Systems-level molecular and immunological evidence identifies Th17/Treg modulation as a key mechanism of CRSJ’s neuroprotection in Parkinson’s disease
Xun Li, XiYu Li, ShiYa Chen, Lin Wang, JinYan Xia, MeiLing Zheng, ChuTian Zhang, XiaoQian Chen, Jing Cai

TL;DR
This study shows that Congrong Shujing Granules (CRSJ) protects against Parkinson's disease by balancing Th17 and Treg immune cells and reducing neuroinflammation.
Contribution
The study identifies Th17/Treg modulation as a novel immunomodulatory mechanism of CRSJ in Parkinson’s disease.
Findings
CRSJ treatment improves motor performance and preserves dopaminergic neurons in a Parkinson’s mouse model.
CRSJ modulates Treg and Th17 immune pathways and suppresses CX3CL1/CX3CR1–Th17 signaling.
CRSJ promotes microglial polarization and reduces α-synuclein accumulation and neuroinflammation.
Abstract
Parkinson’s disease (PD) is a progressive neurodegenerative disorder in which neuroinflammation plays a central role. Congrong Shujing Granules (CRSJ), a traditional Chinese medicine formula, have shown clinical benefits in PD, yet their immunomodulatory mechanisms remain unclear. We investigated the effects of CRSJ on Th17/Treg immune balance. Liquid chromatography–tandem mass spectrometry (LC-MS/MS) was used to identify representative chemical constituents of CRSJ. Representative CRSJ compounds were characterized, and their binding affinities were evaluated by molecular docking and molecular dynamics simulations. An MPTP-induced PD mouse model was established and treated with CRSJ. Behavioral outcomes, dopaminergic neuroprotection, immune cell subsets, transcriptomic profiles, and cytokine networks were assessed using flow cytometry, RNA sequencing, multiplex assays,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
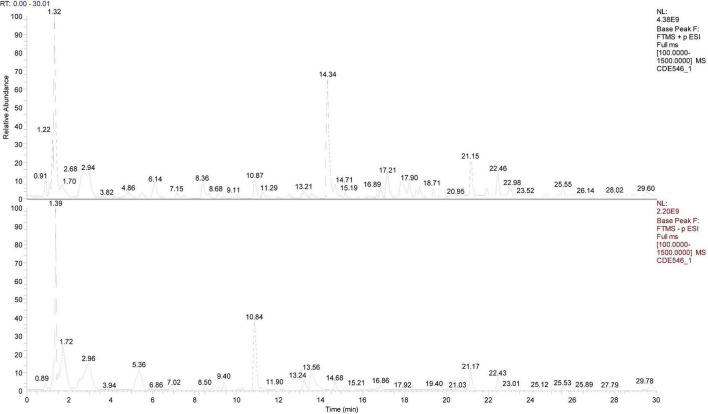 FIGURE 1
FIGURE 1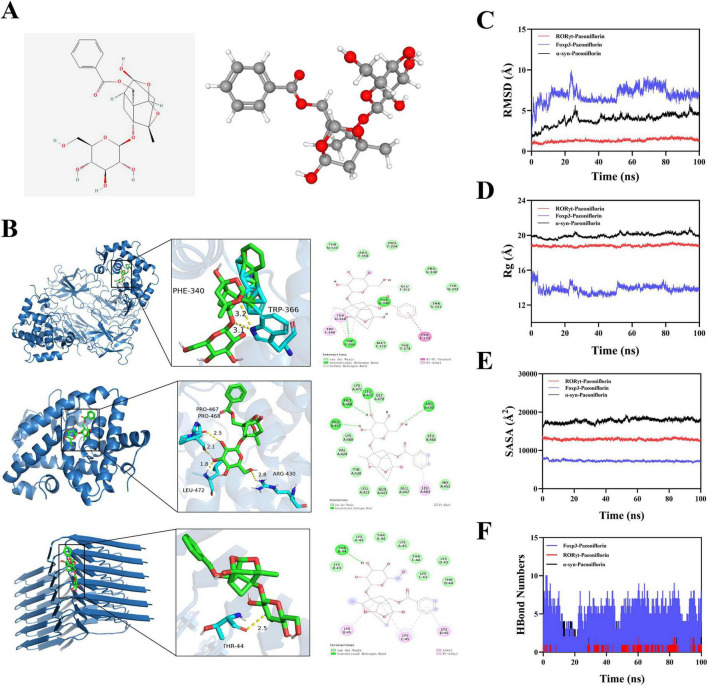 FIGURE 2
FIGURE 2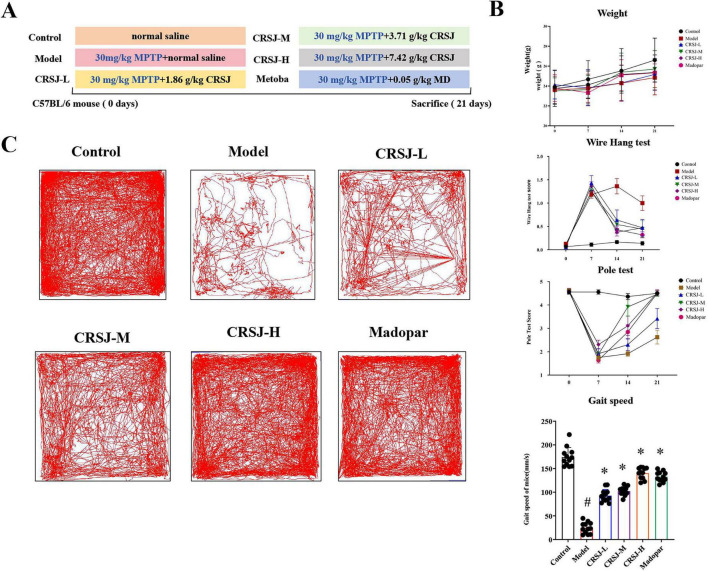 FIGURE 3
FIGURE 3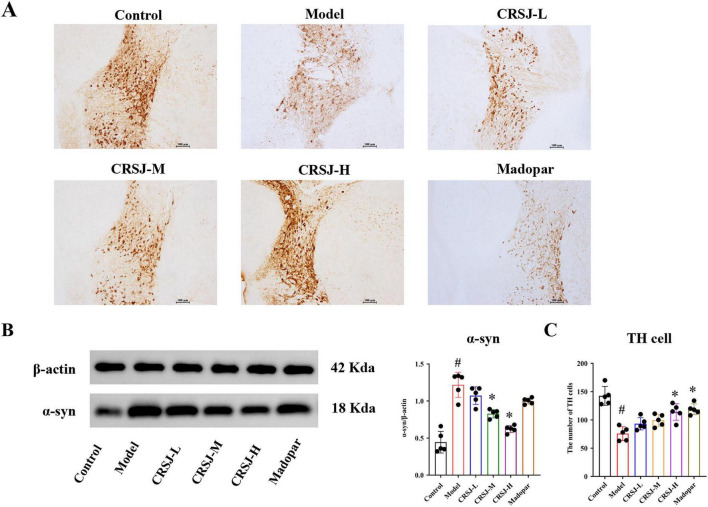 FIGURE 4
FIGURE 4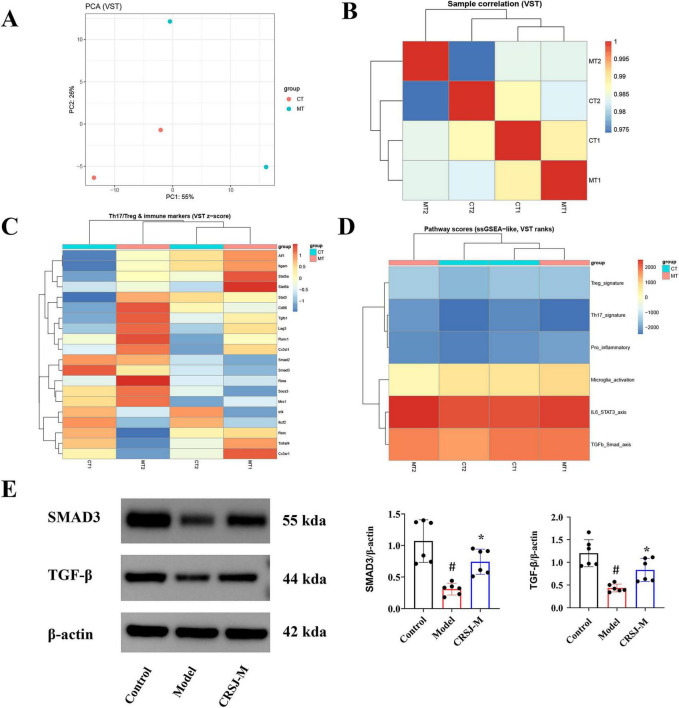 FIGURE 5
FIGURE 5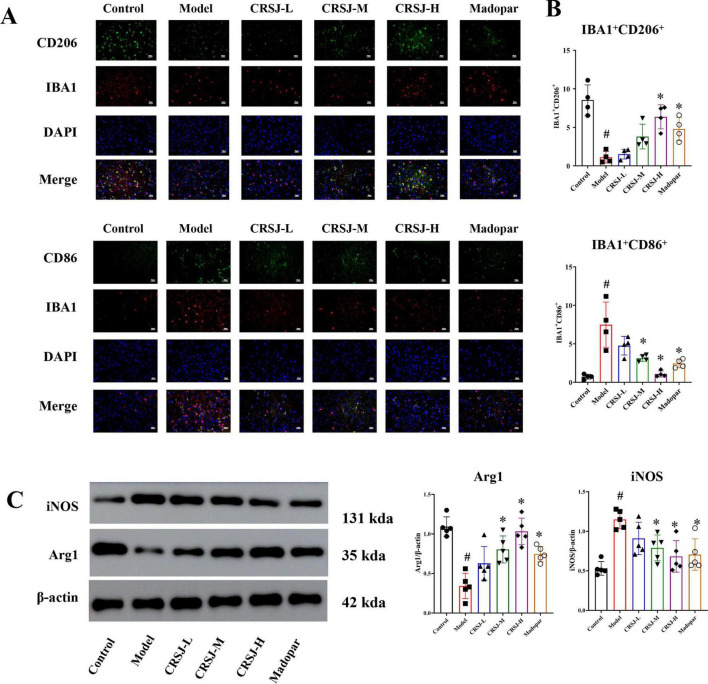 FIGURE 6
FIGURE 6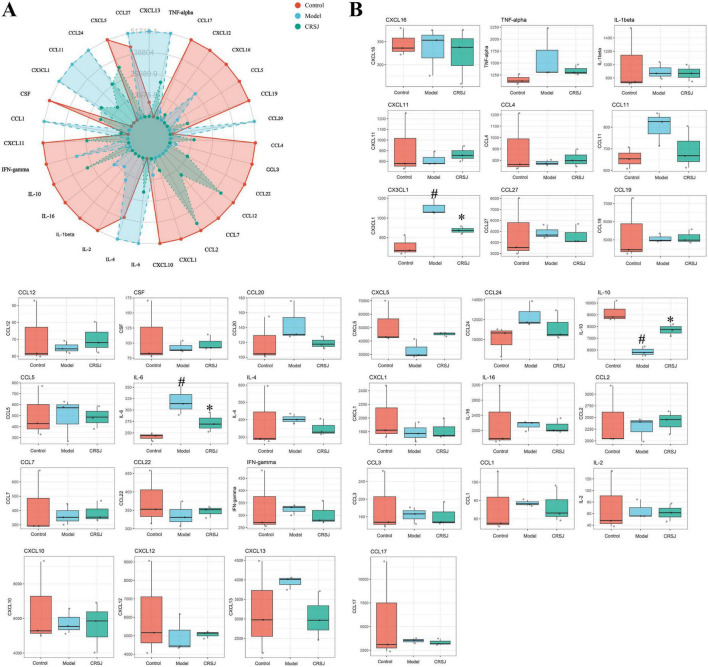 FIGURE 7
FIGURE 7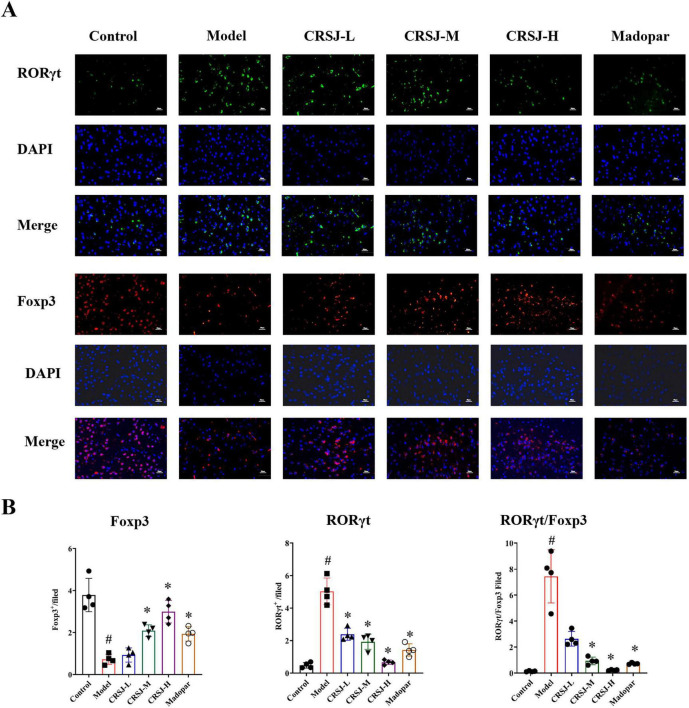 FIGURE 8
FIGURE 8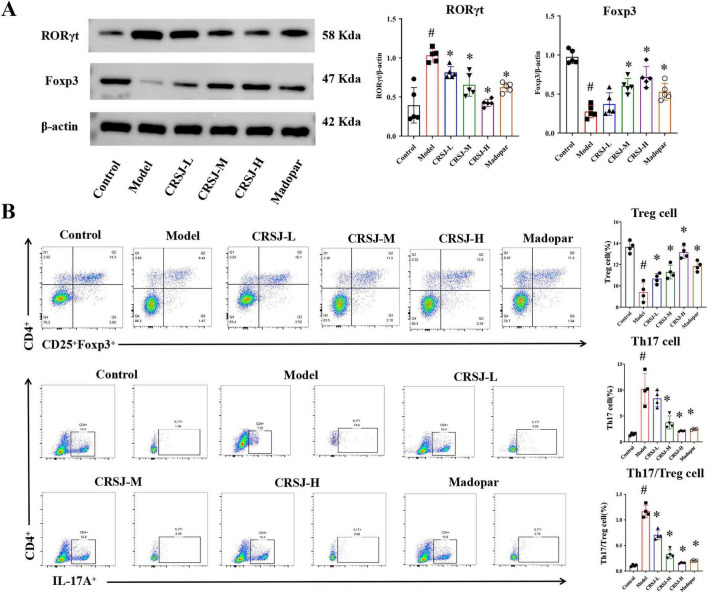 FIGURE 9
FIGURE 9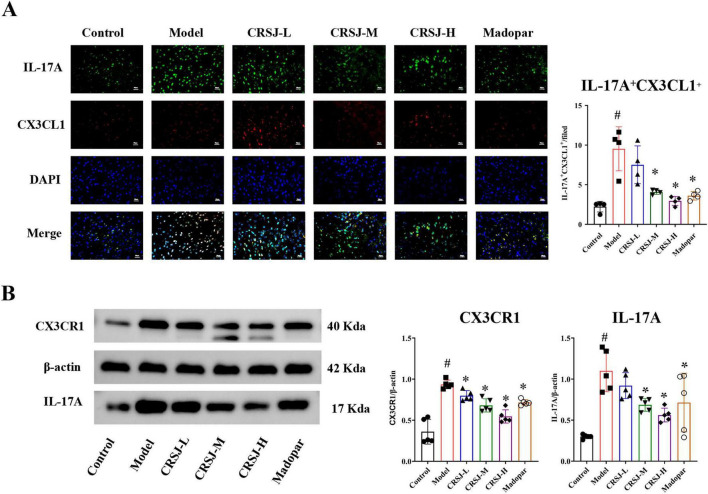 FIGURE 10
FIGURE 10| Herb | Identified compound | Chemical class | RT (min) | Representative biological relevance |
|---|---|---|---|---|
| Cistanche deserticola | Betaine | Alkaloid/osmolyte | 1.67 | Anti-inflammatory, methyl donor, immune modulation |
| Cistanche deserticola | Verbascoside | Phenylethanoid glycoside | 11.906 | Anti-inflammatory, antioxidant, Th17/Treg regulation |
| Cistanche deserticola | Tubuloside A | Phenylethanoid glycoside | 11.657 | Neuroprotection, anti-oxidative stress |
| Cistanche deserticola | Aucubin | Iridoid glycoside | 6.232 | Anti-inflammatory, neuroprotective |
| Cistanche deserticola | Tyrosol | Phenolic alcohol | 10.169 | Antioxidant, mitochondrial protection |
| Cistanche deserticola | Caffeic acid | Phenolic acid | 10.264 | Anti-inflammatory, ROS scavenging |
| Cistanche deserticola | Ferulic acid | Phenolic acid | 11.903 | Neuroprotection, anti-oxidative stress |
| Cistanche deserticola | Verbenalin | Iridoid glycoside | 10.473 | Anti-inflammatory, immune regulation |
| Salvia miltiorrhiza | Danshensu | Phenolic acid | 7.031 | Anti-inflammatory, microcirculatory protection |
| Salvia miltiorrhiza | Salvianolic acid A | Polyphenolic acid | 14.069 | Anti-neuroinflammation, antioxidant |
