First identification and molecular characterization of CTX-M-15 extended-spectrum β-lactamase and OXA-9 β-lactamase in Haemophilus influenzae in the Iberian Peninsula
Aida González-Díaz, Miguel Pinto, Irene Cadenas-Jiménez, Sara Duarte, Carmen Ardanuy, M. Manuela Ribeiro, Sara Martí, Paula Bajanca-Lavado

TL;DR
This study reports the first identification of CTX-M-15 and OXA-9 enzymes in Haemophilus influenzae in the Iberian Peninsula, highlighting growing antibiotic resistance.
Contribution
First report of CTX-M-15 and OXA-9 in H. influenzae in the Iberian Peninsula, with molecular characterization of resistance elements.
Findings
CTX-M-15 was identified in a multidrug-resistant H. influenzae strain from Portugal.
OXA-9 was found in a β-lactam and aminoglycoside-resistant H. influenzae strain from Spain.
Both strains carried novel integrative and conjugative elements (ICEs) associated with resistance.
Abstract
We describe, for the first time, the presence and characterization of CTX-M-15 and OXA-9 in two Haemophilus influenzae strains: PTHi-14525 (CTX-M-15) from Portugal and HUB-HI042681 (OXA-9) from Spain. Multidrug-resistant PTHi-14525 carried blaCTX-M-15 (two copies) and blaTEM-1 in an ICEHpaHUB5-like element. HUB-HI042681, resistant to β-lactams and aminoglycosides, carried a novel ICEHinHUB1. These findings highlight the genomic plasticity and expanded resistome of H. influenzae, raising concerns about treatment and emphasizing the need for continuous genomic surveillance.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
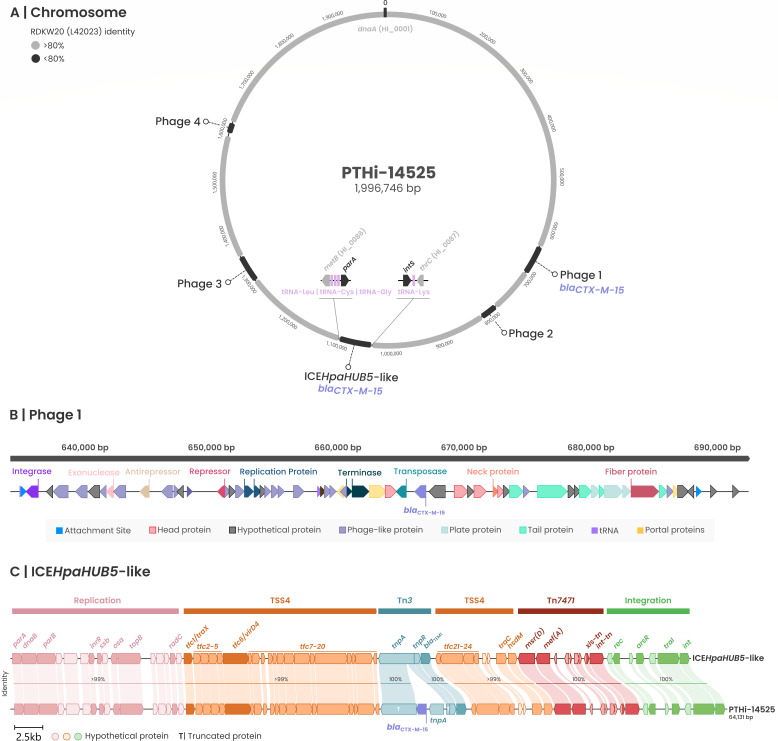 Fig 1
Fig 1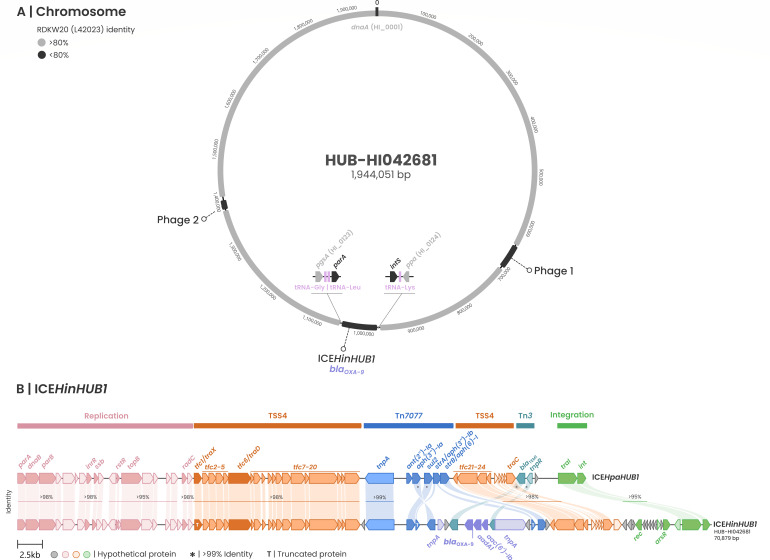 Fig 2
Fig 2| PTHi-14525 | HUB-HI042681 | |
|---|---|---|
| Antimicrobial agent | MIC (mg/L) | MIC (mg/L) |
| Ampicillin | >8 (R) | >4 (R) |
| Amoxicillin/clavulanate | 2 (S) | 2/1 (S) |
| Cefotaxime | >8 (R) | ≤0.06 (S) |
| Cefaclor | >16 (R) | – |
| Cefuroxime | >8 (R) | ≤0.5 (S) |
| Cefepime | 0.5 (R) | ≤0.25 (S) |
| Meropenem | 0.5 (S) | ≤0.25 (S) |
| Ciprofloxacin | ≤0.06 (S) | ≤0.03 (S) |
| Chloramphenicol | ≤1 (S) | ≤1 (S) |
| Tetracycline | ≤1 (S) | ≤1 (S) |
| Trimethoprim/sulfamethoxazole | >2 (R) | ≤0.5/9.5 (S) |
| Azithromycin | >4 (R) | 1 (S) |
- —European Commissionhttp://dx.doi.org/10.13039/501100000780
