Two Novel Trichoderma Species and their Antagonistic Activity against Sclerotia-producing Plant Pathogens
Jessica Rembinski, Lucas S. Sales, Phellippe A. S. Marbach, Jorge T. De Souza

TL;DR
This study discovers two new Trichoderma species and shows they can effectively control plant pathogens that produce sclerotia.
Contribution
The discovery of two novel Trichoderma species and their superior antagonistic activity against sclerotia-producing pathogens.
Findings
Two new Trichoderma species, T. variabile and T. littericola, were identified from the Restinga ecosystem.
T. nordicum was found to have a broader distribution and showed strong antagonistic activity against S. sclerotiorum and S. cepivora.
T. littericola and T. nordicum outperformed the commercial T. harzianum in antagonistic activity.
Abstract
Trichoderma species are among the most employed fungal biocontrol agents in global agriculture. One mechanism of activity of these fungi is mycoparasitism, which is responsible for degrading the cell wall of other fungi through the production of enzymes such as chitinases and cellulases. This mechanism is capable of targeting resistant structures like sclerotia and chlamydospores produced by pathogens such as Sclerotinia sclerotiorum and Stromatinia cepivora. Several Trichoderma species have been reported as antagonists of these pathogens, but most described species were not explored for their antagonism against sclerotia-producing pathogens. The objective of this study was to characterize new species of the Trichoderma genus and evaluate their antagonistic potential against pathogens that produce sclerotia. The taxonomical novelties reported in this study are the description of two…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