| Salvia miltiorrhiza | Salvianolic acid B | Polyphenolic acid | 13.596 | Neuroprotection, α-syn aggregation inhibition |
| Salvia miltiorrhiza | Lithospermic acid | Polyphenolic acid | 13.137 | Anti-inflammatory, vascular protection |
| Salvia miltiorrhiza | Cryptotanshinone | Diterpenoid quinone | 18.726 | Immunomodulation, Th17 inhibition |
| Salvia miltiorrhiza | Tanshinone IIA | Diterpenoid quinone | 19.66 | Anti-neuroinflammation, microglial regulation |
| Salvia miltiorrhiza | Dihydrotanshinone I | Diterpenoid quinone | 17.661 | Anti-inflammatory, neuroprotective |
| Salvia miltiorrhiza | Rosmarinic acid | Polyphenolic acid | 13.231 | Antioxidant, immune modulation |
| Paeonia lactiflora | Paeoniflorin | Monoterpene glycoside | 10.865 | Th17/Treg rebalancing, neuroimmune regulation |
| Paeonia lactiflora | Albiflorin | Monoterpene glycoside | 10.086 | Anti-inflammatory, neuroprotection |
| Paeonia lactiflora | Oxypaeoniflorin | Monoterpene glycoside | 9.401 | Immunomodulation |
| Paeonia lactiflora | Benzoylpaeoniflorin | Monoterpene glycoside | 14.699 | Anti-inflammatory |
| Paeonia lactiflora | Methyl gallate | Phenolic ester | 8.88 | Antioxidant, anti-inflammatory |
| Moutan ortex/Paeonia lactiflora | Gallic acid | Phenolic acid | 5.361 | Antioxidant, immune regulation |
| Moutan cortex | Paeonol | Phenolic ketone | 14.334 | Anti-neuroinflammation, microglial inhibition |
| Moutan cortex | Ellagic acid | Polyphenol | 13.823 | Anti-inflammatory, antioxidant |
| Polygonatum sibiricum | Genistein | Isoflavone | 18.761 | Th17 inhibition, Treg promotion |
| Polygonatum sibiricum | Hesperidin | Flavonoid glycoside | 12.91 | Anti-inflammatory, neuroprotection |
| Polygonatum sibiricum | Stachyose | Oligosaccharide | 1.482 | Gut–immune–brain axis modulation |
| Polygonatum sibiricum | Raffinose | Oligosaccharide | 1.505 | Immune homeostasis, prebiotic effect |
| Polygonatum sibiricum | Nystose | Oligosaccharide | 2.231 | Treg induction, immune tolerance |
| Polygonatum sibiricum | Isomangiferin | Xanthone glycoside | 1.83 | Antioxidant, neuroprotective |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParkinson's Disease Mechanisms and Treatments · Nuclear Receptors and Signaling · Ginseng Biological Effects and Applications
Introduction
1
Parkinson’s disease (PD) is a prevalent neurodegenerative disorder primarily characterized by motor impairments such as resting tremor, muscular rigidity, bradykinesia, and postural instability. With the global population aging, the incidence of PD is steadily increasing, thereby imposing a growing social and economic burden and presenting a significant public health challenge worldwide (Su et al., 2025). Although the precise pathogenic mechanisms of PD are not yet fully elucidated, accumulating evidence suggests that neuroinflammation and immune dysregulation play significant roles in the disease’s onset and progression (Lind-Holm Mogensen et al., 2025).
Over the past decade, converging evidence has established a central role for T-cell dysregulation in PD. Postmortem studies reveal infiltration of CD4^+^ and CD8^+^ T lymphocytes into the PD brain, implicating adaptive immunity in neurodegeneration (Sun et al., 2024). Among CD4^+^ subsets, Th17 cells disrupt blood–brain barrier integrity through IL-17–mediated tight-junction breakdown, thereby promoting the entry of inflammatory mediators and autoreactive immune cells into the central nervous system (Liu et al., 2019). Patients with sporadic PD consistently exhibit increased frequencies of IL-17–producing CD4^+^ T cells in