- —Instituto de Salud Carlos IIIhttp://dx.doi.org/10.13039/501100004587
- —Instituto de Salud Carlos IIIhttp://dx.doi.org/10.13039/501100004587
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumonia and Respiratory Infections · Antibiotic Resistance in Bacteria · Bacterial Infections and Vaccines
INTRODUCTION
Haemophilus influenzae is a major cause of respiratory tract infections and invasive disease. Historically regarded as highly susceptible to antimicrobial agents, this view changed in the early 1970s with the first reports of ampicillin-resistant strains due to TEM-1 β-lactamase production (1–3). In the early 1980s, amino acid substitutions were described in the penicillin-binding protein 3 (PBP3), reducing β-lactam binding affinity through point mutations or genetic recombination, contributing to the stepwise resistance in H. influenzae to ampicillin and third-generation cephalosporins (4–7). The evolution of resistance in H. influenzae has been accelerated by horizontal gene transfer, through the acquisition of transposons and integrative and conjugative elements (ICEs) (8, 9). Of concern is the potential transfer of extended-spectrum β-lactamase (ESBL) genes from closely related species. The recent identification of the ESBL blaCTX-M-15, in a multidrug-resistant (MDR) H. parainfluenzae strain in Spain and France positions this species as a potential reservoir (10, 11). Given the natural competence of H. influenzae, the acquisition of such ESBL genes would represent a significant shift, layering potent enzymatic resistance onto existing mechanisms (12, 13). This could compromise the efficacy of current treatments for invasive H. influenzae infections, reinforcing the importance of monitoring interspecies transfer of resistance mechanisms to contain the dissemination of multidrug resistance (14, 15). The growing burden of β-lactam resistance has placed H. influenzae ampicillin-resistant on the WHO priority list for new antibiotics, a challenge further intensified by the emergence of strains resistant to three or more antibiotic classes (MDR) and to more than five classes (extensively drug-resistant). These strains are associated with ICEs, which facilitate the spread of resistance among genetically diverse species (16, 17).