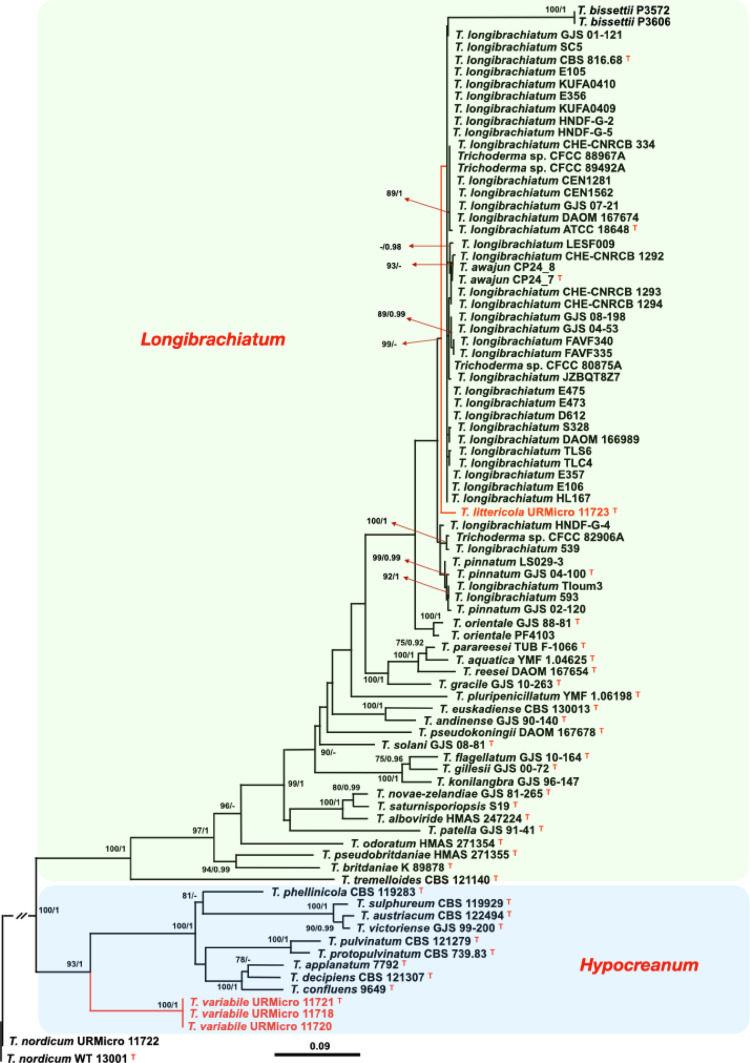 Figure 1
Figure 1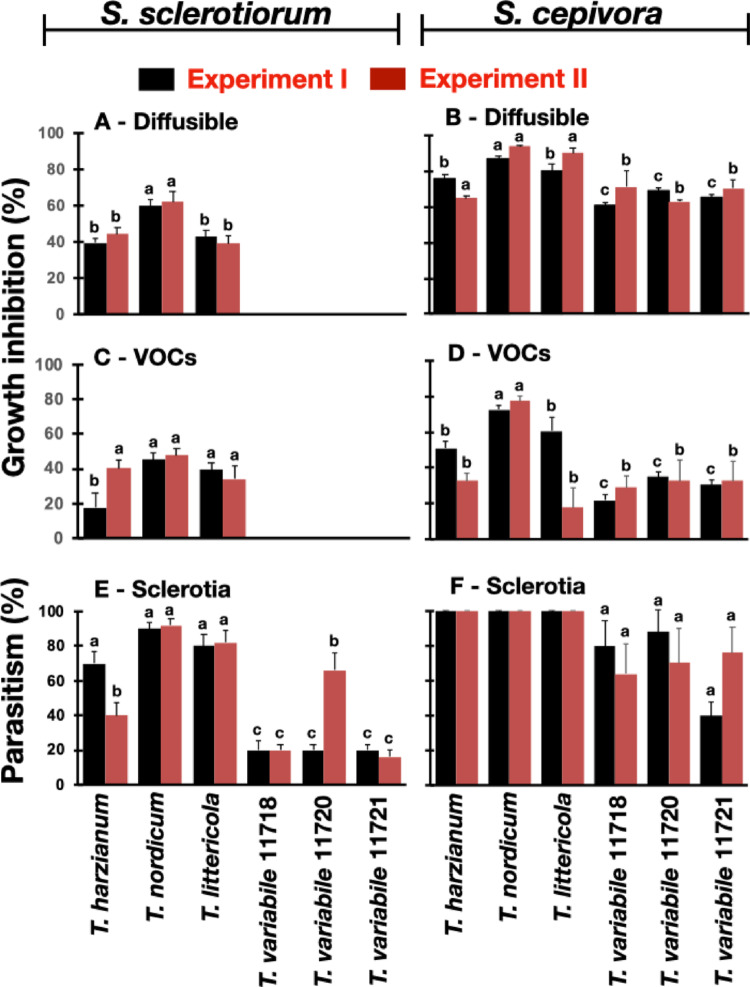 Figure 2
Figure 2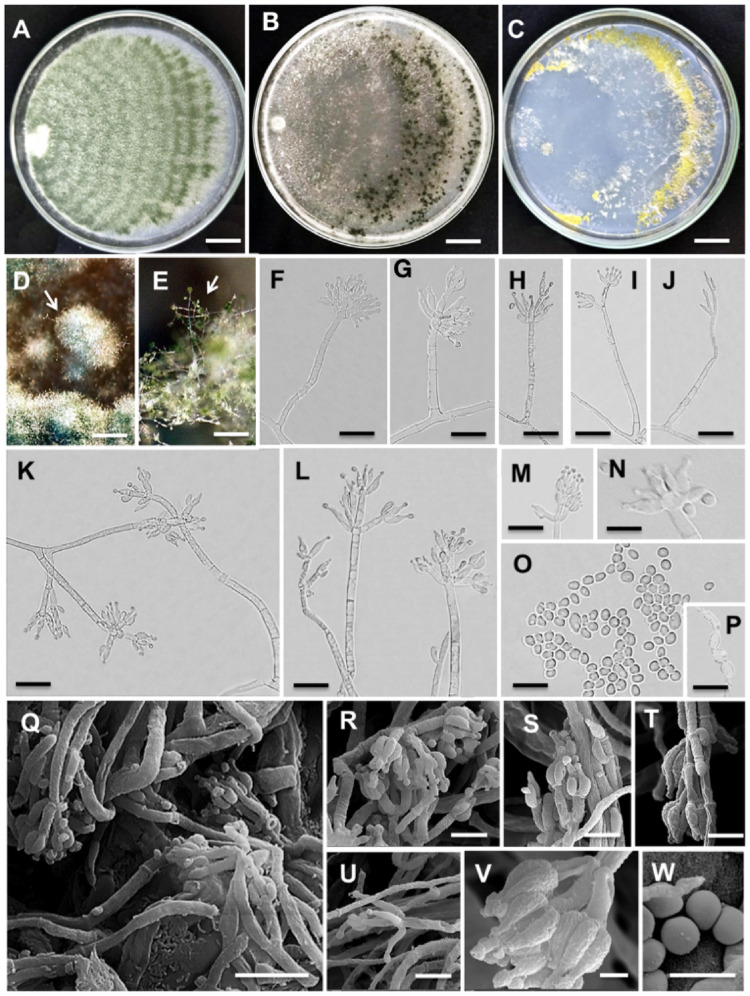 Figure 3
Figure 3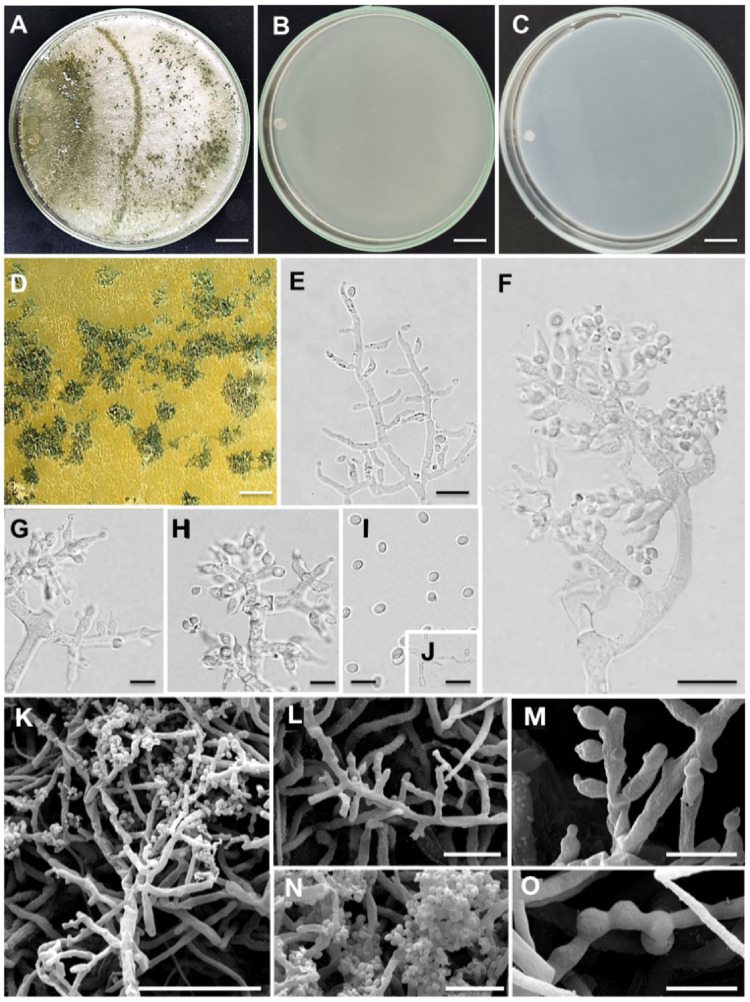 Figure 4
Figure 4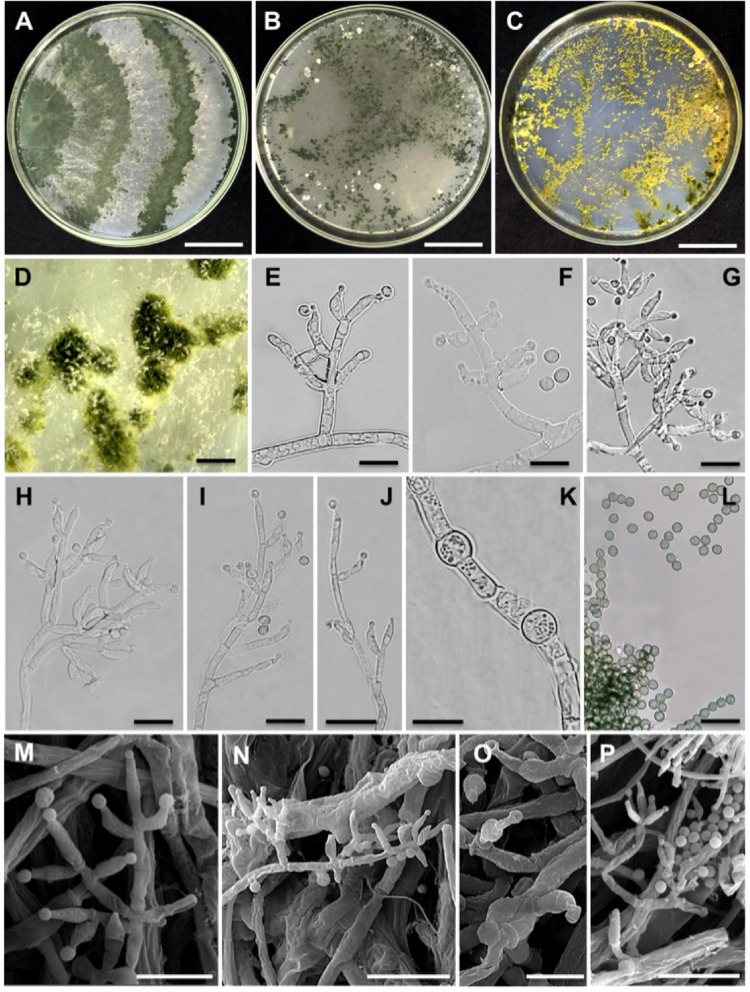 Figure 5
Figure 5- —Universidade Federal De Lavras
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Fungal and yeast genetics research · Plant pathogens and resistance mechanisms
Introduction
Sclerotia are melanized survival structures produced by microorganisms in the Kingdom Fungi and by Myxomycetes in the Kingdom Amoebozoa [1]. Within the Kingdom Fungi sclerotia appear to be produced only by representatives of the Phyla Ascomycota and Basidiomycota and were documented in at least 85 fungal genera in 20 orders [1]. These structures remain viable in soil for long periods of time and are resistant to chemicals, adverse conditions and biological degradation [2]. Sclerotia are very diverse in terms of morphology, composition and physiology. While some have a round shape, smooth surface, dark color and a differentiated ring encircling undifferentiated hyphae, such as the ones produced by Agroathelia rolfsii, others have an irregular shape, undifferentiated ring and lighter color, such as the ones produced by Rhizoctonia solani. Sclerotia produced by Macrophomina phaseolina, Stromatinia cepivora and Verticillium dahliae are smaller than 1 mm, but the ones produced by Polyporus mylittae may reach up to 40 cm in diameter [3].
