Clonal and plasmid-mediated dissemination of CTX-M-14–producing Escherichia coli within a single cattle farm in Japan
Ryuichi Nakano, Yuki Suzuki, Akiyo Nakano, Koichi Yamaguchi, Saori Horiuchi, Yasuo Ono, Hisakazu Yano

TL;DR
This study shows how antibiotic-resistant E. coli spreads between cattle and a farmer, highlighting the role of plasmids in transferring resistance genes.
Contribution
The study provides direct genomic evidence of CTX-M-14 dissemination between cattle and a farmer via IncI1 plasmids.
Findings
Seven CTX-M-14-positive E. coli isolates were found, with two showing recent clonal transmission between a parent cattle and calf.
All CTX-M-14 genes were located on nearly identical IncI1 plasmids, indicating plasmid-mediated spread.
Conjugation experiments confirmed high plasmid transferability, supporting cross-host resistance gene flow.
Abstract
Extended-spectrum β-lactamase (ESBL)–producing Escherichia coli threatens humans and animals. blaCTX-M-14, which is prevalent in Asia, is mainly disseminated via incompatibility group I1 (IncI1) plasmids, as they can efficiently transfer across Enterobacterales. However, direct evidence linking livestock and farmers at the genomic level is limited, and our study addresses this concern. Fecal samples were collected in 2013 from one parent cattle, three calves, and a farmer on a cattle farm in southern Kyushu, Japan. ESBL-producing E. coli were identified and characterized using hybrid Illumina–Nanopore assemblies, resistance gene profiling, and plasmid replicon typing. Clonal relatedness was assessed using core-genome multilocus sequence typing and core single-nucleotide polymorphism (SNP) analysis. Conjugation assays were used to evaluate plasmid transferability. Seven…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
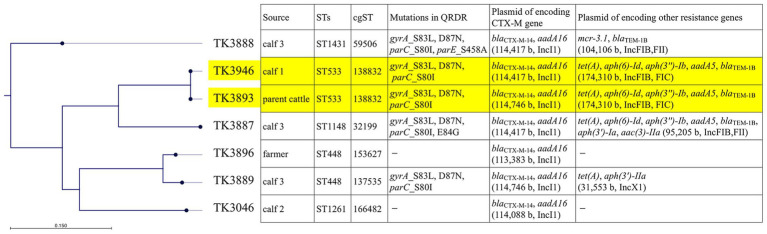 Figure 1
Figure 1 Figure 2
Figure 2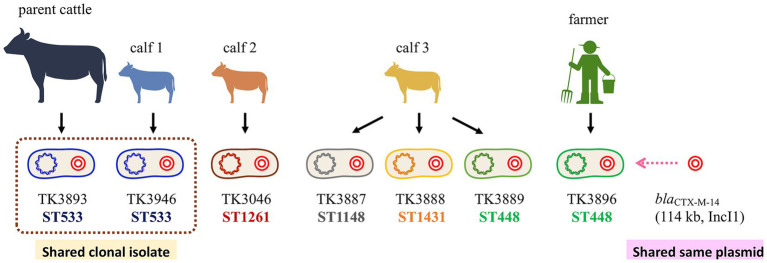 Figure 3
Figure 3| Strains | Source | Sequence types | cgST | CTX-M gene | Other resistance genes | MIC (μg/mL) | Transconjugant profile | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTX | CTX/CLAb | CAZ | CMZ | AZT | LEV | TET | KAN | GEN | COL | Transfer frequencies | Plasmid replicon type | Resistance genes | Plasmid size | ||||||
| TK3893 | parent cattle | ST533 | 138,832 | CTX-M-14 | 16 | 0.125 | 2 | 1 | 2 | 8 | >256 | 2 | 1 | 1 | 3.2 × 10−3 | I1 | CTX-M-14 | 114,746b | |
| TK3946 | calf 1 | ST533 | 138,832 | CTX-M-14 | 8 | ≦0.06 | 2 | 1 | 2 | 16 | >256 | 2 | 1 | 1 | 9.1 × 10−3 | I1 | CTX-M-14 | 114,417b | |
| TK3046 | calf 2 | ST1261 | 166,482 | CTX-M-14 | - | 8 | ≦0.06 | 2 | 0.5 | 2 | ≦0.06 | 8 | 2 | 0.5 | 1 | 3.1 × 10−3 | I1 | CTX-M-14 | 114,088b |
