Independent evolution of a living bridge in the old world army ant lineage
Nobuaki Mizumoto, Kôichi Arimoto, Clement Het Kaliang, Taisuke Kanao

TL;DR
Army ants in the Old World were observed forming living bridges, similar to those seen in New World species, suggesting this behavior evolved independently.
Contribution
The first observation of living bridge formation in Old World army ants, indicating convergent evolution of this behavior.
Findings
Aenictus glabrinotum ants formed living bridges to span gaps in their foraging trails.
Bridges were maintained during high traffic and disassembled when traffic declined, mirroring New World species behavior.
This suggests bridge formation may have evolved independently in different army ant lineages.
Abstract
Living bridges formed by army ants are among the most striking examples of collective behavior in social insects, previously known only from New World Eciton species. Here, we report the field observation of living bridge formation in an Old World army ant, Aenictus glabrinotum. We artificially created a small gap in the foraging trail, formed on a twig, to disrupt traffic flow. Ants accumulated near the opening, explored the air space, and interlinked their bodies to form a living bridge, allowing traffic to resume. Bridges were maintained while traffic was high but spontaneously disassembled as flow declined, suggesting a similar behavioral process to that of Eciton behavior. Although it is limited to a single opportunistic observation, this documentation suggests that bridge-formation may have evolved several times in army ants. By motivating further comparative studies across army…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
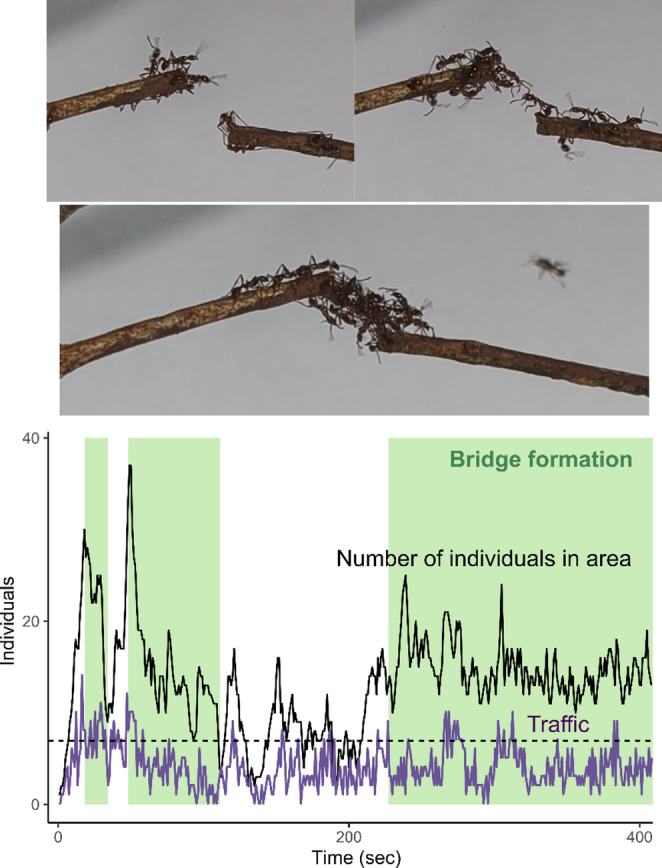 Figure 1
Figure 1 Figure 2
Figure 2- —https://doi.org/10.13039/501100001691Japan Society for the Promotion of Science
- —https://doi.org/10.13039/100005825National Institute of Food and Agriculture
- —JST/JICA, SATREPS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Neurobiology and Insect Physiology Research · Evolutionary Game Theory and Cooperation
Introduction
Collective behavior enables groups of animals to perform tasks that individuals cannot accomplish alone. One striking example is self-assembly, in which individuals physically link their bodies to create dynamic and flexible three-dimensional structures (Anderson et al. 2002; Carlesso and Reid 2023). Such structures provide functional advantages, otherwise impossible for single individuals, such as forming temporary shelters, bridging gaps to facilitate movements, or combining forces to achieve greater strength (Anderson et al. 2002; Carlesso and Reid 2023). Self-assembly is rare across animals and is largely restricted to several derived social Hymenopteran species, e.g., fire ant rafts, honey bee swarms, weaver ant chains, and army ant bivouacs (Carlesso and Reid 2023). Although these are system-specific phenomena, similar ecological pressures may promote the convergent evolution of self-assembly behaviors across unrelated taxa.
Army ants represent a particularly charismatic example of collective behavior, exhibiting a suite of traits known as the “army ant syndrome”, including obligate group foraging and nomadic colony cycles (Schneirla 1971; Gotwald 1995; Kronauer 2020). This syndrome has evolved convergently in two lineages: the New World and the Old World army ants (Borowiec 2019). Among these, the genera Eciton (New World) and Aenictus (Old World) are especially surface-adapted, typically foraging and migrating on open ground, rather than within leaf litter or subterranean tunnels (Gotwald 1995). Their movements depend on the continuity of the ground surface, potentially disrupted by small gaps (Mizumoto and Reid 2024). The New World army ants, Eciton, are well known for their ability to overcome such spatial discontinuities by forming living bridges that span gaps, and the behavioral mechanisms underlying this phenomenon have been well documented (Reid et al. 2015; McCreery et al. 2022). In contrast, behavioral and life history information remain scarce in Aenictus species. Unlike Eciton ants, Aenictus lacks polymorphic workers and forms a simpler collective foraging system (Schneirla 1971; Gotwald 1995; Yamane et al. 2021), yet their surface-adapted lifestyle suggests they might also employ similar bridging behaviors.