The species Sclerotinia sclerotiorum, S. cepivora, R. solani, A. rolfsii and M. phaseolina are among the most harmful sclerotia-producing pathogenic fungi in agriculture worldwide [4–7]. These pathogens are hard to manage due to their survival capabilities through sclerotia associated with successive monocultures in the same area and the lack of efficient soil fumigation alternatives.
Biological control with mycoparasites such as Trichoderma is an interesting strategy to decrease the number of sclerotia in soil and thereby control these pathogenic fungi. All Trichoderma species are known for having the ability to parasitize other fungi, a characteristic that was inherited from the ancestor of this genus [8]. The species T. hamatum, T. viride, T. aureoviride, T. harzianum, T. koningii, T. pseudokoningii, T. longibrachiatum and T. asperellum were reported as parasites of sclerotia [9–12]. This mycoparasitic capacity varies among species and strains of the same species. Trichoderma is highly speciose, currently containing approximately 400 described species [13] and most of them are still unexplored for potential biotechnological applications, such as sclerotia mycoparasitism and control of pathogenic fungi.
During our studies on the fungal diversity of the Brazilian Restinga ecosystem we isolated 26 Trichoderma strains, 22 were found to be related to T. spirale and were reported in another study done by our research group [14]. The other four strains represent two novel species of Trichoderma and will be illustrated and described in this study. Furthermore, the novel species were tested for their potential to parasitize sclerotia of fungal pathogens. We studied a Brazilian strain of a recently described species, T. nordicum [15] and made observations on its taxonomy, mycoparasitic activity and distribution.
Materials and Methods
Fungal Strains and Growth Conditions
The fungi used in this study (Table 1) included four strains of the novel Trichoderma species isolated from the Restinga ecosystem: URMicro 11718, URMicro 11720, URMicro 11721 and URMicro 11723, and one strain of T. nordicum, URMicro 11722, isolated from the Amazonian region of Brazil. Trichoderma harzianum H1 was isolated from a commercial product recommended to control S. sclerotiorum in Brazil and was used for comparison purposes in the experiments to study sclerotia parasitism, but was not included in our phylogenetic analyses. The sclerotia-producing pathogen S. cepivora strain JT101 was obtained from an infected garlic plant in the municipality of São Gotardo, MG, Brazil and S. sclerotiorum strain JT205 was isolated from an infected soybean plant at the municipality of Lavras, MG, Brazil. All fungal strains were routinely grown on PDA (Potato Dextrose Agar). Trichoderma and S. sclerotiorum were grown at 25 °C and S. cepivora at 17 °C. Sclerotia were produced on plates incubated at the above temperatures after 14 days. All strains were preserved at −20 °C in the working collection of the Molecular Phytopathology Laboratory and the Trichoderma strains were also deposited in the URMicro culture collection at −80 °C, Federal University of Lavras, Brazil, which is registered with the World Federation for Culture Collections and can make the strains available for research purposes upon request (Table 1).
Table 1. Accession numbers of sequences used in this study to infer phylogenies. The newly described species are shown in bold and the type strains are indicated with a superscript TSpeciesCladeStrainAccession numberstef1rpb2cal1 T. applanatum Hypocreanum7792 ^T^KJ634759KJ634726- T. austriacum HypocreanumCBS 122494 ^T^FJ860619FJ860525- T. confluens Hypocreanum9649 ^T^KT001959KT001964- T. decipiens HypocreanumCBS 121307 ^T^; GJS 91–101FJ860635DQ835520- T. phellinicola HypocreanumCBS 119283 ^T^FJ860672FJ860569- T. pulvinatum HypocreanumCBS 121279 ^T^FJ860683FJ860577- T. protopulvinatum HypocreanumCBS 739.83 ^T^FJ860679DQ835463- T. sulphureum HypocreanumCBS 119929 ^T^FJ860710FJ179620- T. variabile HypocreanumURMicro 11718OR762225OR762229OR762234 T. variabile HypocreanumURMicro 11720OR762224OR762228OR762233 T. variabile HypocreanumURMicro 11721 ^T^OR762226OR762230OR762235 T. victoriense HypocreanumGJS 99–200 ^T^DQ835500DQ835517- T. alboviride LongibrachiatumTC916; HMAS 247224 ^T^MF371230MF371215- T. andinense LongibrachiatumGJS 90–140 ^T^AY956321AY956321JN175412 T. aquatica LongibrachiatumYMF 1.04625 ^T^MK775507MK775512- T. awajun LongibrachiatumCP24_7 ^T^MW480138MW480147- T. awajun LongibrachiatumCP24_8MW480139MW480148- T. bissettii LongibrachiatumP3572PV081282PV081389- T. bissettii LongibrachiatumP3606PV081290PV081397- T. bissettii LongibrachiatumSZMC 25718--MN650686 T. bissettii LongibrachiatumSZMC 1012--MN641032 T. bissettii LongibrachiatumSZMC 1158--MN641033 T. bissettii LongibrachiatumSZMC 1767--MN650683 T. britdaniae LongibrachiatumK 89878 ^T^JQ685865JQ685881- T. caeruleimontis LongibrachiatumPPRI 23903 ^T^--MF355410 T. caeruleimontis LongibrachiatumTri 43--MF355412 T. caeruleimontis LongibrachiatumTri 62--MF355411 T. euskadiense LongibrachiatumCBS 130013 ^T^KJ665492KJ665269- T. flagellatum LongibrachiatumPPRC-ET58; GJS 10–164 ^T^FJ763184KR297247- T. gillesii LongibrachiatumGJS 00–72 ^T^JN175583JN175527JN175409 T. gracile LongibrachiatumGJS 10–263 ^T^JN175598JN175547JN175427 T. konilangbra LongibrachiatumGJS 96–147AY937425KJ665284- T. longibrachiatum LongibrachiatumCBS 816.68 ^T^AY865640DQ087242EU401459 T. longibrachiatum LongibrachiatumDAOM 166989EU338335EU338339- T. longibrachiatum LongibrachiatumATCC 18648 ^T^AY937412HQ260615MBDJ01001225 T. longibrachiatum LongibrachiatumGJS 01–121JN175564JN175507JN175387 T. longibrachiatum LongibrachiatumGJS 08–198JN175565JN175508JN175388 T. longibrachiatum LongibrachiatumGJS 04–53JN175568JN175512- T. longibrachiatum LongibrachiatumGJS 07–21JN175569JN175513- T. longibrachiatum LongibrachiatumS328JQ685867KJ665291- T. longibrachiatum LongibrachiatumDAOM 167674EU280046KJ842212- T. longibrachiatum LongibrachiatumLESF009KT279040KT278975- T. longibrachiatum LongibrachiatumTLC4MT069949MT069960- T. longibrachiatum