| TK3887 | calf 3 | ST1148 | 32,199 | CTX-M-14 | 16 | 0.125 | 2 | 1 | 4 | 64 | >256 | >256 | 128 | 1 | 3.3 × 10−3 | I1 | CTX-M-14 | 114,417b | |
| TK3888 | calf 3 | ST1431 | 59,506 | CTX-M-14 | 16 | ≦0.06 | 2 | 0.5 | 2 | 16 | 16 | 2 | 1 | 16 | 1.6 × 10−2 | I1 | CTX-M-14 | 114,417b | |
| TK3889 | calf 3 | ST448 | 137,535 | CTX-M-14 |
| 8 | ≦0.06 | 1 | 0.5 | 1 | 16 | >256 | 128 | 1 | 1 | 2.6 × 10−3 | I1 | CTX-M-14 | 114,746b |
| TK3896 | farmer | ST448 | 153,627 | CTX-M-14 | - | 16 | ≦0.06 | 2 | 0.5 | 2 | ≦0.06 | 8 | 2 | 0.5 | 1 | 7.2 × 10−2 | I1 | CTX-M-14 | 113,383b |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Escherichia coli research studies · Infections and bacterial resistance
Introduction
1
Extended-spectrum β-lactamase (ESBL)–producing Escherichia coli are a major global health concern, as they undermine the efficacy of third-generation cephalosporins in both human and veterinary medicine (Tacconelli et al., 2018; Castanheira et al., 2021). Among ESBLs, CTX-M enzymes have become predominant worldwide (Bonnet, 2004; Bush and Bradford, 2020), with CTX-M-14 being one of the most frequently reported variants. In Japan, blaCTX-M-14 has been repeatedly identified in both clinical and livestock isolates, indicating its wide establishment across reservoirs (Kosai et al., 2020; Masui et al., 2022; Nakano et al., 2023). Similar trends have been observed in other Asian countries, highlighting their regional significance (Liao et al., 2015; Bevan et al., 2017).
blaCTX-M-14 is frequently encoded on plasmids and can disseminate through horizontal gene transfer among Enterobacteriaceae (Cantón et al., 2012; Yu et al., 2024). Particularly, incompatibility group I1 (IncI1) plasmids serve as key vectors; they are characterized by high conjugation efficiency, broad host range, and remarkable stability, facilitating their persistence and dissemination (Carattoli, 2009; Wong et al., 2016; Villa and Carattoli, 2020; Yu et al., 2024). These features make IncI1 plasmids highly effective vehicles for interlineage and interhost transmission of resistance genes.
Whole-genome sequencing (WGS)-based approaches, such as core-genome multilocus sequence typing (cgMLST) and core single-nucleotide polymorphism (core-SNP) analysis, allow discrimination of recent clonal transmission from coincidental sequence type (ST) overlap (Yan et al., 2023; Kavanagh et al., 2024). Studies applying these tools often demonstrate that human and animal isolates from the same farm cluster genetically or share nearly identical plasmids, supporting the potential for interspecies exchange (de Been et al., 2014; Aldea et al., 2022). However, definitive one-to-one evidence of direct transmission, such as ≤ 0–5 SNP differences between a farmer and their livestock, remains rare.