Here, we report the first documented cases of acquired β-lactamase-mediated resistance involving blaCTX-M-15 and blaOXA-9 in H. influenzae detected in the Iberian Peninsula. The National Reference Laboratory for Haemophilus influenzae (NRLHi) at the Portuguese National Institute of Health coordinates nationwide surveillance of invasive H. influenzae. The first resistant isolate PTHi-14525, from Porto, was received at NRLHi for antimicrobial resistance confirmation and whole-genome sequencing (WGS) characterization. The second isolate HUB-HI042681 was obtained in the Hospital Universitari de Bellvitge (HUB), a tertiary hospital located in the Southern Barcelona area (Spain) and analyzed as a part of a surveillance program in Catalonia. Antimicrobial susceptibility was tested by broth microdilution using MICroSTREP plus panels (PTHi-14525) and STRHAE2 Sensititre panels (HUB-HI042681) (Table 1). Breakpoints were interpreted following European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines (18).
Genomic DNA was extracted using either the QIAsymphony (PTHi-14525) or MagMax DNA Ultra 2.0 for KingFisher Flex (HUB-HI042681). For short-read sequencing, libraries were prepared using Nextera XT (PTHi-14525) and Illumina DNA Prep (HUB-HI042681), followed by paired-end sequencing on Illumina platforms. For long-read sequencing, libraries were prepared using the Native Barcoding Kit (SQK-NBD114.24) and sequenced on R10.4.1 flow cells (Oxford Nanopore Technologies). Bioinformatic analysis was conducted using Bactopia v3.0.0 (19), which includes hybrid assembly (Unicycler) (20), annotation (Prokka) (21), sequence typing (22), and antibiotic resistance scan (AMRFinder+) (23). Amino acid substitutions in PBP3, GyrA, ParC, DHFR, and DHPS were screened using Geneious R9. Species confirmation was conducted using rMLST (pubmlst.org/species-id). Assemblies were re-annotated using H. influenzae RdKW20 (L42023) as a reference, with a minimum identity threshold of 80%. Prophages were identified with PHASTEST (24), and only intact regions were analyzed. Mobile genetic elements harboring antibiotic resistance genes were aligned with Mauve against ICEHin1056 (AJ627386), ICEHin299 (AM884334), ICEHpa8f (AM884335), ICEHpaT3T1 (FQ312002: 1154165–1212168), ICEHpaHUB1–ICEHpaHUB4 (25), and ICEHpaHUB5 (26). Annotation of the novel ICEHinHUB1 was refined by manually curating Prokka annotations, using Juhas et al. (9) as a reference in combination with BLASTx and UniProt searches. Representations were generated using the Geneious backbone, PHASTEST outputs, and CAGECAT (cagecat.bioinformatics.nl/https://cagecat.bioinformatics.nl/https://cagecat.bioinformatics.nl/). PTHi-14525 (ERR13337076, ERR15705790) and HUB-HI042681 (ERR15760486, ERR15760484) raw reads were uploaded to the European Nucleotide Archive and ICEHinHUB1 was deposited at NCBI (PX634569).
Genomic analysis identified, for the first time, the presence of the ESBL enzyme CTX-M-15 in a Portuguese isolate (PTHi-14525) and the narrow-spectrum β-lactamase (NSBL) OXA-9 enzyme in a Spanish isolate (HUB-HI042681). The first case occurred in March 2024 and involved a 67-year-old male patient who was transferred from another hospital and admitted to the Intensive Care Unit of Unidade Local de Saúde de São João with community-acquired pneumonia complicated by pulmonary septic shock, in which H. influenzae was isolated from a bronchoalveolar lavage. The second case involved a 68-year-old female with a history of classical mycosis fungoides diagnosed in 2009. In March 2024, she presented to the emergency department with bacteremic pneumonia, and H. influenzae was isolated from both blood cultures and bronchoalveolar lavage. PTHi-14525 was resistant to ampicillin, cefotaxime, cefuroxime, cefepime, trimethoprim-sulfamethoxazole and azithromycin (Table 1). β-lactam resistance was explained by amino acid substitutions D350N, G490E, N526K, and A530S in the PBP3, the acquisition of blaCTX-M-15 (two gene copies) and blaTEM-1. HUB-HI042681 showed resistance to penicillin, ampicillin, amikacin, gentamicin, and tobramycin (Table 1) exhibiting PBP3 substitutions D350N, M377I, A502V, and N526K, and carried the blaOXA-9 and blaTEM-1 genes. PTHi-14525 was resistant to trimethoprim-sulfamethoxazole due to the I95L substitution in DHFR and an SFLYND insertion at position 65 of DHPS. Macrolide resistance was only observed in PTHi-14525 and was explained by the presence of mef(A) and msr(D). On the other hand, HUB-HI042681 carried the aminoglycoside resistance genes aadA1, aac(6′)-Ib, and aph(3′)-Ia. Both assemblies yielded complete genomes in a single contig.
