.5 | 2 |
| Cefepime | 4 | 1 | 2 | 0.12 | 0.5 | 0.12 |
| Cefepime-enmetazobactam | 2 | 1 | 1 | ≤0.06 | 0.12 | ≤0.06 |
| Ceftolozane-tazobactam | >16 | >16 | >16 | 1 | 1.5 | 16 |
| Aztreonam | 16 | >16 | >16 | 8 | >16 | 2 |
| Aztreonam-avibactam | 2 | 1 | 4 | 0.12 | 0.5 | 0.12 |
| Cefiderocol | 1 | 2 | 2 | 0.06 | 0.25 | 0.25 |
| Ertapenem | 2 | ≤0.06 | 4 | ≤0.06 | 0.12 | ≤0.06 |
| Imipenem | ≤0.25 | 0.12 | 0.5 | 0.5 | 0.25 | 0.25 |
| Imipenem-relebactam | ≤0.25 | 0.12 | 0.5 | 0.25 | 0.5 | 0.25 |
| Meropenem | ≤0.25 | ≤0.06 | 0.25 | ≤0.06 | ≤0.06 | ≤0.06 |
| Meropenem-vaborbactam | ≤0.25 | ≤0.06 | 0.25 | ≤0.06 | ≤0.06 | ≤0.06 |
| Ciprofloxacin | >2 | >2 | >2 | NR | NR | NR |
| Levofloxacin | >2 | >2 | >2 | NR | NR | NR |
| Amikacin | 2 | 2 | 1 | NR | NR | NR |
| Gentamicin | >16 | >16 | >16 | NR | NR | NR |
| Tigecycline | 0.5 | ≤0.06 | 0.25 | NR | NR | NR |
| Eravacycline | 0.25 | ≤0.06 | 0.25 | NR | NR | NR |
| Colistin | 0.5 | 0.5 | 0.5 | NR | NR | NR |
| Specific activity (mU/mg) | IC50 (nM) | ||
|---|---|---|---|
| Cephalothin | Avibactam | Cloxacillin | |
| CMY-2 | 690 | 320 | 5.3 |
| CMY-42 | 562 | 400 | 3.7 |
| CMY-219 | 4 | 9,900 | 22,000 |
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial Identification and Susceptibility Testing · Antibiotics Pharmacokinetics and Efficacy
INTRODUCTION
Avibactam is the first non-β-lactam β-lactamase inhibitor active against ESBLs, AmpC, and serine carbapenemases. Accordingly, ceftazidime-avibactam (CAZ-AVI) is now considered as the reference for the treatment of infections caused by KPC and OXA-48 producers (1, 2). CAZ-AVI has demonstrated in vitro and clinical efficacy against carbapenemase-producing Enterobacterales, except against metallo-β-lactamase producers (3–5).
Resistance to CAZ-AVI mainly arises through increased expression and/or active-site mutations in class A β-lactamases (notably KPC) (6), often combined with reduced permeability, efflux upregulation, or PBP3 alterations (7–9). Few studies described mutations within CTX-M-15 or chromosomal AmpC in Enterobacterales, whether exposed in vitro to CAZ-AVI in Citrobacter freundii, Enterobacter cloacae (10), or in vivo after a patient’s exposure to Klebsiella pneumoniae and Klebsiella aerogenes (11, 12). Four CMY variants responsible for CAZ-AVI resistance have been identified in clinical isolates: CMY-178, CMY-185, and CMY-192 in Escherichia coli and CMY-172 in K. pneumoniae (13–16). These variants differ from CMY-2 by at least four amino acid substitutions or indels (16). CMY-42, a V231S CMY-2 variant, showed increased hydrolysis of ceftazidime and aztreonam (17). Here, we describe CMY-219, a single variant of CMY-42, selected after CAZ-AVI treatment in E. coli ST410.
Three clinical E. coli isolates (EC1-399F8, EC2-459F3, and EC3-511J2) from the same patient were submitted to the French National Reference Center (F-NRC) for Antimicrobial Resistance for investigation regarding carbapenemase production. MICs were determined by broth microdilution using customized Sensititre plates (Thermo Scientific, Les Ulis, France) and gradient strip (Liofilchem) for compounds not included or to extend the concentration range. Cefiderocol testing used the UMIC Cefiderocol (Bruker, Bremen, Germany). Results were interpreted using EUCAST guidelines 2025. Whole-genome sequencing was performed on a NextSeq 500 Illumina System. Assemblies were generated with Shovill v.1.1.0 and SPAdes v.3.14.0 and were analyzed for resistome, sequence type, and plasmid content on the CGE online platform (https://www.genomicepidemiology.org/). Genomic data are available under BioProject number PRJNA1279145.