Previously, we reported the genetic relatedness of third-generation cephalosporin-resistant E. coli isolated from livestock, farmers, and patients in southern Kyushu, Japan (Nakano et al., 2023). The study demonstrated the occurrence of resistant E. coli across multiple hosts and highlighted the potential for transmission between human and animal populations. However, the relative contribution of direct clonal spread vs. plasmid-mediated transfer remained unclear.
In this study, using high-resolution whole-genome sequencing approaches, we aimed to investigate the clonal relatedness and plasmid characteristics of blaCTX-M-14-positive E. coli isolated from cattle and a farm worker on a single farm in Japan. Using a combination of antimicrobial susceptibility testing, hybrid WGS, cgMLST, core-SNP analysis, and plasmid characterization, we (i) determined the clonal relatedness between isolates from cattle and farmer, (ii) characterized the genomic features of blaCTX-M-14–encoding plasmids, and (iii) evaluated the conjugative transferability of these plasmids. By providing genome-level evidence of both clonal dissemination among livestock and the sharing of nearly identical IncI1 plasmids between livestock and a human host, this study extends previous findings and contributes to a deeper understanding of ESBL transmission dynamics in the farm environment.
Materials and methods
2
Isolation of third-generation cephalosporin-resistant E. coli from cattle and farmer
2.1
Fecal samples were collected from cattle and a farmer on a single cattle farm in the southern part of Kyushu Island, a major cattle farming area in Japan, in 2013. The fecal samples were directly inoculated onto deoxycholate-hydrogen sulfide-lactose agar plates and incubated at 37 °C for 24 h. Three colonies were randomly selected, and the species were identified using matrix-assisted laser desorption ionization-time-of-flight mass spectrometry (Vitek MS system; bioMérieux, Co., Ltd.). The isolates identified as E. coli were used for analyses. The Ethical Review Committee of the Teikyo University School of Medicine (No. 13–118) approved the study protocol, and the farmer provided written informed consent to participate in the study.
Antimicrobial susceptibility testing and detection of ESBL genes
2.2
Antimicrobial susceptibility of the isolates was determined through agar microdilution according to the Clinical & Laboratory Standards Institute guidelines, and quality control was performed using the reference strain E. coli ATCC 25922 (CLSI, 2025). The presence of ESBL genes in the third-generation cephalosporin-resistant E. coli isolates was identified using multiplex PCR (Dallenne et al., 2010). Gene-specific PCR was performed to identify the genotype, and the amplified products were confirmed using DNA sequencing (Ohnishi et al., 2013). Sequence alignment and analysis were performed on the National Center for Biotechnology Information website using the Basic Local Alignment Search tool (Altschul et al., 1990)1.
WGS and genomic analysis
2.3
Genomic DNA was extracted using the QIAGEN Genomic-tip 500/G kit (Qiagen, Germany). WGS was performed on both the Illumina MiSeq platform (Illumina Inc., USA) and Oxford Nanopore MinION platform (Oxford Nanopore Technologies, UK). Hybrid de novo assemblies were generated using Unicycler v0.5.0 (Wick et al., 2017), and genome annotation was conducted via DFAST v1.6.0 on default parameters.
Plasmid replicon types were determined with PlasmidFinder, and plasmid/genome structures were compared and visualized using Easyfig v2.2.2. Antimicrobial resistance genes were identified using ResFinder2 with thresholds ≥90% identity and ≥60% minimum length. Multilocus sequence typing (MLST) of E. coli isolates was performed, and STs were assigned using the PubMLST database3 (Wirth et al., 2006). Pairwise nucleotide identity among blaCTX-M-14-encoding plasmids was evaluated using the JSpeciesWS online platform4 (Richter and Rosselló-Móra, 2009). Average Nucleotide Identity based on MUMmer (ANIm) was calculated for all plasmid pairs under default settings.
cgMLST
2.4
cgMLST was performed using cgMLSTFinder 1.2 (Center for Genomic Epidemiology).5 Assembled genomes of all isolates were submitted to the pipeline, and allele calling was performed against the E. coli cgMLST scheme (2,513 loci). The output data included the total number of called alleles, ST assignments [core-genome Sequence Type (cgST)], and allelic distances between isolates. A minimum-spanning tree was constructed from the cgMLST allelic profiles using GrapeTree v2.2 (Zhou et al., 2018). The resulting phylogenetic relationships were further visualized and annotated with FigTree v1.4.4.6 Isolates with identical cgSTs and zero allelic distance were considered clonally indistinguishable, whereas those with small allelic differences (≤10 alleles) were regarded as highly related, according to previously described criteria (Zhou et al., 2021).