peripheral blood (Sommer et al., 2019), and α-synuclein–activated Th17 cells can directly induce dopaminergic neuron loss in experimental models (Clarke et al., 2025). By contrast, regulatory T cells (Tregs) restrain effector T-cell activity and maintain immune homeostasis (Wang L. et al., 2025). In MPTP-induced PD models, adoptive Treg transfer mitigates dopaminergic neurodegeneration and microglial activation while enhancing neurotrophic factor expression (Reynolds et al., 2007). Tregs can also cross the blood–brain barrier and acquire memory-like phenotypes within the brain parenchyma, where they dampen local inflammation and confer neuroprotection (Machhi et al., 2020). Together, these findings highlight the Th17/Treg axis as a mechanistic driver of PD pathogenesis and a promising target for therapeutic intervention (Chen et al., 2015; McGinley et al., 2020).
Traditional Chinese Medicine (TCM), with its multi-component and systems-level therapeutic framework, has shown growing potential in neuroprotection and is considered a promising source of novel interventions for PD (Hao et al., 2024; Li et al., 2024; Zhang et al., 2014). CRSJ, a classical TCM formula composed of Cistanche deserticola, processed Polygonatum, Salvia miltiorrhiza, Paeonia lactiflora, and Moutan cortex, is clinically used to alleviate PD symptoms (Chen S. Y. et al., 2020). In TCM, CRSJ is thought to tonify kidney function, promote blood circulation, resolve stasis, and clear internal heat. Pharmacological studies indicate that CRSJ enhances dopaminergic neuronal survival via Wnt/β-catenin–mediated upregulation of tyrosine hydroxylase (TH) (Xu et al., 2021). Its medicated serum contains major bioactive constituents—including echinacoside, paeoniflorin, salvianolic acid B, acteoside, and tanshinone IIa—suggesting synergistic anti-inflammatory actions (Zhang et al., 2025; Supplementary Table 1). Among them, paeoniflorin is well documented to modulate the Th17/Treg balance by suppressing Th17-related factors (RORγt, IL-17, IL-6) and promoting Treg markers (Foxp3, IL-10, TGF-β) (Wang C. et al., 2025). These findings support the potential immunomodulatory role of CRSJ in PD, although its precise therapeutic mechanisms remain to be fully elucidated.
This study investigates the neuroprotective effects of CRSJ in MPTP-induced PD mice, focusing on its capacity to modulate the Th17/Treg immune axis. By delineating how CRSJ influences the dynamic balance between Th17 and Treg cells, this work aims to provide mechanistic insight into the neuroimmune regulation exerted by traditional Chinese medicine formulas (Pu et al., 2025; Tian et al., 2025).
Materials and methods
2
Experimental drugs
2.1
The CRSJ formulation consisted of Cistanche deserticola (6 g), processed Polygonatum (12 g), Salvia miltiorrhiza (15 g), Paeonia lactiflora (12 g), and Moutan cortex (10 g) (Supplementary Table 2). All herbal materials were supplied by the Third Affiliated Hospital of Fujian University of Traditional Chinese Medicine. Each component was decocted twice in water for 1–2 h, and the combined filtrates were concentrated at 50–85°C, dried, and homogenized to obtain the crude extract. Extracts were mixed according to the prescribed ratios and processed into 12–40 mesh granules by dry granulation. Based on body surface area conversion (factor = 9.1), the mouse-equivalent dose for a 20 g animal was calculated as 3.71 g/kg. Three CRSJ dosage groups were therefore established: low (1.68 g/kg), medium (3.71 g/kg), and high (7.42 g/kg), corresponding to 0.5 × , 1 × , and 2 × the adult equivalent dose.