EC1-399F8 was isolated at day 1 from a rectal swab of a patient recently diagnosed with pancreatic adenocarcinoma. This E. coli isolate produced an OXA-484 carbapenemase, along with CMY-42 and TEM-1 β-lactamases. This isolate was susceptible to CAZ-AVI (MIC = 1 mg/L) (Fig. 1, Table 1). One month later, the patient was admitted to the emergency department for a gangrenous acute cholecystitis with a fluid collection near the right colic angle and peritoneal effusion. Empirical antimicrobial therapy based on piperacillin-tazobactam first (2 days) then on imipenem (2 days) was suboptimal despite low MIC to imipenem (≤0.25 mg/L) (Fig. 1). After blood cultures turned positive, CAZ-AVI was initiated for 14 days. This treatment, combined with surgical management, including replacement of the biliary prosthesis, led to clinical resolution. Nine months later, EC2-459 F3 was recovered from a new intra-abdominal sample. Surprisingly, this isolate did not produce any carbapenemase but displayed resistance to CAZ-AVI (MIC = 16 mg/L) (Table 1). A new CMY-42 variant was identified, CMY-219, carrying a single G156D substitution according to the structural alignment-based numbering of class C β-lactamases scheme (18) (Fig. S1). A third isolate, EC3-511J2, produced both OXA-484 and CMY-219 and was also categorized as resistant to CAZ-AVI (Table 1).
Timeline of clinical events, antimicrobial treatments, and E. coli isolate collection in a patient treated with ceftazidime-avibactam for 14 days.
All three E. coli isolates belonged to ST410. SNP analysis supported within-patient clonality compared with all other OXA-484-producing ST410 sent to the F-NRC until December 2024 (n = 38) (Table S1). Among these 38 isolates, blaCMY was frequent (n = 35). Of note, no mutation was identified in the PBP2 or PBP3 sequences between the three isolates. The ST410-associated PBP3 polymorphisms (YRIN insertion and I536L), known to confer decreased susceptibility to ceftazidime, cefepime, aztreonam, and cefiderocol, were present (19, 20).
A total of eight plasmid replicons were identified in the genomes: IncY, IncFI1–IncFII, IncX3, IncIγ, IncFIB and IncQ1, and ColKP3. Contig alignment of EC1-399F8 revealed a partial match with known IncIγ-type plasmids pL3452210II_4 carrying blaCMY-42 (GenBank accession number NZ_CP076531) and pMB7671_5 carrying blaCMY-185 (NZ_CP127853) from E. coli (Fig. S2).
To assess the contribution of CMY-219 to CAZ-AVI resistance, blaCMY-2, blaCMY-42, and blaCMY-219 were amplified with their native upstream region using the primers CMY-For (5′-AACACACTGATTGCGTCTGACG-3′) and CMY-Rev (5′-AAGGAGGCCCAATATCCTGG-3′), cloned into pCR-Blunt II-TOPO, and electroporated into E. coli TOP10. Transformants expressing CMY-219 displayed a fourfold increase (0.25–4.0 mg/L) in CAZ-AVI MICs compared to those expressing CMY-42 or CMY-2. Conversely, MICs to all other cephalosporins except ceftolozane–tazobactam and cefiderocol were significantly decreased (Table 1). Ceftazidime MICs on cloxacillin-supplemented agar were higher for CMY-219 (24 mg/L) than for CMY-2/CMY-42 transformants (Table 1).
Crude extract assays were used to compare CMY activity and inhibition properties of AVI and cloxacillin in pTOPO-CMY-2, CMY-42, and CMY-219 transformants, as previously described (21). Ceftazidime hydrolysis was very low for CMY-2 and CMY-42 and undetectable for CMY-219. Cephalothin was therefore used as a reporter substrate for IC_50_ determination. CMY-2 and CMY-42 displayed comparable hydrolytic activities toward cephalothin, with specific activities of 690 and 562 mU/mg, respectively, whereas activity was approximately 150-fold lower for CMY-219 (4 mU/mg). This finding is consistent with the lower MICs observed for most β-lactams in pTOPO-CMY-219 transformants. Avibactam IC_50_ values were 27-fold higher for CMY-219 than for CMY-42, indicating reduced inhibition (Table 2). Cloxacillin IC_50_ values were also markedly increased (4,000-fold) for CMY-219 versus comparators, consistent with the cloxacillin-agar phenotype. Both avibactam and cloxacillin exhibited reduced inhibitory activity against CMY-219 in accordance with the observed increase in MICs.
Analysis of read depth did not support major blaCMY copy-number variation across clinical isolates (blaCMY/chromosomal gene ratio 1.0–1.6) (22). Promoter region revealed an identical genetic environment among the three isolates, suggesting that different levels of expression were not involved in CAZ-AVI resistance (Fig. S2).
This in vivo selection of CMY-219 is concerning because CAZ-AVI is a key option against OXA-48-like-producing Enterobacterales (1, 2). Notably, the cefepime–enmetazobactam combination remains active against these isolates, raising the question of its potential role as an alternative treatment for OXA-48-like Enterobacterales, possibly limiting avibactam-driven selective pressure and the emergence of CMY variants. Recently, a large-scale genomic analysis of 167,518 E. coli genomes from EnteroBase identified blaCMY-42 in 23.05% of ST410, 11.56% of ST167, and 5.90% of ST405 isolates (23). Worryingly, these successful clones also frequently harbor carbapenemase genes (mainly NDM-5, OXA-181, and OXA-48) and exhibit intrinsic reduced susceptibility to β-lactams due to PBP3 mutations (9).