LongibrachiatumTLS6MT069951MT069959- T. longibrachiatum LongibrachiatumTloum3MT081437MT118251- T. longibrachiatum LongibrachiatumHL167MZ241241MZ241240- T. longibrachiatum LongibrachiatumHNDF-G-2MW504035MZ357387- T. longibrachiatum LongibrachiatumHNDF-G-4MW504037MZ357389- T. longibrachiatum LongibrachiatumHNDF-G-5MW504038MZ357390- T. longibrachiatum LongibrachiatumJZBQT8Z7ON649943ON649996- T. longibrachiatum LongibrachiatumSC5ON808971ON815270- T. longibrachiatum LongibrachiatumD612ON934353ON934388- T. longibrachiatum LongibrachiatumE105ON934356ON934391- T. longibrachiatum LongibrachiatumE106ON934357ON934392- T. longibrachiatum LongibrachiatumE356ON934360ON934395- T. longibrachiatum LongibrachiatumE357ON934361ON934396- T. longibrachiatum LongibrachiatumE473ON934362ON934397- T. longibrachiatum LongibrachiatumE475ON934363ON934398- T. longibrachiatum LongibrachiatumKUFA0409OP132641OP132653- T. longibrachiatum LongibrachiatumKUFA0410OP132642OP132654- T. longibrachiatum LongibrachiatumCHE-CNRCB-334OP410938OP410935- T. longibrachiatum Longibrachiatum593OR548051OR548102- T. longibrachiatum Longibrachiatum539OR548074OR548106- T. longibrachiatum LongibrachiatumFAVF335PP195794PP195792- T. longibrachiatum LongibrachiatumFAVF340PP195795PP195793- T. longibrachiatum LongibrachiatumCEN1281ON101480PP854164ON241222 T. longibrachiatum LongibrachiatumCEN1562ON101481PP854165- T. longibrachiatum LongibrachiatumCHE-CNRCB 1292PV555431PV555433- T. longibrachiatum LongibrachiatumCHE-CNRCB 1293PV555432PV579007- T. longibrachiatum LongibrachiatumCHE-CNRCB 1294PV579006PV579008- T. longibrachiatum LongibrachiatumIAA 1--EU401444 T. longibrachiatum LongibrachiatumDAOM 231258--EU401480 T. longibrachiatum LongibrachiatumDAOM 231259--EU401479 T. longibrachiatum LongibrachiatumPPRC SS8--EU401486 T. longibrachiatum LongibrachiatumCPK 744--JN182288 T. longibrachiatum LongibrachiatumTp-05--KC967327 T. longibrachiatum LongibrachiatumTp-09--KC967334 T. longibrachiatum LongibrachiatumIMI 287096--EU401462 T. longibrachiatum LongibrachiatumIMI 291014--EU401463 T. longibrachiatum LongibrachiatumATCC 201044--EU401468 T. longibrachiatum LongibrachiatumATCC 208859--EU401469 T. longibrachiatum LongibrachiatumCBS 446.95--EU401470 T. longibrachiatum LongibrachiatumCNM-CM 382--EU401472 T. longibrachiatum LongibrachiatumCNM-CM 2277--EU401475 T. longibrachiatum LongibrachiatumCNM-CM 1798--EU401473 T. longibrachiatum LongibrachiatumTl-03--KC967339 T. longibrachiatum LongibrachiatumTl-04--KC967340 T. longibrachiatum LongibrachiatumTl-08--KC967344 T. longibrachiatum LongibrachiatumTl-10--KC967346 T. longibrachiatum LongibrachiatumTl-12--KC967348 T. longibrachiatum LongibrachiatumTl-14--KC967350 T. longibrachiatum LongibrachiatumSZMC 23386--MN650682 T. longibrachiatum LongibrachiatumUAMH 7955--EU401464 T. longibrachiatum LongibrachiatumUAMH 9515--EU401466 T. longibrachiatum LongibrachiatumCECT 20105--EU401476 T. longibrachiatum LongibrachiatumCECT 2412--EU401491 T. longibrachiatum LongibrachiatumTUB F-363--EU401449 T. longibrachiatum LongibrachiatumTUB F-1036--EU401454 T. longibrachiatum LongibrachiatumTUB F-828--EU401450 T. longibrachiatum LongibrachiatumKazan 8--EU401478 T. longibrachiatum LongibrachiatumTH2--KU554610 T. longibrachiatum LongibrachiatumY 19--EU401457 T. longibrachiatum LongibrachiatumGEV 3550--JASTZJ010000008 T. longibrachiatum LongibrachiatumR6902--JAXQJK010000013 T. longibrachiatum LongibrachiatumM236--ON157427 T. novae-zelandiae LongibrachiatumGJS 81–265 ^T^AY937448.2JN133563JN175406 T. odoratum LongibrachiatumHMAS 271354 ^T^KT224463KT224468- T. orientale LongibrachiatumGJS 88 − 81 ^T^EU401581JN175522EU401448 T. orientale LongibrachiatumTUB F-831--EU401451 T. orientale LongibrachiatumTUB F-837--EU401452 T. orientale LongibrachiatumCPK 3598--JQ425719 T. orientale LongibrachiatumGJS 04–333--JN175399 T. orientale LongibrachiatumGJS 04–316--JN175400 T. orientale LongibrachiatumGJS 10–230--JN175403 T. orientale LongibrachiatumGJS 04–321--N175397 T. orientale LongibrachiatumGJS 10–253--JN388899 T. orientale LongibrachiatumGJS 91–157--EU401461 T. orientale LongibrachiatumBMU09741--MN447669 T. orientale LongibrachiatumDIS 270 F--JN175401 T. orientale LongibrachiatumSZMC IM1--EU401493 T. orientale LongibrachiatumSZMC IM2--EU401494 T. orientale LongibrachiatumUAMH 9573--EU401467 T. orientale LongibrachiatumCECT 2606--EU401477 T. parareesei LongibrachiatumTUB F-1066 ^T^GQ354353HM182963GQ354288 T. patella LongibrachiatumGJS 91–141^T^KJ665630KJ665323- T. pinnatum LongibrachiatumGJS 04–100 ^T^JN175571JN175515JN175395 T. pinnatum LongibrachiatumGJS 02–120JN175516JN175572- T. pinnatum LongibrachiatumLS029-3OK652543OK652544ON890478 T. pluripenicillatum LongibrachiatumYMF 1.06198 ^T^MT070159MT070160- T. pseudobritdaniae LongibrachiatumHMAS 271355 ^T^KT224462KT224466- T. pseudokoningii LongibrachiatumDAOM 167678 ^T^KJ713204KJ842214JN175416 T. reesei LongibrachiatumDAOM 167654 ^T^; CBS 383.78KJ713193HM182969JN180917 T. saturnisporiopsis LongibrachiatumS19 ^T^JQ685869JQ685885JQ349444 T. littericola LongibrachiatumURMicro 11723 ^T^OR762227OR762231OR762236 T. solani LongibrachiatumGJS 08–81 ^T^JN175597JN175546JN175426 T. tremelloides LongibrachiatumCBS 121140 ^T^FJ860714FJ860603-Trichoderma spLongibrachiatumCFCC 80875 APV755843PV617129-Trichoderma spLongibrachiatumCFCC 82906 APV755851PV617137-Trichoderma spLongibrachiatumCFCC 88967 APV755878PV617166-Trichoderma spLongibrachiatumCFCC 89492 APV755881PV617169-Trichoderma sp.LongibrachiatumPF4103 ctg 22--JAMAKC010000022Trichoderma sp.LongibrachiatumCBS 243.63--EU401460 T. nordicum VirideURMicro 11722HG325833OR762232- T. nordicum VirideWT 13001 ^T^MH287501MH287502- T. strigosum VirideDAOM 166121 ^T^EU280019AF545556JN133544
Morphological Characterization
Morphological studies were done on corn meal agar dextrose (CMD), synthetic nutrient agar (SNA) and potato dextrose agar (PDA) according to [15–18]. Growth rates were determined at 15, 20, 25, 30 and 35 °C with a 12/12 h light/dark photoperiod using near UV halogen lamps (300–400 nm) [18]. All experiments were installed with four replicates and were done two times.