Liquid chromatography–tandem mass spectrometry analysis
2.2
A precisely weighed 0.2 g aliquot of CRSJ powder was extracted with 10 mL of 80% methanol containing grinding beads. After grinding for 5 min and vortexing for 10 min, the mixture was centrifuged at 13,000 rpm for 10 min, and the supernatant was collected for analysis. Mass spectrometry was performed on a Q Exactive instrument (Thermo Fisher Scientific, Shanghai, China) using electrospray ionization (ESI) in both positive and negative switching modes. Chromatographic separation employed an UltiMate 3000 RS HPLC system (Thermo Fisher Scientific, Shanghai, China) with an AQ-C18 column (150 × 2.1 mm, 1.8 μm; Welch) at a flow rate of 0.7 mL/min, using water with 0.1% formic acid as the mobile phase. The gradient elution program is provided in Supplementary Table 5. High-resolution spectra were preprocessed with Compound Discoverer 3.3 (Thermo Fisher Scientific) and searched against the mzCloud database. Compounds scoring above the threshold were initially selected and subsequently confirmed by MS^2^ and MS^2^ spectral matching.
Molecular dock
2.3
Two-dimensional ligand structures were retrieved from PubChem, and their three-dimensional conformations were generated in ChemOffice and saved as mol2 files. High-resolution crystal structures of target proteins were obtained from the RCSB PDB. All protein structures were prepared in PyMOL by removing water molecules and other heteroatoms. Docking was performed using AutoDock Vina (version 1.5.6). Protein and ligand structures were processed by adding hydrogens, assigning charges, and defining rotatable bonds. Grid box coordinates were set according to the predicted binding pocket. The best binding pose was selected based on docking affinity scores. Protein–ligand interactions were visualized using Discovery Studio 2019. Binding affinity < −5.0 kcal/mol was considered indicative of good binding, whereas < −7.0 kcal/mol indicated strong binding. Lower binding energy reflects higher affinity and greater conformational stability.
Molecular dynamics
2.4
Molecular dynamics (MD) simulations were performed using GROMACS 2022 to evaluate the stability of protein–ligand complexes. The CHARMM36 force field was applied to proteins, and ligand parameters were generated using CGenFF. Complexes were solvated in a TIP3P water box with a 1.2 nm margin and neutralized with counterions. Long-range electrostatics were treated with the particle-mesh Ewald (PME) method, and the Verlet cutoff scheme was used. After energy minimization, NVT and NPT equilibration were performed for 200 ps each with position restraints on protein heavy atoms. Production MD simulations were run for 100 ns at 310 K and 1 bar. RMSD and RMSF analyses were performed to assess structural stability and residue flexibility (Li X. et al., 2025).
Animal studies
2.5
Male C57BL/6J mice (8 weeks, 20 ± 2 g) were obtained from the Laboratory Animal Center of Fujian University of Traditional Chinese Medicine and housed in SPF conditions [license: SYXK (Min) 2023-0004]. All procedures were approved by the Institutional Animal Care and Use Committee (FJTCM IACUC 0024037). After 1-week acclimation, 88 mice received intraperitoneal injections of MPTP (30 mg/kg/day) for 7 days to induce PD-like symptoms, while 12 mice served as untreated controls. Based on behavioral scoring (Supplementary Table 3), 60 successfully modeled mice were randomized into five groups: model, CRSJ-L (low-dose Congrong Shujing Granules, 1.68 g/kg), CRSJ-M (medium-dose Congrong Shujing Granules, 3.71 g/kg), and CRSJ-H (high-dose Congrong Shujing Granules, 7.42 g/kg), and Madopar (0.05 g/kg). Treatments were administered once daily by oral gavage for 14 days. Body weight was recorded weekly, and doses were adjusted accordingly. At the study endpoint, mice were anesthetized with pentobarbital sodium (150 mg/kg), and striatum and spleen were collected for analysis.
Behavioral assessments
2.6
Mice were habituated to the testing room for 1 h before each assessment. Behavioral tests—including the wire hang test, pole test, and open-field test—were conducted on day 9 after MPTP induction and on days 7 and 14 of treatment. Detailed scoring criteria are provided in Supplementary Tables 4, 5. In the pole test, mice were placed head-up at the top of a gauze-wrapped wooden pole (50 × 1 cm) and the time to descend was recorded. In the open-field test, mice were placed in the center of a 40 × 40 × 25 cm arena divided into 16 squares, and locomotor activity was monitored for 5 min using the TOP SCAN Super Maze 3.0 system. The apparatus was cleaned with 75% ethanol between trials.
Immunohistochemistry
2.7
Brain tissues were collected from treated mice and fixed in 4% paraformaldehyde, followed by dehydration, clearing, paraffin embedding, and sectioning into 5 μm slices. After deparaffinization and rehydration, antigen retrieval was performed in citrate buffer (pH 6.0) at 95–100°C for 15 min. Sections were then blocked with 5% goat serum for 30 min at room temperature. Subsequently, they were incubated overnight at 4°C with an anti–tyrosine hydroxylase primary antibody (Proteintech, 25859-1-AP). The next day, sections were incubated for 1 h with an HRP-conjugated goat anti-mouse/rabbit IgG polymer secondary antibody (Boster, SA1020). DAB chromogenic development was carried out using a commercial kit (Boster, AR1022) for 1–3 min until optimal staining was achieved, followed by dehydration and mounting. Images were acquired under a light microscope at 100 × magnification. For quantification, 3–5 sections per mouse were analyzed, and three randomly selected fields per section were imaged. TH-positive neurons in the substantia nigra were quantified using ImageJ to obtain the mean neuronal count for each animal.
Flow cytometry
2.8
Fresh spleens were gently dissociated through a 70-μm cell strainer to obtain single-cell suspensions, followed by red blood cell lysis. Cells were stimulated with PMA (Elabscience, No. E-CK-A091) for 5 h at 37°C in the presence of a protein transport inhibitor. After stimulation, cells were fixed and permeabilized using the FoxP3/Transcription Factor Staining Buffer Kit (LianKe, No. IC001). For Treg staining, cells were incubated with FITC-conjugated anti-CD4 (Elabscience, No. E-AB-F1353C), APC-conjugated anti-CD25 (Elabscience, No. E-AB-F1102E), and PE-conjugated anti-Foxp3 (Elabscience, No. E-AB-F1238D). For Th17 staining, cells were labeled with FITC-anti-CD4 and APC-anti-IL-17A antibodies (Elabscience, No. E-AB-F1199E). After washing and filtration to remove bubbles and aggregates, samples were analyzed on an LSRFortessa™ flow cytometer (BD Biosciences, New Jersey, United States). Based on established gating strategies, CD4^+^CD25^+^Foxp3^+^ cells were defined as Tregs, and CD4^+^IL-17A^+^ cells were identified as Th17 cells.
Immunofluorescence
2.9
Paraffin-embedded brain tissue sections were deparaffinized, rehydrated, and subjected to antigen retrieval. Sections were permeabilized with 1% Triton X-100 (Servicebio, No. G1204), washed with 0.1% PBS-T, and blocked with 10% goat serum at 37°C for 30 min. Primary antibodies against Foxp3 (Servicebio, No. GB112325-50), CX3CL1 (Proteintech, No. 60339), RORγt (Affinity, No. DF3196), IBA1 (Servicebio, No. GB12105), CD206 (Servicebio, No. GB113497), CD86 (Servicebio, No. GB150054), and IL-17 (Servicebio, No. GB11110) were applied and incubated overnight at 4°C. The next day, fluorescently labeled secondary antibodies (Servicebio, No. GB22303, GB27301, or GB28301) were added and incubated for 1 h at room temperature in the dark. Slides were mounted using DAPI-containing mounting medium (Servicebio, No. G1401). Fluorescence images were captured under identical exposure settings using a fluorescence microscope, and positive signals were quantified using Fiji software.
Western blot
2.10
Brain tissue samples were lysed using a protein extraction buffer (Servicebio, No. G2002), and protein concentrations were determined using the BCA assay. Equal amounts of protein were separated by SDS-PAGE and transferred onto PVDF membranes (Servicebio, No. G6045). Membranes were blocked with 5% non-fat milk at room temperature for 1.5 h, followed by overnight incubation at 4°C with primary antibodies against RORγt (Affinity, No. DF3196), CX3CR1 (Proteintech, No.60339), α-synuclein (Servicebio, No. GB11773), Foxp3 (Servicebio, No. GB112325), IL-17A (No. GB11110), Arg1 (Servicebio, No. GB11285), iNOS (Servicebio, No. GB153965), TGF-β (Servicebio, No. GB111876), smad3 (Servicebio, No. GB150085), and β-actin (Servicebio, No. GB15003). Membranes were then incubated with appropriate secondary antibodies for 1 h (Servicebio, No. GB23303, GB23301). Protein bands were visualized using a Bio-Rad chemiluminescence imaging system, and relative expression levels were quantified using AIWBwell™ analysis software.