Chromosomal core-SNP analysis
2.5
Chromosomal core-SNP analysis was performed to assess the genetic relatedness. Therefore, plasmid sequences were excluded, and only chromosomal sequences were used. Sequence reads were mapped to the E. coli TK3888 genome (GenBank accession no. AP044770), which was used as the reference, using the CLC Genomics Workbench v24.0 (Qiagen) with default mapping parameters. To ensure accuracy, SNPs were called under the following thresholds: minimum coverage of 10×, minimum variant frequency of 90%, and minimum base quality score of 20. Putative SNPs located in repetitive or low-complexity regions were excluded, and only high-confidence SNPs in conserved chromosomal regions were retained for downstream analyses.
An SNP alignment was generated from all isolates, and a maximum-likelihood phylogenetic tree was constructed in CLC Genomics Workbench using the Jukes–Cantor model with 1,000 bootstrap replicates. Pairwise SNP distances were also calculated to determine the degree of genetic relatedness between isolates. Based on prior studies on Enterobacteriaceae, isolates differing by 0–5 core-SNPs were considered clonally identical, whereas larger distances (e.g., >15–25 core-SNPs) were interpreted cautiously as indicative of more distant relationships (Dallman et al., 2015; Ludden et al., 2021).
Plasmid conjugation assay
2.6
Conjugation experiments were performed using the broth mating method with E. coli J53 (sodium azide–resistant) as the recipient, as previously described (Masui et al., 2022). Briefly, donor and recipient strains were grown to the exponential phase in Luria–Bertani (LB) broth, mixed at a 1:1 ratio (vol/vol), and incubated overnight at 37 °C without shaking. After which, aliquots of the mating mixtures were plated onto LB agar supplemented with cefpodoxime (8 μg/mL) and sodium azide (100 μg/mL) to select for transconjugants. The presence of the resistance gene blaCTX-M-14 in transconjugants was confirmed using PCR. Conjugation frequency was calculated as the number of transconjugant colonies (cfu/mL) divided by the total number of recipient cells (cfu/mL).
Results
3
Characteristics of ESBL-producing E. coli isolates from cattle and farmer
3.1
Overall, 15 isolates (three each from one parent cattle, three calves, and the farmer) were obtained and identified as 13 E. coli and 2 Klebsiella pneumoniae isolates. Among them, seven E. coli isolates exhibited resistance to third-generation cephalosporins with an ESBL phenotype (Table 1). One isolate each was recovered from the parent cattle, two calves, and the farmer, whereas three isolates were obtained from the remaining calf. These seven isolates displayed diverse susceptibility profiles to other antimicrobials. Five isolates were resistant to levofloxacin and carried mutations in the quinolone resistance-determining regions of gyrA and parC (Figure 1). Four isolates carried tet(A) and exhibited resistance to tetracycline, two of which also carried aph(3′)-Ia or aph(3′)-IIa and were resistant to kanamycin. All seven isolates harbored the blaCTX-M-14 gene. Additionally, two isolates obtained from calf 3 carried further resistance determinants: TK3887 harbored aac(3)-IIa and was resistant to gentamicin, whereas TK3888 carried mcr-3.1 and was resistant to colistin.