The micromorphological characteristics were analyzed on structures produced on SNA after three days of incubation at 25 °C. Images were captured in a microscope with fluorescence using the Apotome system and confocal laser (Carl Zeiss) and a scanning electron microscope STEM-FEG model Clara Tescan. Measurements were performed in at least 50 structures of each type, including conidia, phialides, conidiophores and chlamydospores.
Molecular Characterization
Extraction of DNA was done according to the CTAB protocol [19] with mycelium produced with cultures grown for 3–5 days in potato dextrose broth (PDB) (HiMedia) at 25 °C. The primers and amplification conditions for translation elongation factor 1-alpha (tef1), RNA polymerase subunit II (rpb2) and calmodulin (cal1) are described in [20–22]. PCR products were sequenced by the Sanger method in an ABI 3100 according to the manufacturer’s instructions (Applied Biosystems). DNA sequences were assembled with Sequencher v. 5.1 (Gene Codes).
The sequences were compared with the ones deposited in public databases with the program BlastN in the NCBI platform. Multiple alignments were done with the program MAFFT [23] and edited in the program Mega 7.0 [24]. The phylogenetic analyses were done in the CIPRES portal [25] with the Maximum Likelihood (ML) method. Evolutionary models were selected with the program Jmodeltest 2.1.10 [26] using the corrected Akaike information criterion (AICc). The ML analyses were done with RAxML v.1.3 [27] with rapid bootstrapping parameters and 1,000 resamplings. Bayesian inferences were done using the program MrBayes version 3.2 [28] with four Monte Carlo Markov Chains (MCMC), one hot, three cold in one million generations with samplings at every one thousand generations. At the burning phase, 25% of the trees were excluded and the posterior probabilities (PP) were estimated by the remaining 75% of the trees. Trees were visualized with the program FigTree v1.4.4 [29].
Antagonistic Activity of Trichoderma
The antagonism of Trichoderma was evaluated in bioassays to detect the secretion of diffusible and volatile organic compounds and also their capacity to parasitize sclerotia of S. sclerotiorum and S. cepivora. To test diffusible and volatile compounds, mycelial plugs of 5 mm diameter of each pathogen and of the different Trichoderma strains were placed in opposite positions at 0.5 cm from the edge of 9-cm diameter Petri plates containing PDA medium. Volatile compounds were evaluated in plastic plates split into two compartments (Axygen). Controls were plates containing each of the pathogens alone. Plates were incubated at 17 °C for approximately 7 days, when the controls reached the edge of the plate. This temperature was used to favor sclerotial germination and growth of the pathogens. The diameter of the pathogen colonies was determined with a caliper and percentage of inhibition in relation to the control was calculated for each of the Trichoderma strains [14]. The treatments were strains of the newly described species, T. variabile sp. nov. and T. littericola sp. nov., T. harzianum and T. nordicum. The experiments were installed with five replicates and were done twice.
Evaluation of sclerotia parasitism was performed by depositing a drop (~ 8 uL) of a suspension containing 2 × 10^7^ conidia/mL on the surface of each sclerotia of S. sclerotiorum and S. cepivora, which were produced on petri dishes with PDA medium. The sclerotia of S. cepivora were placed on the surface of sterile filter paper (Whatman no. 2) moistened with sterile water and incubated at 17 °C for 14 days. Sclerotia of S. sclerotiorum were placed on the surface of gerbox boxes containing 150 g of sterilized soil maintained approximately at the field capacity and incubated at 17 °C for 15 days. Each treatment, which were the Trichoderma strains mentioned above, was installed in five replicates of 10 sclerotia of each pathogen and the whole experiment was done twice. After the incubation period, sclerotia were surface sterilized with 70% ethanol for 1 min and 1% sodium hypochlorite for 2 min and three rinses with sterile water and plating on PDA medium and incubation at 17 °C. Sclerotia that showed mycelial growth were considered viable and the percentage of dead sclerotia was determined by observing the colonies of the pathogens and/or Trichoderma emerging from the sclerotia.
The data on antagonism of the different Trichoderma species against the sclerotia-producing fungi were submitted to statistical analysis, including the Shapiro-Wilk normality test, analysis of variance and multiple mean comparison with the Scott-Knott test at 5% probability in the program R using the package Scott-Knott version 1.3–2.3 [30].
Results
Phylogeny and Delimitation of the New Species
Sequences of the genes tef1, rpb2 and cal1 (Table 1) were used to reconstruct phylogenetic trees with the methods maximum likelihood (ML) and Bayesian inference (BI). The individual and concatenated sequences of tef1 and rpb2, and sequences of cal1 supported Trichoderma variabile as a novel species with the maximum level of support in all cases (Fig. 1, Figs. S1, S2 and S3). On the other hand, T. littericola was well supported by ML in all the datasets, but not by the BI analysis in the combined tef1 and rpb2 and in tef1 trees (Fig. 1, Fig. S1). Nevertheless, it appeared as a well-defined branch in all datasets.
Fig. 1. Phylogenetic tree inferred with the maximum likelihood (ML) and Bayesian inference (BI) methods with combined sequences of tef1 and rpb2 genes. The ML tree was inferred with two partitions, 617 bp of tef1 and 544 bp of rpb2 and the evolutionary model TPM3uf + I+G. The species Trichoderma nordicum strains URMicro 11722 and WT 13001 were used as outgroups. The numbers on the nodes represent the bootstraps (1000 resamplings) in ML analysis followed by the posterior probabilities (PP) in the Bayesian analysis. A dash means that the bootstrap or PP values are not significant according to the following reference values for significance: >70% for bootstrap and > 0.95 for PP. The species described as novel in this study are shown in red. The clades of Trichoderma are shown in different background colors. The type strains of the described species are indicated with a superscript T. The scale indicates the number of substitutions per site
BlastN analyses of tef1 sequences of the type strain of Trichoderma variabile sp. nov. URMicro 11721 revealed that it was closest to the type strain of T. confluens, with a 93.9% identity (accession number KT001959) and 76% coverage; its rpb2 sequence was 89.1% identical to the rpb2 of T. taxi (DQ859032) with 99% coverage and cal1 was 80.3% identical to T. protrudens (JN133540) with 80.3% coverage.