Luminex liquid suspension chip assay
2.11
Serum samples were fully thawed at room temperature and diluted 1:4 before loading into 96-well plates for multiplex cytokine analysis using the Luminex^®^ 200 system (Luminex Corporation, Texas, United States). A 31-plex cytokine/chemokine panel (Bio-Rad, Bio-Plex Pro™ Mouse Chemokine Panel 31-Plex, No. 12009159) was used according to the manufacturer’s instructions. Samples and standards were incubated with magnetic beads for 1.5 h at room temperature in the dark with gentle shaking, followed by a series of wash steps and incubation with detection antibodies and streptavidin–PE. All serum samples were measured in duplicate. Quantification was based on bead classification and fluorescence intensity, and data acquisition and analysis were performed using Bio-Plex Manager™ software.
RNA sequencing and transcriptomic analysis
2.12
Two mice were randomly selected from each group for collection of bilateral striatum tissues. For RNA-seq, tissues from each mouse were processed independently, with each sample treated as one biological replicate. Total RNA was extracted using the MJzol Animal RNA Isolation Kit (Majorbio) and further purified with the RNAClean XP Kit (Beckman Coulter) and RNase-Free DNase Set (QIAGEN). rRNA-depleted, strand-specific libraries were constructed from each sample.
Sequencing was performed on the Illumina NovaSeq 6000 platform to generate 150-bp paired-end reads, yielding approximately 30 million reads per sample. Raw reads were subjected to quality control and trimming using TrimGalore (v0.6.x) and seqtk (v1.0). Clean reads were aligned to the mouse reference genome (GRCm39) using HISAT2 (v2.2.x), and alignment files were processed with SAMtools (v1.9) and Sambamba (v0.6.4). Transcript quantification was performed using StringTie (v1.3.3b) to obtain gene-level expression matrices. Differentially expressed genes (DEGs) were identified using edgeR (v3.2.0) with thresholds of Q-value ≤ 0.05 and | log2 fold change| ≥ 1. To investigate Th17/Treg-related immunoregulatory programs, single-sample gene set enrichment–like analyses were performed using curated Th17/Treg, IL-6–STAT3, and TGFβ–SMAD pathway signatures. Pathway scores and immune marker expression patterns were visualized using heatmaps. Given the limited sample size, pathway-level trends and coordinated transcriptional patterns were emphasized.
Antibody
2.13
The antibodies used in the IHC, IF, FCM, and WB are listed in Supplementary Tables 6–8.
Statistical analysis
2.14
All data are presented as mean ± standard deviation (mean ± SD). Statistical analyses were conducted using IBM SPSS Statistics 26.0, and graphical representations were generated with GraphPad Prism 9.2. Data normality and homogeneity of variance were assessed using the Shapiro–Wilk test, respectively. For data meeting the assumptions of normal distribution and homogeneity of variance, one-way analysis of variance followed by Tukey’s post-hoc test was used for multiple group comparisons. When the assumption of homogeneity of variance was violated, the Games–Howell post-hoc test was applied. Non-normally distributed data were analyzed using non-parametric tests. The significance level was set at α, = 0.05, with p < 0.05 considered statistically significant.
Results
3
Multi-class bioactive compounds in CRSJ support coordinated regulation of immune balance, neuroinflammation, and oxidative stress
3.1
Liquid chromatography–tandem mass spectrometry (LC–MS/MS) analysis of the CRSJ aqueous extract identified 44 representative compounds derived from its constituent herbs (Figure 1 and Table 1). Based on prior pharmacological evidence, these compounds were functionally associated with immunomodulatory and neuroprotective activities through partially overlapping pathways. Specifically, paeoniflorin and its analogs (albiflorin and oxypaeoniflorin), together with verbascoside, genistein, and paeonol, have been linked to regulation of the Th17/Treg axis via suppression of RORγt/IL-17 signaling and enhancement of Foxp3- and IL-10–related pathways. Salvianolic acid A and B, rosmarinic acid, danshensu, tanshinone IIA, and cryptotanshinone have been associated with attenuation of microglial activation, oxidative stress, and α-synuclein aggregation (Table 1). Additional phenolic compounds, including tyrosol, caffeic acid, ferulic acid, gallic acid, ellagic acid, and isomangiferin, are implicated in mitochondrial and redox homeostasis (Table 1). Oligosaccharides such as stachyose, raffinose, and nystose have been reported to contribute to peripheral immune regulation and Treg induction (Table 1). Collectively, the identified compounds span multiple chemical classes and suggest a multi-component profile consistent with coordinated regulation of immune balance, neuroinflammation, and oxidative stress.
LC–MS/MS total ion chromatogram of CRSJ. Column 1 shows the total ion current in the positive ion mode, and column 2 shows the total ion current in the negative ion mode. CRSJ, Congrong Shujing Granules; LC–MS/MS, liquid chromatography-tandem mass spectrometry.
Molecular-level evidence supports paeoniflorin as a Th17/Treg immunomodulator hub modulating Th17/Treg balance among CRSJ-derived serum constituents
3.2
CRSJ-medicated serum contains multiple bioactive constituents, including echinacoside, paeoniflorin, salvianolic acid B, acteoside, and tanshinone IIA, consistent with a cooperative anti-inflammatory profile. Among these, paeoniflorin (PF) has been extensively implicated in Th17/Treg regulation across inflammatory and autoimmune models, primarily through suppression of Th17-associated signaling and enhancement of Treg-related pathways (Figures 2A). Based on this established immunomodulatory relevance, PF was prioritized for molecular docking and molecular dynamics analyses with Foxp3, RORγt, and α-synuclein. Paeoniflorin exhibited favorable binding affinities toward all three targets, with relatively stronger interactions observed for RORγt (Figures 2B and Supplementary Table 9). Stable protein–ligand associations were supported by hydrogen bonding and hydrophobic interactions, and molecular dynamics simulations confirmed conformational stability of the complexes, particularly for the RORγt–paeoniflorin interaction (Figures 2C–F). Rather than assuming uniform bioactivity across all detected constituents, we integrated serum exposure, prior pharmacological evidence, and pathway relevance to prioritize functionally plausible components. Within this framework, paeoniflorin emerges as a key immunoregulatory node linked to Th17/Treg modulation, while other constituents likely provide complementary anti-inflammatory, antioxidant, and neuroprotective support. This systems-level chemical architecture aligns with the multi-target pharmacology of traditional formulations and offers a mechanistic basis for the immuno-neuroprotective effects of CRSJ.
Molecular docking and molecular dynamics simulations of paeoniflorin–protein complexes. (A) Chemical structure and three-dimensional conformation of paeoniflorin. (B) Predicted binding poses of paeoniflorin within the active sites of Foxp3, RORγt, and α-synuclein, obtained by molecular docking analysis. (C) RMSD profiles of paeoniflorin–protein complexes during the MD simulation. (D) Rg, reflecting the compactness and structural stability of each complex. (E) SASA of the complexes throughout the simulation period, indicating dynamic changes in protein–ligand exposure to solvent. (F) Time-dependent hydrogen bond number between paeoniflorin and target proteins, used to evaluate binding stability. RMSD, Root mean square deviation; SASA, solvent-accessible surface area; Rg, Radius of Gyration.
CRSJ improves motor function and neuromuscular performance in MPTP-induced PD mice
3.3
To assess CRSJ’s therapeutic efficacy, we established an MPTP-induced PD mouse model and evaluated motor performance (Figure 3A). Baseline body weight and motor performance were comparable across all groups prior to MPTP administration (Figure 3B). Following MPTP induction, PD model mice exhibited significant impairments in body weight gain, neuromuscular strength, and motor coordination, as reflected by altered wire hang and pole test performance, confirming successful model establishment (Figures 3B,C). CRSJ treatment dose-dependently improved body weight and motor function after 1 and 2 weeks of intervention, with the mid- and high-dose groups showing the most pronounced recovery, comparable to Madopar. Although motor performance did not fully return to baseline, all CRSJ-treated groups performed significantly better than the untreated model group, indicating that CRSJ effectively ameliorates MPTP-induced motor dysfunction. Overall, these findings indicate that CRSJ effectively alleviates MPTP-induced motor dysfunction and enhances neuromuscular performance in PD mice.