Shropshire et al. reported that 48 days of CAZ-AVI treatment selected CMY-185 in an E. coli ST410 isolate (15). In their study, double or triple mutations in CMY-2, including N346Y, were required to reduce CAZ-AVI susceptibility (15). In contrast, a single G156D substitution in the H5 alpha helix of CMY-42 emerged after a single 14-day CAZ-AVI course. This represents the first description of a CAZ-AVI resistance-conferring mutation within this structural domain of CMY. Interestingly, CMY-219 lacks the N346 substitution shared by other CAZ-AVI-resistant CMY variants (CMY-172/CMY-178/CMY-185/CMY-192). It also retains all the other highly conserved residues already known to mediate avibactam binding (e.g., 4S, 67K, 120Q, 150Y, 152N, 315K, and 316T) supporting an alternative resistance route (17). In silico modeling using Chimera software suggested that introducing a bulky Asp at position 156 may perturb the local environment near the highly conserved Y150 in the active-site region. Concomitantly, the lower activity against several β-lactams suggests a functional trade-off between avibactam resistance and cephalosporin hydrolysis, as reported for CMY-185 with cephalothin (24). Further structural and kinetic work (inhibitor docking and acylation/deacylation parameters) will be required to define the molecular basis of G156D-mediated resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tamma PD, Aitken SL, Bonomo RA, Mathers AJ, van Duin D, Clancy CJ. 2023. Infectious Diseases Society of America 2023 guidance on the treatment of antimicrobial resistant gram-negative infections. Clin Infect Dis:ciad 428. doi:10.1093/cid/ciad 42837463564 · doi ↗ · pubmed ↗
- 2Paul M, Carrara E, Retamar P, Tängdén T, Bitterman R, Bonomo RA, de Waele J, Daikos GL, Akova M, Harbarth S, Pulcini C, Garnacho-Montero J, Seme K, Tumbarello M, Lindemann PC, Gandra S, Yu Y, Bassetti M, Mouton JW, Tacconelli E, Rodríguez-Baño J. 2022. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines for the treatment of infections caused by multidrug-resistant Gram-negative Bacilli (endorsed by European Society of Intensive Care Medicine). Clin Microbiol Inf · doi ↗ · pubmed ↗
- 3Bonnin RA, Bernabeu S, Emeraud C, Creton E, Vanparis O, Naas T, Jousset AB, Dortet L. 2022. Susceptibility of OXA-48-producing Enterobacterales to imipenem/relebactam, meropenem/vaborbactam and ceftazidime/avibactam. Int J Antimicrob Agents 60:106660. doi:10.1016/j.ijantimicag.2022.10666035988666 · doi ↗ · pubmed ↗
- 4Castanheira M, Doyle TB, Collingsworth TD, Sader HS, Mendes RE. 2021. Increasing frequency of OXA-48-producing Enterobacterales worldwide and activity of ceftazidime/avibactam, meropenem/vaborbactam and comparators against these isolates. J Antimicrob Chemother 76:3125–3134. doi:10.1093/jac/dkab 30634459890 PMC 8598286 · doi ↗ · pubmed ↗
- 5Aslan AT, Ezure Y, Horcajada JP, Harris PNA, Paterson DL. 2023. In vitro, in vivo and clinical studies comparing the efficacy of ceftazidime-avibactam monotherapy with ceftazidime-avibactam-containing combination regimens against carbapenem-resistant Enterobacterales and multidrug-resistant Pseudomonas aeruginosa isolates or infections: a scoping review. Front Med (Lausanne) 10:1249030. doi:10.3389/fmed.2023.124903037727767 PMC 10506411 · doi ↗ · pubmed ↗
- 6Hobson CA, Pierrat G, Tenaillon O, Bonacorsi S, Bercot B, Jaouen E, Jacquier H, Birgy A. 2022. Klebsiella pneumoniae carbapenemase variants resistant to ceftazidime-avibactam: an evolutionary overview. Antimicrob Agents Chemother 66:e 0044722. doi:10.1128/aac.00447-2235980232 PMC 9487638 · doi ↗ · pubmed ↗
- 7Nicolas-Chanoine M-H, Mayer N, Guyot K, Dumont E, Pagès J-M. 2018. Interplay between membrane permeability and enzymatic barrier leads to antibiotic-dependent resistance in Klebsiella pneumoniae. Front Microbiol 9:1422. doi:10.3389/fmicb.2018.0142230008709 PMC 6034560 · doi ↗ · pubmed ↗
- 8Nelson K, Hemarajata P, Sun D, Rubio-Aparicio D, Tsivkovski R, Yang S, Sebra R, Kasarskis A, Nguyen H, Hanson BM, Leopold S, Weinstock G, Lomovskaya O, Humphries RM. 2017. Resistance to ceftazidime-avibactam is due to transposition of KPC in a porin-deficient strain of Klebsiella pneumoniae with increased efflux activity. Antimicrob Agents Chemother 61:e 00989-17. doi:10.1128/AAC.00989-1728739787 PMC 5610495 · doi ↗ · pubmed ↗