The type strain of T. littericola sp. nov. URMicro 11723 had a tef1 sequence 91.9% identical to the tef1 of T. reesei (CP040224) with 98% coverage; its rpb2 was 98.2% identical to the rpb2 of T. longibrachiatum (HQ260615) with 94% coverage; and its cal1 was 96% identical to T. pinnatum (JN175395) with 98% coverage.
Strain URMicro 11722 of T. nordicum, which is not a new species, had its tef1 99.8% (93% coverage) identical to the type strain WT 13001 of T. nordicum (accession number MH287501) and 99.4% identical (98% coverage) to the type strain (T32781) of T. nigricans (accession OP357974). Similarly, its rpb2 sequence was 100% (91% coverage) identical to the type strain of T. nordicum (MH287502) and 100% (75% coverage) identical to the type strain of T. nigricans (accession OP357959).
Antagonism of Trichoderma against Sclerotia-producing Pathogens
The antagonistic potential of six strains of Trichoderma was tested, including strains of the species being described in this study, T. variabile sp. nov. and T. littericola sp. nov., one strain of T. harzianum isolated from a commercial product and one of T. nordicum (Fig. 2). The antagonism of these strains was studied against S. sclerotiorum and S. cepivora considering three mechanisms of activity, production of diffusible compounds, release of volatile organic compounds and sclerotia parasitism. Mycelia and sclerotia of S. cepivora were more sensitive to Trichoderma than the sclerotia and mycelia of S. sclerotiorum (Fig. 2). The mechanism that showed the strongest activity against the pathogens was sclerotia parasitism. The new species T. variabile sp. nov. showed the poorest antagonistic activity in all the three mechanisms against both pathogens, whereas, in general, T. nordicum showed the strongest activity, surpassing T. harzianum in most bioassays. For example, T. nordicum parasitized over 90% of the sclerotia of S. sclerotiorum, while parasitism by T. harzianum was approximately 55% (Fig. 2E). The newly described species T. littericola sp. nov. showed a spectrum of activity similar to T. harzianum, but a slightly higher parasitism (P < 0.05) of S. sclerotiorum sclerotia (Fig. 2E).
Fig. 2. Antagonism of different Trichoderma strains to Sclerotinia sclerotiorum and Stromatinia cepivora in in vitro experiments done twice. A-D Mycelial growth of S. sclerotiorum (A,** C)** and S. cepivora (B,** D)** influenced by diffusible compounds produced by strains of Trichoderma in the culture medium (A,** B)** or by organic volatile compounds (VOCs) released in the headspace of sealed plates split into two compartments (C,** D). E, F** Parasitism of sclerotia of S. sclerotiorum (E) and S. cepivora (F). Data are means of five replicates per treatment per experiment and are shown in percentage of mycelial inhibition or in percentage of sclerotia parasitism. The experiments were done in plates and mycelial growth was measured when the control (pathogens alone) reached the edge of the plates. Parasitism was determined by inoculating sclerotia with spore suspensions of Trichoderma and placing them on the surface of sterilized soil or filter paper, respectively for sclerotia of S. sclerotiorum and S. cepivora. The number of parasitized sclerotia was determined after incubation for two weeks at 17 °C. Means followed by the same letter are not significantly different according to the Scott-Knott test at 5% probability. Comparisons should be done among means of treatments in the same experiment, not between experiments. Treatments without variance (0 or 100%) were removed from the statistical analysis
Taxonomy
Trichoderma variabile J. Rembinski, P.A.S. Marbach, J.T. De Souza, sp. nov. Figure 3.
Fig. 3. Morphological characteristics of Trichoderma variabile sp. nov. strain URMicro 11721. A Growth on PDA, B on SNA and C on CMD at 25 °C after 7 days. D,** E** Pustules and conidiophores in a stereomicroscope. F-P Conidiophores (F-L), phialides (K-N), conidia (N, O) and chlamydospores (P) in optical microscopy. Q-W Conidiophores (Q-U), phialides (R-V) and conidia (W) in scanning electron microscopy (SEM) micrographs. Scale bars: (A-C) = 14 mm; (D,** E**) = 5 mm; (F-P) = 5 μm; (Q-W) = 50 μm
MycoBank: MB 854568.
Type: Brazil, Bahia state, Valença municipality, Guaibim Restinga (13° 18’ 03” S/38° 57’ 57” W) from leaf litter samples, 10/11/2012, collected by J.P. Andrade. Holotype HURB 38020, metabolically inactive, dried culture on PDA deposited in the Herbarium of Recôncavo da Bahia Federal University (UFRB). Ex-type culture deposited at the culture collection of UFRB under number MON21 and at Federal University of Lavras under number URMicro 11721 at −80 °C. Accession numbers: tef1 = OR762226; rpb2 = OR762230; cal1 = OR762235.
Etymology: variabile refers to the variable morpho-physiological characteristics of different strains of this species.
Additional strains examined: Brazil, Bahia, Valença, Guaibim Restinga (13° 18’ 03” S/38° 57’ 57” W), from leaf litter, 10/11/2012, J.P. Andrade. Strains: MON10A = URMicro 11718; MON10B = URMicro 11719 and MON19 = URMicro 11720 preserved in glycerol at −80 °C in the collections of Federal University of Recôncavo da Bahia (Bahia state) and Federal University of Lavras (Minas Gerais state).
Description: Conidiophores verticillium- and pachybasium-like, solitary, sometimes ramified and formed on sparse mycelia. Phialides in asymmetrical arrangement, varying from 1 to 3, lageniform, ampulliform and subulate, measuring (7.5–)8–16(–20) µm long, 2–3.5(–4) µm at the widest point, L/W ratio 3.5–8(–10), 1.5–2 μm (n = 50) at the base, surging from a cell 2–3 μm (n = 50). Conidia ellipsoidal and globose, smooth, varying from pale to dark green, measuring (2–) 3.5–7 × 2–4.5 μm (n = 50), L/W ratio (0.77)1–3.5. Chlamydospores 3–7 × 4–7 μm (n = 50), abundant, globose and suboval. Teleomorph not observed.
Culture characteristics: colony growth on SNA at 25 °C after 72 h, radius 35 mm, at 35 °C, 30 mm and at 15 °C, 25 mm. Mycelium covers the plate in 7 d at 25 °C. Colony initially hyaline turning dark green with conidiation, which starts at 96 h. Pustules distant from the inoculation point, mostly at the periphery of the plate (Fig. 3B), solitary to aggregated, sparse and circular, initially white, turning light green and later dark green, 5–10 mm. No distinct odor noticed. No pigment observed. On PDA at 25 °C after 72 h, colony radius 43 mm, at 35 °C, 38 mm, at 15 °C, 29 mm. Mycelium covers the plate in 7 d at 25 °C. Colonies are smooth, dense and white, turning green at conidiation, color is lighter when compared to SNA. Conidiation begins 96 h after inoculation, in concentric rings without forming pustules, directly on aerial mycelia (Fig. 3A). Light coconut odor noticed. No pigment observed. On CMD at 25 °C after 72 h, colony radius 32 mm; at 35 °C, 28 mm, at 15 °C, 26 mm. Mycelium covering the plate in 7 d at 25 °C. Colonies initially white, turning yellow and later on light green. Conidiation begins 96 h after the inoculation, on pustules distant from the inoculation point, produced in concentric rings at the plate periphery (Fig. 3C). No distinct odor noticed. No pigment observed.