Core-genome SNP-based phylogenetic tree showing the isolation source, sequence types, mutations in QRDR, and plasmid contents of E. coli from parent cattle, calves, and a farmer. The tree was constructed based on core-genome SNPs using the General Time Reversible (GTR) model with 1,000 bootstrap replicates. The accompanying metadata matrix displays relevant genomic and phenotypic attributes for each isolate. The two E. coli isolates belonging to ST533 (from the parent cattle and calf 1), which showed clonal identity (one SNP difference and identical cgST), are highlighted with a yellow background to emphasize their close genetic relatedness. SNP, single-nucleotide polymorphism; QRDR, quinolone resistance–determining region; ST, sequence type; cgST, core genome ST.
MLST analysis revealed five distinct STs among the seven isolates (Table 1). Notably, the three isolates recovered from calf 3 belonged to different STs. Shared STs were identified between the parent cattle and calf 1 (ST533), as well as between calf 3 and the farmer (ST448).
Clonal relatedness of E. coli isolates between cattle and farmer
3.2
To investigate the genetic relatedness of the isolates, both cgSNP analysis and cgMLST were performed (Table 1; Figure 1). The two ST533 isolates from the parent cattle (TK3893) and calf 1 (TK3946) differed by only one SNP in the core genome, strongly suggesting clonal identity. Consistently, both isolates were assigned to the same cgST (138832), further confirming their close genetic relationship. This represents a direct example of clonal dissemination between livestock individuals within the same farm.
Contrastingly, ST448 isolates from calf 3 (TK3889) and the farmer (TK3896) differed by 3,891 core SNPs and had distinct cgSTs (137,535 and 153,627), showing a distant relation, despite their sharing of the same ST (Supplementary Figure 1). Other isolates (ST1261, ST1148, and ST1431) differed by over 15,000 SNPs with distinct cgSTs, consistent with independent lineages. The three isolates from calf 3 belonged to different STs and cgSTs, indicating coexistence of multiple unrelated blaCTX-M-14-positive E. coli lineages within a single animal.
These findings highlight clonal dissemination among livestock (ST533) and demonstrate that shared STs alone (e.g., ST448) do not necessarily indicate recent transmission, underscoring the value of integrating SNP and cgMLST analyses.
Genomic features of CTX-M-14–encoding plasmids
3.3
Hybrid assembly and comparative analysis revealed that the blaCTX-M-14 gene in all positive isolates was consistently located on highly similar IncI1-type plasmids of approximately 114 kb in size (Figure 2). These plasmids exhibited a conserved backbone structure with only minor variations in accessory regions. Pairwise ANIm analysis using JSpeciesWS demonstrated extremely high sequence similarity among the seven IncI1 plasmids, with ANIm values ranging from 99.98 to 100%, and alignment coverage between 99.1 and 100% (Supplementary Table 1). These results clearly indicate that nearly identical blaCTX-M-14-encoding IncI1 plasmids were shared among the cattle and farmer within the same farm.
Linear comparison of blaCTX-M-14-carrying plasmid sequences from parent cattle, calves, and a farmer with plasmid p15A44 in Japan, and exposition of the detailed genetic structures around blaCTX-M-14. A legend explains the color code for gene functions. A magnified gene cluster with labels illustrates key resistance elements. Gray shading indicates sequence similarity, and sequence sizes are shown in base pairs. The GenBank accession numbers were: p15A44 (LC567051), pTK3893 (LC903451), pTK3946 (LC903453), pTK3046 (AP044764), pTK3887 (AP044768), pTK3888 (AP044771), pTK3889 (AP045037), and pTK3896 (AP045044).
No additional resistance genes were encoded on these IncI1 plasmids, indicating that they primarily functioned as dedicated vectors for the dissemination of blaCTX-M-14. Other resistance determinants—mcr-3.1, aac(3)-IIa, aph variants, tet(A), and blaTEM-1B—were carried on separate plasmids belonging to different incompatibility groups (e.g., IncFIB/FII, IncFIB/FIC, IncX1; Figure 1), highlighting the diversity of plasmid backgrounds contributing to multidrug resistance within the same farm.