The strains were nonencapsulated and showed 100% ribosomal MLST support for H. influenzae. Figure 1A depicts the chromosome of PTHi-14525 (ST2930), while Fig. 2A shows that of HUB-HI042681 (ST136). Four intact prophages were identified in PTHi-14525 using PHASTEST, and the predicted proteins are summarized in Supplementary Data set 1A. Predicted prophage 1 harbors the first copy of blaCTX-M-15, as shown in Fig. 1B. The second copy of blaCTX-M-15 was integrated together with a tnpA transposase within the Tn3 transposon of a previously described ICEHpaHUB5-like element (identity >99%) (26), which also carries a Tn7471 containing the msr(D) and mef(A) genes. HUB-HI042681 contained two prophages identified by PHASTEST and a novel ICE (ICEHinHUB1), carrying blaOXA-9, blaTEM-1, aadA1, and aac(6′)-Ib. The genetic structure of ICEHinHUB1 is compared with its closest relative, ICEHpaHUB1 (Fig. 2A). High identity was observed in the replication and T4SS genes, whereas significant differences were detected in the Tn7077 region (Fig. 2B). The analysis of the genomic contexts revealed different arrangements of β-lactamase genes in the two studied isolates. PTHi-14525 harbored two copies of blaCTX-M-15, one integrated into a prophage and another within a Tn3 transposon together with the blaTEM-1 gene, located in an ICEHpaHUB5-like element previously described (26). In contrast, the blaOXA-9 in HUB-HI042681 was identified in a novel ICE, designated ICEHinHUB1, in which transposons Tn7077 and Tn3 had merged into a new rearrangement. This new structure harbored multiple aminoglycoside resistance genes and the blaTEM-1 gene.
(A) Schematic representation of the PTHi-14525 chromosome. Regions sharing more than 80% sequence identity with H. influenzae Rd KW20 are shown in light gray. The four intact prophages detected by PHASTEST are indicated in dark gray, along with the ICEHparHUB5-like element and its chromosomal insertion site (gene names and Rd KW20 locus tag), tRNA (pink) and the first (parA) and last (intS) ICE genes. The blaCTX-M-15 genes are highlighted in purple. (B) Phage carrying the first copy of blaCTX-M-15. The genomic positions and gene types predicted by PHASTEST are shown. (C) Mauve alignment representation of the PTHi-14525 ICE elements carrying the second copy of the blaCTX-M-15 and the previously described ICEHparHUB5-like element, with pairwise sequence identity indicated for each region.
(A) Schematic representation of the HUB-HI042681 chromosome. Regions sharing more than 80% sequence identity with H. influenzae Rd KW20 are shown in light gray. The two intact prophages detected by PHASTEST are indicated in dark gray, along with the ICEHinHUB1 element and its chromosomal insertion site (gene names and Rd KW20 locus tag), tRNA (pink), and the first (parA) and last (intS) ICE genes. The blaOXA-9 gene is highlighted in purple. (B) Mauve alignment representation of the new ICEHinHUB1 elements carrying the blaOXA-9 and the previously described ICEHparHUB1 element, with pairwise sequence identity indicated for each region.