Distribution: Only known from Brazil, Valença, Guaibim Restinga.
Notes: The species T. variabile fell in a well-supported cluster, separated from its closest relatives (Fig. 1). Trichoderma variabile could not be distinguished from its closest relatives, T. confluens, T. decipiens and T. applanatum on the basis of conidia and phialide morphology as well as in optimal growth temperatures [31–35; Table S2]. However, the growth rate on PDA at 25 °C differed among these species. Trichoderma variabile filled the plate in 7 days, whereas T. confluens did it in 3–4 days and the other two species, T. applanatum and T. decipiens were similar, taking 10–12 days (Table S2). Trichoderma applanatum was the only one with hyaline conidia and T. decipiens the only one producing a pinkish white pigment in the medium, whereas the other species, including T. variabile, did not release any pigment in the culture medium (Table S2).
Trichoderma littericola J. Rembinski, P.A.S. Marbach, J.T. De Souza, sp. nov. Figure 4.
Fig. 4. Morphological characteristics of Trichoderma littericola sp. nov. strain URMicro 11723. A Growth on PDA, B on SNA and C on CMD at 25 °C after 7 days. D Pustules on PDA under a stereomicroscope. E-J Conidiophores (E-H), phialides (E-H), conidia (I) and chlamydospores (J) in optical microscopy. K-O Conidiophores (K, L), phialides (L, M), conidia (K, M, N) and chlamydospores (O) in scanning electron microscopy (SEM) micrographs. Bars: (A-C) = 14 mm; (D) = 5 mm; (E-J) = 5 μm; (K-O) = 50 μm
MycoBank: MB 854567.
Type: Brazil, Bahia state, Guaibim Restinga (13°18’36” S/38° 58’ 13” W), from leaf litter, 02/10/2012, P.A.S. Marbach. Holotype HURB 38021, metabolically inactive, dried culture on PDA deposited in the UFRB herbarium. Ex-type culture URMicro 11723 deposited in the culture collection of Federal University of Lavras and at Federal University of Recôncavo da Bahia under number MTS13 at −80 °C. Accession numbers: tef1 = OR762227; rpb2 = OR762231; cal1 = OR762236.
Etymology: littericola is a reference to leaf litter, the isolation source of this species. Description: Conidiophores trichoderma-like, formed on aerial mycelium and on small pustules (2–5 mm). Conidiophores typically with a central axis from which phialides are formed in vertexes and/or solitary. Phialides (3–)4.2–9(–10) µm long, (2–)2.2–3.8(–4) µm at the widest point (n = 50), L/W ratio (1.8)2–2.8(6), (1–)1.5–2.2(− 3) µm at the base, originating from a cell (1.4–)2–3(–3.5) µm of width, typically lageniform to ampulliform in the aerial mycelium. Conidia 2–3.5(–5) × 1.7–3 μm, L/W ratio (0.66)1.16–1.35(1.6) (n = 50), ellipsoidal and globose, smooth, light to dark green. Chlamydospores 7–10 × 3–5 μm (n = 50), globose to ovoid, abundant, intercalary and terminal. Teleomorph not observed.
Culture characteristics: optimum growth temperature on PDA, SNA and CMD, 25 °C. On PDA after 72 h, 90 mm at 15, 25 and 35 °C. Colonies were white transitioning to yellow and finally olive green and dark green. Pustules, small (3–7) mm, sparse, observed in the whole plate. On SNA after 72 h, 90 mm at 15, 25 and 35 °C. Colonies hyaline, difficult to observe (Fig. 4B). No pustules formed. On CMD after 72 h, 90 mm at 15, 25 and 35 °C. Colonies hyaline, sparse, without the formation of pustules, culture medium with a pallid yellow olivaceous pigment. Indistinct odor detected.
Distribution: Only known from Brazil, Valença, Guaibim Restinga.
Notes: Trichoderma littericola has shown fast growth at 35 °C and yellow pigment in the culture medium (PDA), and these are common characteristics in representatives of the Longibrachiatum clade [31]. The morphological characteristics, including the dimensions of the phialides, conidia and chlamydospores did not differentiate T. littericola from its closely-related species, T. pinnatum and T. orientale [31, 32]. However, chlamidospores were not observed in cultures of T. pinnatum, separating this species from T. littericola and T. orientale. Pustules were small (0.25–1 mm) in cultures of T. pinnatum, but these structures were not produced by T. littericola on SNA, differentiating these two species. The growth rate of T. littericola was the fastest, followed by T. pinnatum and T. orientale, although the temperature ranges were slightly different. Trichoderma littericola filled PDA and SNA plates in 3 days, whereas T. pinnatum took 3 days to fill PDA plates and 4 days to fill SNA plates and T. orientale filled both PDA and SNA plates in 4 days (Table S2).
Trichoderma nordicum G.Z. Zhang [15], MycoBank: MB 8212301. Accession numbers: tef1 = MH287501; rpb2 = MH287502.
Synonym: Trichoderma nigricans C.L. Zhang [33], MycoBank: MB 845506. Accession numbers: tef1 = OP357974; rpb2 = OP357959.
Illustrations and descriptions: [15, 33]; Fig. 5; Tables S3 and S4.
Fig. 5. Morphological characteristics of Trichoderma nordicum strain URMicro 11722. A Growth on PDA, B on SNA and C on CMD at 25 °C after 7 days. D Pustules and conidiophores under a stereomicroscope. E-L Conidiophores (E-J), phialides (G-I), conidia (F, I, L) and chlamydospores (K) in optical microscopy. M-P Conidiophores (M, N, P), phialides (M-P) and conidia (M, N, P) in scanning electron microscopy (SEM) micrographs. Bars: (A-C) = 23 mm; (D) = 5 mm; (E-L) = 5 μm; (M-P) = 50 μm
Strain examined: Brazil, Amazonas, Manaus (3° 05’ 28” S/59° 59’ 36” W), soil, 12/10/2012. J.T. De Souza. Culture URMicro 11722. Accession numbers: tef1 = HG325833; rpb2 = OR762232.
Description: Conidiophores in a main axis, with lateral branches frequently at right angles. Phialides solitary or 2–3 in a spiral or cruciform pattern. Micromorphology of phialides, conidia, chlamydospores and culture characteristics in comparison with the Chinese strains are shown in Table S4. Cultures without any noticeable pigment or distinct odor.
Distribution: Asia (China, India, Sri Lanka, Taiwan), Americas (Brazil, Mexico, USA) (Table S3).
Notes: T. nordicum [15] and T. nigricans [33] are genetically and morphologically indistinguishable (Fig. S4; Table S4) and therefore should be synonymized under T. nordicum, according to the priority of publication. Strain URMicro 11722 of T. nordicum differs from the described Chinese strains by growing faster at 35 °C, but is similar in the other micromorphological characteristics (Table S4). Other sequences deposited in the database and misidentified, mainly as T. atroviride and as Trichoderma sp. were analyzed and correctly identified as T. nordicum (Table S3; Fig. S4).