Notably, the two clonally identical ST533 isolates from the parent cattle (TK3893) and calf 1 (TK3946) carried indistinguishable IncI1 plasmids, as well as nearly identical IncFIB/FIC plasmids encoding tet(A) and aminoglycoside resistance genes. Moreover, the blaCTX-M-14-encoding IncI1 plasmids exhibited high sequence similarity (>99.9%) to an IncI1 plasmid deposited in GenBank (accession no. LC567051), based on BLAST analysis (Figure 2). This reference plasmid was isolated from a clinical E. coli strain in Japan in 2015. This finding is consistent with the possibility that highly conserved plasmids can disseminate across human and livestock reservoirs.
Conjugative transferability of CTX-M-14 plasmids
3.4
Conjugation experiments were performed with all seven CTX-M-14–producing E. coli isolates. The transfer frequencies of the plasmids ranged from 10^−2^ to 10^−3^ per recipient, indicating a relatively high conjugation potential (Table 1). All transconjugants consistently carried only the blaCTX-M-14–encoding IncI1 plasmid, whereas plasmids harboring additional resistance determinants, such as aac(3)-IIa and mcr-3.1, were not co-transferred. The antimicrobial susceptibility profiles of the transconjugants were uniform, and their MIC values for third-generation cephalosporins were comparable across all donor strains (Supplementary Table 2).
Discussion
4
Presence of CTX-M-14–producing E. coli in cattle and farmer
4.1
To the best of our knowledge, this is the first report in Japan providing genome-level evidence that nearly identical IncI1 plasmids carrying blaCTX-M-14 were shared between humans and animals in a farm. CTX-M-14–producing E. coli are frequently isolated from livestock (Hayashi et al., 2018; Nakano et al., 2023) and prevalent among individuals (Yano et al., 2013; Komatsu et al., 2018; Masui et al., 2022). This suggests widespread dissemination across human and animal reservoirs. Similar patterns have been reported in other Asian countries, especially China, highlighting the regional significance of CTX-M-14 in both clinical and agricultural contexts (Liu et al., 2018; Zheng et al., 2019; Chen et al., 2024).
A major factor in this predominance is the association of blaCTX-M-14 with IncI1 plasmids, exhibiting high conjugation efficiency, broad host range within Enterobacterales, and stability during bacterial replication (Cottell et al., 2011; Di Pilato et al., 2019). These properties facilitate persistence and rapid dissemination across bacterial lineages, host species, and ecological niches (Beyrouthy et al., 2021). The strong linkage between blaCTX-M-14 and IncI1 plasmids likely underlies the successful establishment and widespread distribution of blaCTX-M-14-positive E. coli in both clinical and livestock environments.
Notably, among all isolates recovered from cattle and the farmer, blaCTX-M-14 was the only ESBL gene detected. The absence of other ESBL types within the same farm suggests a limited diversity of ESBL-producing E. coli at the time of sampling, which may reflect a relatively restricted introduction of ESBL determinants into this farm environment.
STs and clonal spread
4.2
cgSNP analysis revealed that E. coli isolates from the parent cattle and calf 1 belonging to ST533 differed by only a single SNP, indicating clonal identity. Consistent with this, cgMLST analysis noted that these two ST533 isolates shared the same cgST, further supporting their close genetic relationship. This represents an example in which a parent cattle and her calf on the same farm shared a clonally identical strain at the chromosomal level (0–1 core SNP difference), a finding that is biologically plausible given the close contact between animals. Such a precise one-to-one chromosomal match between a specific animal pair (parent cattle and calf) within the same farm has been infrequently reported; most previous studies have instead described low-SNP clusters involving multiple animals and/or humans within a shared farm or environmental setting (Pietsch et al., 2018; Peng et al., 2022; Bachmann et al., 2024).