Recent studies have already identified the CTX-M-15 β-lactamase in H. parainfluenzae isolates from Spain and France (10, 11). While the urogenital origin of the CTX-M-15-positive H. parainfluenzae isolates precludes a direct ecological connection to respiratory H. influenzae, their existence demonstrates that this β-lactamase can be acquired by Haemophilus species. This establishes a precedent, suggesting that those H. influenzae isolated in the respiratory tract could similarly acquire the blaCTX-M-15 gene from other co-colonizing bacteria, reinforcing the role of such niches as potential reservoirs for resistance genes. The detection of H. influenzae isolates combining β-lactamases (either ESBL or NSBL), PBP3 amino acid substitutions, and additional resistance genes within mobile genetic elements underscores the species’ genomic adaptability to acquire and disseminate multiple resistance determinants (13–15, 27, 28). Both isolates likely acquired the resistance genes through horizontal transfer from co-colonizing species, facilitating the development of resistant strains capable of invading the bloodstream. Moreover, the patient carrying the β-lactamase CTX-M-15–producing isolate (PTHi-14525) had been transferred between at least three hospital units in two hospitals, suggesting a possible healthcare-associated acquisition and interspecies gene transfer from other hospital-associated pathogens (29, 30). This situation underscores the urgent need for ongoing surveillance, particularly through WGS, which allows the rapid detection of both novel and known resistance genes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mathies AW Jr. 1972. Penicillins in the treatment of bacterial meningitis. J R Coll Physicians Lond 6:139–146.5061041 PMC 5366020 · pubmed ↗
- 2Khan W, Ross S, Rodriguez W, Controni G, Saz AK. 1974. Haemophilus influenzae type B resistant to ampicillin. A report of two cases. JAMA 229:298–301.4546094 · pubmed ↗
- 3Tomeh MO, Starr SE, Mc Gowan JE Jr, Terry PM, Nahmias AJ. 1974. Ampicillin-resistant Haemophilus influenzae type B infection. JAMA 229:295–297.4546093 · pubmed ↗
- 4Ubukata K, Shibasaki Y, Yamamoto K, Chiba N, Hasegawa K, Takeuchi Y, Sunakawa K, Inoue M, Konno M. 2001. Association of amino acid substitutions in penicillin-binding protein 3 with beta-lactam resistance in beta-lactamase-negative ampicillin-resistant Haemophilus influenzae. Antimicrob Agents Chemother 45:1693–1699. doi:10.1128/AAC.45.6.1693-1699.200111353613 PMC 90533 · doi ↗ · pubmed ↗
- 5Deghmane AE, Hong E, Chehboub S, Terrade A, Falguières M, Sort M, Harrison O, Jolley KA, Taha MK. 2019. High diversity of invasive Haemophilus influenzae isolates in France and the emergence of resistance to third generation cephalosporins by alteration of fts I gene. J Infect 79:7–14. doi:10.1016/j.jinf.2019.05.00731100360 · doi ↗ · pubmed ↗
- 6Cherkaoui A, Francois P, Gaia N, Renzi G, Fischer A, Schrenzel J. 2025. Extensively drug-resistant Haemophilus influenzae isolated in Geneva, Switzerland. Eur J Clin Microbiol Infect Dis 44:1273–1277. doi:10.1007/s 10096-025-05093-w 40048099 PMC 12062098 · doi ↗ · pubmed ↗
- 7Diricks M, Petersen S, Bartels L, Lâm TT, Claus H, Bajanca-Lavado MP, Hauswaldt S, Stolze R, Vázquez OJ, Utpatel C, Niemann S, Rupp J, Wohlers I, Merker M. 2024. Revisiting mutational resistance to ampicillin and cefotaxime in Haemophilus influenzae. Genome Med 16:140. doi:10.1186/s 13073-024-01406-439633433 PMC 11616347 · doi ↗ · pubmed ↗
- 8Juhas M, Crook DW, Dimopoulou ID, Lunter G, Harding RM, Ferguson DJP, Hood DW. 2007. Novel type IV secretion system involved in propagation of genomic islands. J Bacteriol 189:761–771. doi:10.1128/JB.01327-0617122343 PMC 1797279 · doi ↗ · pubmed ↗