*CRSJ ameliorates motor deficits in MPTP-induced PD mice. (A) Experimental design and timeline. (B) Body weight, hanging test scores, pole test performance, and gait velocity in the nesting test before and after CRSJ treatment (n = 12). (C) Summary of locomotor trajectories of mice in the open field test (n = 12). P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ-L, low-dose Congrong Shujing Granules; CRSJ-M, medium-dose Congrong Shujing Granules; and CRSJ-H, high-dose Congrong Shujing Granules.
CRSJ preserves dopaminergic neurons and reduces α-Syn accumulation in PD mice
3.4
To assess the neuroprotective effects of CRSJ, we examined dopaminergic neuron survival in the substantia nigra and α-syn protein expression in the striatum of MPTP-induced PD mice. TH immunohistochemistry revealed a marked reduction of TH-positive neurons in the model group (P < 0.05), confirming dopaminergic degeneration (Figures 4A,C). CRSJ treatment significantly preserved TH-positive cells in a dose-dependent manner, with mid- and high-dose groups showing the strongest protection, comparable to the Madopar control (Figure 4C). Western blot analysis further demonstrated elevated α-syn expression in the striatum of model mice relative to controls (P < 0.05), consistent with pathological aggregation. CRSJ intervention significantly reduced α-syn levels, particularly in the mid- and high-dose groups, whereas the low-dose group showed a non-significant downward trend (Figure 4B). Together, these findings indicate that CRSJ suppresses abnormal α-syn accumulation and preserves dopaminergic neurons, supporting its neuroprotective potential in PD.
*CRSJ alleviates neuropathological alterations in the substantia nigra and striatum of PD mice. (A) Tyrosine hydroxylase (TH) immunoreactivity in the substantia nigra (n = 5). (B) Western blot and quantitative analysis of α-synuclein expression in the striatum (n = 5). (C) Quantification of TH-positive neurons in the substantia nigra (n = 5). #P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ-L, low-dose Congrong Shujing Granules; CRSJ-M, medium-dose Congrong Shujing Granules; CRSJ-H, high-dose Congrong Shujing Granules.
Transcriptomic profiling reveals that CRSJ restores Th17/Treg balance associated with modulation of the TGF-β/SMAD3 signaling axis
3.5
To characterize the molecular correlates of CRSJ-mediated immunomodulation in Parkinson’s disease, transcriptomic profiling was performed on striatal tissues from PD mice treated with mid-dose CRSJ. Principal component analysis (PCA) of variance-stabilized expression values showed clear separation between the model (MT) and CRSJ-treated (CT) groups along the first principal component, accounting for 55% of total variance (Figure 5A). Consistently, sample correlation analysis demonstrated high intra-group similarity and distinct inter-group clustering, indicating treatment-associated transcriptional differences with minimal technical variability (Figure 5B). Analysis of Th17/Treg-related immune markers revealed a coordinated shift toward a regulatory transcriptional profile following CRSJ treatment. Treg-associated components, including Tgfb1, Smad2/3, and Stat5 family members, were relatively upregulated, whereas Th17-associated regulators (Rorc, Rora, Stat3) and immune activation markers were attenuated (Figure 5C), indicating restoration of Th17/Treg-associated transcriptional balance. Pathway-level assessment using single-sample GSEA–like scoring further demonstrated coordinated pathway activity shifts, including suppression of Th17-related programs and the IL-6–STAT3 axis, accompanied by enhancement of Treg-associated signatures and the TGF-β–SMAD signaling pathway, together with reduced pro-inflammatory and microglial activation signatures in CRSJ-treated mice (Figure 5D). Among these pathways, TGF-β signaling emerged as a consistently altered feature across multiple analytical layers. Consistent with the transcriptomic results, Western blot analysis confirmed increased protein expression of TGF-β and SMAD3 in the striatum of CRSJ-treated mice compared with the PD model group (P < 0.05) (Figure 5E). Collectively, these data indicate that CRSJ treatment is associated with restoration of Th17/Treg-related immune programs, accompanied by consistent modulation of the TGF-β/SMAD3 signaling axis.
*Exploratory transcriptomic analysis reveals CRSJ-associated Th17/Treg immune-related expression patterns. (A) PCA based on VST–normalized expression values. (B) Sample-to-sample correlation heatmap generated from VST-normalized expression data. (C) Hierarchical clustering heatmap of selected Th17/Treg-associated and immune regulatory genes based on VST z-score–normalized expression values. (D) Heatmap of ssGSEA-like pathway activity scores estimated from VST-ranked gene expression data. (E) Western blot validation and quantitative analysis of TGF-β signaling pathway–related proteins (n = 6). #P < 0.05 vs. model group; P < 0.05 vs. control group. CRSJ-M, medium-dose Congrong Shujing Granules; CT, control group; MT, model group. PCA, Principal component analysis; VST, variance-stabilized transformation; SsGSEA, Single-sample GSEA.
CRSJ shifts microglial polarization from a pro-inflammatory M1 phenotype toward an anti-inflammatory M2 state in PD mice
3.6
Given the established links between Th17/Treg imbalance, dysregulated TGF-β/SMAD3 signaling, and microglial activation, we next examined whether CRSJ modulates microglial M1/M2 polarization in a PD mouse model. Western blot analysis showed that PD model mice exhibited significantly increased expression of the M1 marker inducible nitric oxide synthase (iNOS) and reduced expression of the M2 marker arginase-1 (Arg1) compared with normal controls (P < 0.05) (Figure 6C). CRSJ treatment markedly reversed this polarization shift, with decreased iNOS and increased Arg1 levels across medium-, and high-dose groups (P < 0.05) (Figure 6C). Immunofluorescence double staining further confirmed these effects at the cellular level. The proportion of IBA1^+^CD86^+^ microglia was significantly increased in the model group, whereas IBA1^+^CD206^+^ cells were reduced (P < 0.05) (Figures 6A,B). CRSJ treatment significantly decreased IBA1^+^CD86^+^ co-expression in CRSJ-M and CRSJ-H, but not in the low-dose group (P > 0.05) (Figure 6A). In contrast, IBA1^+^CD206^+^ polarization was significantly enhanced only in the high-dose group, with no significant differences observed in the low- or medium-dose groups compared with the model group (P > 0.05) (Figures 6A,B). The Madopar group showed partial improvement. Together, these findings indicate that CRSJ shifts microglial polarization from a pro-inflammatory M1 phenotype toward an anti-inflammatory M2 state in the PD brain.
*CRSJ associated modulation of M1/M2 immune-related expression markers. (A) Immunofluorescent staining of IBA1+CD86+ and IBA1+CD206+ expression (n = 4). (B) quantification of IBA1+CD86+ and IBA1+CD206+ expression (n = 4). (C) Western blot and quantitative analysis of iNOS and Arg1 proteins (n = 5). #P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ-L, low-dose Congrong Shujing Granules; CRSJ-M, medium-dose Congrong Shujing Granules; CRSJ-H, high-dose Congrong Shujing Granules.
CRSJ attenuates cytokine and chemokine dysregulation in PD mice
3.7
Thirty one cytokines and chemokines was quantified to characterize systemic immune alterations. The Model group showed broad pro-inflammatory activation, with notable increases in neuroimmune chemokines (CX3CL1, CXCL10, CXCL12, CCL4) and interleukins such as IL-1β, IL-16, and IL-4, confirming heightened microglial and peripheral immune activity. CRSJ treatment partially normalized these abnormalities, reducing CX3CL1, CXCL10, CXCL12, TNF-α, IL-1β, and recruitment-related chemokines (CCL2, CCL3, CCL22), while markers such as CCL7, CCL12, and CCL20 remained largely unchanged (Figures 7A,B). Consistent with these trends, IL-6 was significantly elevated and IL-10 reduced in PD mice (P < 0.05), indicating Th17/Treg imbalance; CRSJ effectively reversed both changes (P < 0.05) (Figure 7B). Together, these results demonstrate that CRSJ attenuates PD-associated cytokine dysregulation and restores key neuroimmune regulatory pathways.
*Serum cytokine expression profiles. (A) Immuno-radar plot illustrating the expression levels of 31 serum cytokines across groups (n = 3). (B) Boxplots showing the distribution of individual cytokines (n = 3). #P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ, medium-dose Congrong Shujing Granules.