The seven CTX-M-14–producing isolates encompassed five distinct STs (ST1431, ST533, ST1148, ST448, and ST1261), reflecting genetic diversity within the farm. The three isolates from calf 3 exhibited different STs, indicating multiple circulating lineages. Among these, ST533 has been frequently detected in livestock from both Asia and Europe, suggesting adaptation to animal hosts (Huber et al., 2013; Dahms et al., 2015). Conversely, ST448 has been detected in both humans and animals, highlighting its potential to cross host boundaries and disseminate between humans and animals (Blaak et al., 2014; Quiñones et al., 2020; Sivarajan et al., 2025).
The identification of clonally identical ST533 isolates between the parent cattle and calf 1, supported by both SNP and cgMLST data, strongly indicates recent transmission and local clonal expansion within the livestock population. However, the ST448 isolates from calf 3 and the farmer differed by more than 3,800 core SNPs. These findings underscore the utility of high-resolution genomic analysis in detecting direct animal-to-human transmission, which may be difficult to infer from conventional typing methods. Thus, chromosomal SNP analysis demonstrated that the clonal spread of blaCTX-M-14-positive E. coli occurs independently of plasmid transfer, emphasizing the dual mechanisms whereby resistance persists and disseminates on farms. These relationships are schematically summarized in Figure 3.
Summary of clonal and plasmid-mediated dissemination of blaCTX-M-14–positive E. coli on a cattle farm. Schematic summary of genomic relationships among blaCTX-M-14–positive E. coli isolates from cattle and a farmer in 2013. Core-genome SNP analysis identified a clonally identical ST533 pair from a parent cattle and its calf (0–1 SNP difference), consistent with recent clonal transmission. In contrast, distinct E. coli lineages harbored highly conserved IncI1 blaCTX-M-14–encoding plasmids (>99.9% sequence identity), suggesting dissemination of closely related plasmids across hosts and bacterial backgrounds. High conjugative transferability supports the potential role of these plasmids in ESBL spread at the livestock–human interface.
Dissemination of identical CTX-M-14 plasmids
4.3
The transfer frequencies of IncI1 plasmids have been reported at levels of 10^−2^–10^−3^ per donor cell, which is considerably higher than those of many other plasmid types. Additionally, they stably persist without imposing a significant fitness cost on the host bacterium (Carattoli, 2009; Rozwandowicz et al., 2018). These properties facilitate the long-term maintenance and interspecies transfer of blaCTX-M-14.
Notably, we found that the 99.9% similarity between the plasmids detected in this study and those isolated in Japan further underscores the close genetic relationship between plasmids circulating in human and livestock populations. Therefore, IncI1 plasmids act as a “mobile resistome,” linking bacterial communities across different reservoirs. Similar observations have been reported in other countries, where IncI1 plasmids carrying blaCTX-M-14 were found to disseminate across diverse Enterobacterales lineages and between human and animal hosts (Liao et al., 2015; Yu et al., 2024). Our results reinforce the fact that plasmid-mediated dissemination is a major driver of ESBL spread in both clinical and agricultural settings.
Additional resistance determinants of concern
4.4
In addition, some isolates carried other clinically important resistance genes. One calf-derived isolate harbored aac(3)-IIa (gentamicin resistance), and another carried mcr-3.1 (colistin resistance). The presence of mcr genes in livestock is concerning, as colistin is a last-resort antimicrobial for multidrug-resistant Gram-negative infections (Liu et al., 2016; Yin et al., 2017; Wang et al., 2018). Tet(A) and aph(3′) genes, conferring resistance to tetracyclines and kanamycin, respectively, were also detected, reflecting selective pressure from long-standing antimicrobial use in livestock (Van Boeckel et al., 2019). The coexistence of blaCTX-M-14 with additional resistance determinants on separate mobile elements may facilitate co-selection and persistence of multidrug resistance. These findings underscore the public health risk of accumulating multiple resistance genes in livestock-associated E. coli and highlight the need for ongoing genomic surveillance within a One Health framework.