Discussion
In this study, two novel species of Trichoderma were described, T. variabile and T. littericola, and a third species, T. nordicum, was illustrated and T. nigricans was synonymized with it on the basis of taxonomic priority. The antagonistic potential of these Trichoderma species was tested in vitro, considering three mechanisms of activity, secretion of diffusible and volatile compounds, and parasitism of sclerotia. The results of this study extended our knowledge on the diversity of Trichoderma in the Restinga environment and point out for the potential application of the studied species in the biocontrol of sclerotia-producing plant pathogens.
Trichoderma is a hyperdiverse genus, with approximately 400 species described worldwide in 2021 [13, 34]. In this study we used sequences of tef1, rpb2 and cal genes to infer phylogenies and to describe two novel Trichoderma species. The best marker to identify Trichoderma at the species level in terms of discriminatory power and representativeness in public databases is tef1 [13, 14]. This marker is followed by cal in discriminatory power, but it unfortunately has a poor representativeness in public databases. On the other hand, rpb2 has a good representativeness and ranks third in terms of discriminatory power [14]. Fragments of the ITS region of the rDNA have been recommended in Trichoderma phylogenies [13]. However, this region has no discriminatory power to distinguish Trichoderma species and is commonly not included in the description of novel species [14, 15, 17, 22, 31, 33, 35–37]. Due to the reasons outlined above, we did not determine the ITS sequences for the novel species that are being described in this study.
Studies showing the diversity of Trichoderma in natural and agricultural areas [37, 38], certain environments, such as the Restinga, are not well represented. The Restinga is an endangered ecosystem due to its exposure to disorganized economical exploitation and human occupation, and rising sea levels attributed to global warming [39]. The Restinga ecosystem harbors a diverse fungal community that may be exploited in biotechnological applications, such as the biocontrol of fungal pathogens. Our research group is systematically sampling the Restinga ecosystem in Brazil to study the fungal communities occurring in this environment, with emphasis on the genus Trichoderma with the objective of ex-situ preservation and application in the control of plant diseases, especially the ones caused by sclerotia-producing soilborne pathogens.
The phylogenetic analysis performed in this study lead to the description of two novel species and also to the invalidation of the species T. nigricans and its synomyzation with T. nordicum. One of the problems frequently encountered in fungal taxonomy is the incompleteness of sequence databases and the lack of standardization of the sequenced regions. In this case study, T. nordicum was described by [15], but the sequences of the type strain were not annotated as such in the database, leading Zhao et al. [33] to describe T. nigricans. To avoid this type of confusion, it is advisable that all authors make sure the species published by them are fully annotated in sequence databases.
Trichoderma nordicum was previously known only from China [15], but our phylogeographic analysis of deposited sequences revealed that this species is distributed in Asia and the Americas (Table S3). Besides that, T. nordicum was found in very diverse environments and substrates such as soil, dead wood, algae, leaves cut by ants, from healthy plant tissues, manatee skin and turtle shell (Table S3), pointing out to the high degree of opportunism of this species.
The newly described species were isolated from soil collected at the Guaibim Restinga ecosystem. This ecosystem is subjected to extreme conditions, such as high temperature variation, UV radiation and strong winds. The strain of T. nordicum we examined was obtained from the Amazoniam region, which is also exposed to high temperatures. The adaptation to high temperatures of all the studied strains might, at least in part, explain the fast growth rate they had on culture media, with exception of T. variabile, which had a slower growth rate. It appears that, at least in some cases, there is an association between antagonistic activity and growth rate. There is evidence that temperature and the previous adaptation of the Trichoderma strains influence their activity in mycoparasitism and induction of resistance [40–42]. In fact, the strains of T. variabile examined in this study showed a slower growth rate at high temperatures and no inhibition of S. sclerotiorum with diffusible and volatile compounds (Fig. 2). It would be interesting to study the correlation between adaptation to high or low temperatures, for example, and antagonistic activity in the same background species. In this regard, testing Brazilian and Chinese strains of T. nordicum, adapted to annual average temperatures of 25–27 °C in Bahia and Amazonas states (Table S3), Brazil and 12–13 °C in Beijing and Shandong province, China (https://en.climate-data.org/) in sclerotia parasitism in vitro and in field trials would bring important knowledge. Most Trichoderma species are well adapted to grow at temperatures around 25 °C [43, 44]. There are, however, plant pathogens, such as S. sclerotiorum and S. cepivora that cause damage in the field at temperatures lower than 18 °C [45, 46]. In these cases, it is important to select strains adapted to grow, sporulate and survive at these temperatures.
Sclerotinia sclerotiorum produced mycelia and sclerotia that were more resistant to the Trichoderma strains than Stromatinia cepivora, although both pathogens are in the same family, Sclerotiniaceae. The size, structure and resistance of the sclerotia produced by these fungi differ considerably. While sclerotia produced by S. cepivora are 0.2–0.5 mm in diameter, have a thin rind of 1–2 cell layers and can stay viable for up to 20 years in soil, the ones produced by S. sclerotiorum are 2–10 mm in diameter, 2–3 cell layers thick and survive in soil on average for 3 years [47]. The reasons for this differential susceptibility shown by the two pathogens to Trichoderma are still unknown, although there are some indications on the sclerotial phase of these fungi. Sclerotia of Sclerotinia spp. were shown to secrete metabolites [48], and some metabolites have antimicrobial activity, such as phenolic and ferulic acids [49]. However, further studies are necessary to corroborate this hypothesis.
The species T. littericola and T. nordicum showed the highest potential to inhibit and parasitize sclerotia of S. sclerotiorum and S. cepivora, surpassing in most cases the T. harzianum strain used in our comparisons, which is the active ingredient of a commercial product. It is known that Trichoderma species employ several mechanisms of activity, frequently synergistically, depending on the system [50]. In our study, although we did initial tests on three mechanisms of activity commonly employed by Trichoderma, our main targets are the sclerotia and therefore, we consider mycoparasitism as the most important mechanism to decrease the concentration of these structures in soil. We are aware that the relationship between in vitro and in vivo experiments is poor [51], and therefore it is imperative to perform experiments in planta, preferentially in field trials.
All the Trichoderma strains studied were able to produce chlamydospores abundantly, especially on SNA at 25 °C. This opens up the possibility of producing inocula of these strains to apply as biocontrol agents based on these propagules as they are more resistant to adverse conditions than conidia [52]. Chlamydospores are hard to obtain in high concentrations and currently there is a lack of ideal fermentation technologies to produce them efficiently [52]. Understanding the genetic mechanisms that lead to their formation will help improve mass production of these resistance structures and optimize biocontrol formulations.
Conclusions
Two new species of Trichoderma, T. variabile and T. littericola, were described in this study, contributing to increase our knowledge of the fungal diversity in the Restinga ecosystem. Additionally, the potential antagonism in vitro of these newly described species was demonstrated against two sclerotia-producing pathogens of worldwide importance. The species T. nordicum was reported from Brazil and T. nigricans was shown to be an invalid species name. The taxonomical novelties reported herein advance our knowledge on the diversity of the genus Trichoderma.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1R Core Team. A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (2021) Available at: https://www.R-project.org/