CRSJ restores Th17/Treg immune balance by modulating RORγt and Foxp3 expression
3.8
To assess CRSJ’s immunomodulatory effects, we examined Th17/Treg-associated transcription factors and cell populations. In PD mice, RORγt expression was increased and Foxp3 was decreased in the mesencephalic region, indicating Th17-skewed immune differentiation (Figure 8A). CRSJ treatment significantly reduced RORγt expression across all doses, whereas Foxp3 expression and the RORγt/Foxp3 ratio were significantly corrected only in the medium- and high-dose groups but not in the low-dose group compared with the model group (P > 0.05), with similar effects observed in the Madopar group (Figure 8B). Flow cytometry further confirmed systemic immune imbalance: model mice displayed increased Th17 (CD4^+^IL-17A^+^) and decreased Treg (CD4^+^CD25^+^Foxp3^+^) populations, resulting in a disrupted Th17/Treg ratio (Figure 9B). Mid- and high-dose CRSJ effectively normalized these proportions (P < 0.05). Correspondingly, Western blot analysis showed striatum of upregulated RORγt and downregulated Foxp3 in model mice, both of which were significantly corrected by CRSJ-M, CRSJ-H, and Madopar (Figure 9A). Together with cytokine profiling and immunofluorescence data, these results demonstrate that CRSJ restores Th17/Treg homeostasis by modulating transcriptional programs and T-cell differentiation at multiple regulatory levels.
*Th17/Treg transcription factors distribute. (A) Immunofluorescent staining of RORγt and Foxp3 expression (n = 4). (B) quantification of RORγt and Foxp3 expression (n = 4). #P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ-L, low-dose Congrong Shujing Granules; CRSJ-M, medium-dose Congrong Shujing Granules; CRSJ-H; high-dose Congrong Shujing Granules.
*Quantification of Th17 and Treg cells and their transcription factor expression. (A) Western blot and quantitative analysis of Foxp3 and RORγt proteins (n = 5). (B) Flow cytometric profiling and quantification of Th17 and Treg cell populations (n = 4). #P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ-L, low-dose Congrong Shujing Granules; CRSJ-M, medium-dose Congrong Shujing Granules; CRSJ-H, high-dose Congrong Shujing Granules.
CRSJ suppresses CX3CL1/CX3CR1 signaling and Th17 activation in PD mice
3.9
To examine whether CRSJ regulates chemokine CX3CL1 and Th17-cell–related immune responses in PD mice, we quantified CX3CL1 expression and key Th17-associated indicators. Among these, CX3CL1 was significantly elevated in the Model group compared with controls (P < 0.05), and was markedly reduced following CRSJ-M and CRSJ-H treatment (P < 0.05) (Figure 7B). To assess downstream mediators, striatal expression of CX3CR1—the cognate receptor of CX3CL1—and IL-17A, a key Th17 effector molecule, was examined. Western blotting revealed significant upregulation of both CX3CR1 and IL-17A in the Model group, which were significantly suppressed by CRSJ-M, CRSJ-H, and Madopar treatments (P < 0.05); IL-17A reductions in the CRSJ-L group did not reach statistical significance (P > 0.05) (Figure 10A). Dual immunofluorescence co-localization analysis further demonstrated increased CX3CL1–IL-17A co-expression in the Model group (P < 0.05), which was significantly attenuated by all CRSJ doses and Madopar (P < 0.05) (Figure 10B). Collectively, these results show that CRSJ effectively modulates CX3CL1/CX3CR1 signaling and Th17-related activity, thereby contributing to restoration of Th17/Treg balance and exerting neuroprotective effects in PD.
*Expression of CX3CL1 and Th17 Cells in PD mice. (A) Co-localization of CX3CL1 and IL-17A in immunofluorescence images, along with quantitative analysis and serum CX3CL1 levels (n = 4). (B) Western blot and quantitative analysis of CX3CL1 and IL-17A (n = 5). #P < 0.05 vs. Model group; P < 0.05 vs. Control group. CRSJ-L, low-dose Congrong Shujing Granules; CRSJ-M, medium-dose Congrong Shujing Granules; CRSJ-H, high-dose Congrong Shujing Granules.
Discussion
4
PD is the second most common neurodegenerative disorder worldwide, with a rapidly increasing prevalence driven by global population aging (Luo et al., 2024). PD is characterized by progressive dopaminergic neuron loss, reduced TH expression, and pathological α-syn aggregation, features faithfully reproduced in our MPTP-induced model (Nagatsu, 2024; Praveen et al., 2025). CRSJ significantly improved motor coordination and locomotor performance, indicating functional rescue at the behavioral level. Mechanistically, CRSJ preserved TH-positive neurons in the substantia nigra and reduced α-syn accumulation in the striatum. Such dual engagement of immune-regulatory pathways and protein aggregation processes supports the disease-modifying potential of CRSJ rather than merely symptomatic relief.
Th17 and Treg cells drive opposing immunological outcomes in neurodegeneration. Th17 cells, regulated by RORγt, secrete IL-17A and IL-6, disrupt blood–brain barrier integrity, and exacerbate dopaminergic neurotoxicity (Chen J. et al., 2020; Jiang et al., 2025; Liu et al., 2022; Shi et al., 2022; Zheng and Luo, 2025). Conversely, Foxp3^+^ Treg cells secrete IL-10 and TGF-β, attenuating microglial activation and promoting neuronal survival (Huang et al., 2020; Sun and Guo, 2025; Xie et al., 2024). Our data show that PD mice exhibit a pronounced Th17/Treg imbalance, reflected by increased Th17 cell numbers, elevated RORγt and IL-17A expression, and reduced Foxp3 and IL-10 signals—consistent with clinical and preclinical observations that Treg exhaustion and Th17 overactivation contribute to PD progression (Li et al., 2023). CRSJ reversed these abnormalities at transcriptional, protein, and cellular levels. By suppressing RORγt and restoring Foxp3, CRSJ shifts immune programming toward a regulatory phenotype, thereby reducing neuroinflammation and neuronal vulnerability. This supports the concept that Th17/Treg rebalancing is a crucial mechanism of CRSJ’s therapeutic action.
Transcriptomic analyses suggested that CRSJ treatment was associated with coordinated changes in immune- and neuroinflammation-related networks in the striatum, rather than widespread gene-level changes. Although few genes met stringent differential expression criteria, pathway- and module-level analyses consistently highlighted immunoregulatory signaling, with the TGF-β pathway emerging as a dominant feature. Single-sample GSEA–like pathway scoring and immune marker profiling further demonstrated suppression of Th17-associated transcriptional programs accompanied by enhancement of Treg-related signaling following CRSJ treatment. Collectively, these findings highlight the TGF-β/SMAD3 axis as a recurrently modulated pathway, characterized by upregulation of TGF-β/SMAD signaling components and concomitant attenuation of the IL-6–STAT3 pathway, a key driver of Th17 differentiation (Cheng et al., 2025; Park and Ciofani, 2025; Xu Lou et al., 2025). Consistent with these transcriptomic findings, increased TGF-β and SMAD3 protein expression was confirmed in the striatum, supporting a model in which CRSJ confers neuroprotection by selectively reinforcing Treg-mediated immunoregulatory networks while limiting Th17-driven neuroinflammation and microglial overactivation.
Accumulating evidence indicates that Th17/Treg imbalance is closely linked to maladaptive microglial activation in Parkinson’s disease (Roodveldt et al., 2024). Consistent with this framework, our data show that CRSJ shifts microglial polarization from a pro-inflammatory M1 phenotype toward an anti-inflammatory M2 state, as evidenced by reduced iNOS, increased Arg1 expression, and reciprocal changes in IBA1^+^CD86^+^ and IBA1^+^CD206^+^ microglia. Th17 effector signaling, particularly via the IL-17/IL-17R axis, promotes microglial activation and neuroinflammatory amplification (Li T. et al., 2025), whereas regulatory T cells constrain CNS innate immune responses (Jie et al., 2026). Together with emerging transcriptomic evidence that immune-intrinsic programs reshape Th17 differentiation, these findings support a model in which restoration of Th17/Treg balance is closely associated with normalization of microglial states (Sun et al., 2026). These cellular alterations parallel transcriptomic signatures of restored Th17/Treg balance and attenuation of Th17-associated inflammatory programs, suggesting that CRSJ modulates microglial activation primarily through upstream immune differentiation rather than direct inhibition of terminal inflammatory mediators. Although changes in TGF-β/SMAD3 signaling were observed, this pathway likely represents one component of a broader, multi-pathway immunoregulatory network. Accordingly, the neuroprotective effects of CRSJ are best interpreted as arising from integrated modulation of immune programs that collectively restrain chronic neuroinflammation and support dopaminergic neuron resilience.