Limitations and implications
4.5
This study has some limitations. First, the sampling was restricted to a single farm with a limited number of cattle and a single participant, which may not represent broader epidemiological trends. Second, although clonal and plasmid sharing were observed, the precise transmission direction (human-to-animal, animal-to-human, or environment-mediated) could not be determined. Third, longitudinal and environmental sampling was not performed, preventing assessment of temporal dynamics and external sources. Fourth, detailed epidemiological metadata such as the animal age, origin, housing conditions, or duration of cohabitation are lacking from data collected in this study, limiting our ability to fully reconstruct transmission pathways or exclude environmental or external sources. Finally, we did not investigate the detailed structural plasticity of the plasmids, such as transposable elements or recombination events.
Conclusion
5
The identification of nearly identical plasmids in both cattle and human hosts suggests that plasmid-mediated ESBL resistance traverses host boundaries in agricultural settings, reinforcing the importance of a One Health perspective in surveillance. Additionally, the similarity to plasmids found in clinical isolates indicates that livestock may act as microbial reservoirs, contributing to the wider dissemination of resistance determinants in human populations. Finally, the high transferability of IncI1 plasmids encoding blaCTX-M-14 further underlines the potential for rapid spread under antibiotic selection pressure.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aldea I. Gibello A. Hernández M. Leekitcharoenphon P. Bortolaia V. Moreno M. A. (2022). Clonal and plasmid-mediated flow of ESBL/Amp C genes in Escherichia coli in a commercial laying hen farm. Vet. Microbiol. 270:109453. doi: 10.1016/j.vetmic.2022.10945335640410 · doi ↗ · pubmed ↗
- 2Altschul S. F. Gish W. Miller W. Myers E. W. Lipman D. J. (1990). Basic local alignment search tool. J Mol Biol. 215, 403–410. doi: 10.1016/S 0022-2836(05)80360-22231712 · doi ↗ · pubmed ↗
- 3Bachmann L. Weber L. Liermann W. Hammon H. M. Delling C. Dengler F. . (2024). Colostrum as a source of ESBL-Escherichia coli in feces of newborn calves. Sci. Rep. 14:9929. doi: 10.1038/s 41598-024-60461-4, 38688984 PMC 11061148 · doi ↗ · pubmed ↗
- 4Bevan E. R. Jones A. M. Hawkey P. M. (2017). Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 72, 2145–2155. doi: 10.1093/jac/dkx 146, 28541467 · doi ↗ · pubmed ↗
- 5Beyrouthy R. Sabença C. Robin F. Poeta P. Igrejas G. Bonnet R. (2021). Successful dissemination of plasmid-mediated extended-spectrum β-lactamases in Enterobacterales over humans to wild fauna. Microorganisms. 9:1471. doi: 10.3390/microorganisms 9071471, 34361907 PMC 8305760 · doi ↗ · pubmed ↗
- 6Blaak H. Hamidjaja R. A. van Hoek A. H. A. M. de Heer L. de Roda Husman A. M. Schets F. M. (2014). Detection of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli on flies at poultry farms. Appl. Environ. Microbiol. 80, 239–246. doi: 10.1128/AEM.02616-13, 24162567 PMC 3910986 · doi ↗ · pubmed ↗
- 7Bonnet R. (2004). Growing group of extended-spectrum beta-lactamases: the CTX-M enzymes. Antimicrob. Agents Chemother. 48, 1–14. doi: 10.1128/AAC.48.1.1-14.2004, 14693512 PMC 310187 · doi ↗ · pubmed ↗
- 8Bush K. Bradford P. A. (2020). Epidemiology of β-lactamase-producing pathogens. Clin. Microbiol. Rev. 33, e 00047–e 00019. doi: 10.1128/CMR.00047-19, 32102899 PMC 7048014 · doi ↗ · pubmed ↗