CX3CL1 emerged as a pivotal chemokine altered in the PD model. Although CX3CL1 can exert context-dependent neuroprotective effects (Chu et al., 2025; Gutiérrez et al., 2025), many studies report early stage elevation and later decline during PD progression (Gutiérrez et al., 2025; Iemmolo et al., 2023; Li et al., 2022; Qu et al., 2023), reflecting dynamic microglial–neuronal interactions. Importantly, CX3CL1 is tightly linked to Th17 biology: IL-17A induces CX3CL1 expression, and CX3CL1/CX3CR1 signaling promotes Th17 chemotaxis and polarization (Akiyama et al., 2025; Yang et al., 2025). We observed significant increases in CX3CL1, CX3CR1, and Th17 markers in PD mice, supporting a cooperative inflammatory amplification loop (Li X. et al., 2025). CRSJ markedly suppressed CX3CL1/CX3CR1 expression and reduced IL-17A levels, indicating that CRSJ may disrupt this chemotactic–inflammatory circuit. By breaking this feedback loop, CRSJ limits Th17 recruitment and activation, thereby reducing inflammatory burden on vulnerable dopaminergic neurons. These findings highlight CX3CL1 as an immune–chemotactic target modulated by CRSJ and underscore its potential relevance in PD immunopathology.
CRSJ, as a traditional multi-herb formulation, exhibits therapeutic potential in PD. LC–MS analysis identified 44 representative compounds in the CRSJ aqueous extract spanning multiple chemical classes with established immunomodulatory and neuroprotective relevance. By integrating serum exposure, prior pharmacological evidence, and pathway involvement, we prioritized functionally plausible constituents rather than assuming uniform bioactivity. Recent studies support that paeoniflorin/total glucosides of peony and verbascoside-containing preparations can modulate Th17/Treg homeostasis (Zhao et al., 2025; Zheng et al., 2020), consistent with suppression of Th17-skewing inflammatory programs and enhancement of regulatory outputs (e.g., IL-10/Foxp3), while paeonol-containing regimens have likewise been reported to normalize Th17/Treg-related immune imbalance in inflammatory settings (Shi et al., 2021). In parallel, salvianolic acids (Akhtar et al., 2025; Bi et al., 2025), tanshinones (Zeng et al., 2024), and related phenolics have been linked to attenuation of microglial activation, oxidative stress, and α-synuclein pathology (Thi Lai et al., 2024; Yang et al., 2024). Consistent with this prioritization, molecular docking and molecular dynamics simulations identified paeoniflorin as a key immunoregulatory constituent, exhibiting stable binding to Foxp3, RORγt, and α-synuclein, with the strongest affinity toward RORγt. Collectively, these findings position paeoniflorin as a functionally prioritized immunoregulatory component, supported by complementary anti-inflammatory and neuroprotective constituents, providing a mechanistic basis for the multi-target immuno-neuroprotective effects of CRSJ.
While this study provides convergent evidence that CRSJ modulates Th17/Treg differentiation, suppresses chemokine-driven neuroinflammation, and attenuates α-syn pathology, several limitations should be acknowledged. First, although microglial polarization was systematically characterized, additional immune components, including B-cell subsets and α-syn–reactive antibody-producing cells, were not comprehensively examined and may represent further layers of CRSJ-mediated immunoregulation. Future work using multiparameter flow cytometry, CyTOF, or single-cell RNA sequencing will allow cell type–resolved mapping of CRSJ-induced immune responses in the central nervous system, together with detailed pharmacokinetic and bioavailability profiling of prioritized CRSJ constituents. Second, part of the immunological evidence was generated in an exploratory context, with transcriptomic analyses based on two biological replicates per group and flow cytometry performed in four mice per group. Although sufficient to identify consistent immune signatures and pathway-level alterations supported by independent validation, these sample sizes limit statistical power and sensitivity to inter-individual variability. Finally, reliance on a single MPTP-induced mouse model and the absence of human immune or clinical data constrain direct translational inference, warranting validation in additional PD models and human-derived samples.
In summary, this study identifies CRSJ as a conceptually distinct immunomodulatory strategy for Parkinson’s disease. Unlike existing approaches that target single inflammatory mediators or discrete immune cell subsets, CRSJ exerts coordinated, multi-target regulation across Th17/Treg differentiation, chemokine signaling, microglial activation, and α-syn–associated pathology. Such network-level immune reprogramming may be particularly advantageous in the multifactorial and dynamic inflammatory milieu of PD. Nevertheless, the exploratory nature of the transcriptomic analyses, limited sample sizes, reliance on a single toxin-induced model, and absence of human immune data constrain direct translational inference. Future studies incorporating larger cohorts, complementary disease models, and human-derived samples will be essential to validate and refine the therapeutic potential of CRSJ.
Conclusion
5
Collectively, this study provides convergent evidence that CRSJ treatment is associated with improved motor performance, preservation of dopaminergic neurons, and attenuation of α-synuclein accumulation in a Parkinson’s disease mouse model. These benefits are accompanied by coordinated modulation of immune-related processes, including restoration of Th17/Treg-associated immune balance, suppression of CX3CL1/CX3CR1–related inflammatory signaling, and consistent regulation of the TGF-β–SMAD3 pathway, with downstream normalization of microglial activation and overall reduction of neuroinflammation. Our findings support a multi-component, synergistic mode of action, in which paeoniflorin acts in concert with other active constituents to modulate interconnected immune, inflammatory, and proteostatic pathways. Although limited by sample size and the absence of human validation, this work provides a mechanistic foundation for the further development of CRSJ as a disease-modifying, immunoregulatory strategy for Parkinson’s disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akhtar A. Singh P. Admane N. Grover A. (2025). Salvianolic acid B prevents the amyloid transformation of A 53T mutant of α-synuclein. Biophys. Chem. 318:107379. 10.1016/j.bpc.2024.107379 39693815 · doi ↗ · pubmed ↗
- 2Akiyama M. Wakasugi S. Yoshimoto K. Saito K. Ishigaki S. Inukai R. (2025). CX 3CR 1+ age-associated CD 4+ T cells contribute to synovial inflammation in late-onset rheumatoid arthritis. Inflamm. Regen. 45:4. 10.1186/s 41232-025-00367-4 39910629 PMC 11800492 · doi ↗ · pubmed ↗
- 3Bi S. Liu S. Zhu K. Gao D. Chen L. Yu C. (2025). Preclinical and experimental evidence of salvianolic acid B in the treatment of neurological diseases. Front. Pharmacol. 16:1606146. 10.3389/fphar.2025.1606146 40657643 PMC 12245778 · doi ↗ · pubmed ↗
- 4Chen J. Liu X. Zhong Y. (2020). Interleukin-17A: The key cytokine in neurodegenerative diseases. Front. Aging Neurosci. 12:566922. 10.3389/fnagi.2020.566922 33132897 PMC 7550684 · doi ↗ · pubmed ↗
- 5Chen S. Y. Xiao S. J. Lin Y. N. Li X. Y. Xu Q. Yang S. S. (2020). Clinical efficacy and transcriptomic analysis of congrong shujing granules () in patients with Parkinson’s disease and syndrome of shen (Kidney) essence deficiency. Chin. J. Integr. Med. 26 412–419. 10.1007/s 11655-020-3080-0 32291608 · doi ↗ · pubmed ↗
- 6Chen Y. Qi B. Xu W. Ma B. Li L. Chen Q. (2015). Clinical correlation of peripheral CD 4+-cell sub-sets, their imbalance and Parkinson’s disease. Mol. Med. Rep. 12 6105–6111. 10.3892/mmr.2015.4136 26239429 · doi ↗ · pubmed ↗
- 7Cheng H. Nan F. Ji N. Ma X. Zhang J. Liang H. (2025). Regulatory T cell therapy promotes TGF-β and IL-6-dependent pro-inflammatory Th 17 cell generation by reducing IL-2. Nat. Commun. 16:7644. 10.1038/s 41467-025-62628-7 40818959 PMC 12357911 · doi ↗ · pubmed ↗
- 8Chu Y. Harms A. S. Boehringer A. Kordower J. H. (2025). Decreased neuronal and increased endothelial fractalkine expression are associated with neuroinflammation in Parkinson’s disease and related disorders. Front. Cell. Neurosci. 19:1557645. 10.3389/fncel.2025.1557645 40842561 PMC 12364955 · doi ↗ · pubmed ↗