Here, we report observations of living bridges in *A. glabrinotum *(Fig. 1, Video S1). This finding provides rare insight into collective behavior in the Old World army ants and suggests that comparable ecological challenges may have driven similar self-assembly behaviors across independent lineages of army ants.
Fig. 1. Living bridge formation by Aenictus glabrinotum. Time development of the number of individuals around the gap (25 mm) was recorded (see Figure S1 for the scale). Also, traffic was measured as the number of individuals entering or leaving this area per second. The dashed horizontal line indicates the minimum number of ants in the gap area while the bridge was formed
Methods
All observations were performed in the Lambir Hills National Park, Miri, Sarawak, Malaysia. Between May 31 and June 12, 2023, we conducted daily field surveys of marching termites (Hospitalitermes spp. and Longipeditermes longipes) along the ~ 4 km trail (Inoue-Pantu-Main trail), walking the route once or twice per day to search for termite trails. During these surveys, we occasionally encountered foraging or emigration trails of Aenictus spp., both during the day and at night. On June 6, while returning to the chalet after observations on Hospitalitermes lividiceps, we encountered the tail of an A. glabrinotum colony at 10:20 pm local time (4.2023437° N, 114.0302110° E). There are 18 Aenictus species in the Lambir Hills National Park (Yamane et al. 2021), and the information about the species-level identity is provided in the Supplementary Text S1. The trail crossed a narrow twig of dead wood on the forest floor, providing a natural opportunity to test whether this species could form a living bridge when the substrate was interrupted.
We cut the twig bridge with scissors and recorded the ant responses using the 4 K video camera (HC-X1500-K, Panasonic) at 30 fps, illuminated by an LED video light (Pixel G1s) (Fig. 1). The resulting gap was approximately 5.3 mm wide, slightly larger than the body length of ants (4.11 ± 0.34 mm, Mean ± S.D.; n = 10, measured from the snapshot of the video). Preliminary observations indicated that LED lights at night did not disturb trail activity. Recording lasted for about 400 s, limited by battery capacity and safety considerations for returning from the field site.
To quantify traffic dynamics during bridge formation, we analyzed the video using the open-source event-logging software BORIS (Friard and Gamba 2016). The observation area was defined as a 50 × 50 mm square centered on the twig gap (Fig. S1). We counted the number of ants entering and leaving the area each second and binned the data accordingly. The traffic flow was defined as the total number of individuals entering or leaving the area per second, and the estimated number of ants present was derived from cumulative entry and exit counts over the observation period. These data were used to describe the temporal dynamics of collective bridge formation.
The autocorrelation function of the traffic flow data shows no detectable correlation at lags > 11 s (Fig. S2). Thus, we obtained the total traffic flow and the mean number of ants near the gap for each 11-second bin. We investigated the effects of the mean number of ants near the gap, bridge formation, and interactions on traffic flow using a generalized linear model (GLM) with a Poisson error distribution and a log link function. The likelihood ratio test was used to determine the statistical significance of each explanatory variable (type II test). All analyses were performed using R (R Core Team 2024).
Results
During the observation period, we observed three bridge formation events and two spontaneous collapses (Fig. 1). After a gap was created, traffic was disrupted, and some ants went back in the other direction, while others stayed near the gap to explore the air by moving their antennae and fore legs (Video S1). Traffic disruptions sometimes led to congestion, with ants accumulating near the opening (Fig. 1). The decision to explore the airspace or turn back seemed to be made individually, rather than one forcing the other. When ants from both sides explored the air and made contact, they antennated each other, then interlocked their bodies to form a living bridge. Once the bridge was established, traffic resumed, and the local ant density around the gap decreased. The bridge persisted as long as ants continued to cross but collapsed spontaneously when traffic declined (Fig. 1).
During bridge formation, the minimum number of ants present in the gap area was seven, indicating that at least seven individuals were required to span the 5.3 mm gap. The number of ants near the gap was positively correlated with traffic flow (GLM; slope, 95%CI 0.039—0.066, χ^2^₁ = 125.4, P < 0.001, Fig. S3, Table S1), and the slope of this relationship was significantly steeper when the bridge was present (GLM; interaction, 95%CI: 0.002—0.047 χ^2^1 = 4.7, P = 0.03, Fig. S3, Table S1).
Discussion
Old World and New World army ants show many convergent traits, yet also display key differences in their collective behaviors. Both Aenictus and Eciton are surface-adapted foragers (Gotwald 1995), but Eciton species typically form larger and more complex raid networks, while Aenictus raids tend to be simpler (Schneirla 1971; Gotwald 1995). Our observations reveal that Aenictus ants can also form living bridges. However, bridge formation in Aenictus appears more limited, typically spanning only small gaps and often breaking down spontaneously (Fig. 1). This likely reflects the lower overall traffic intensity in Aenictus trails (Schneirla and Reyes 1966). Also, as the gap was artificially introduced in this study, it remains unknown whether Aenictus forms a living bridge to actively cross the gap or only to repair the existing trail. Repetitive observations on larger-scale trials could further clarify the bridge formation by Aenictus ants, confirming our anecdotal observation.
Living bridge formation in Eciton workers can emerge from simple behavioral rules. Individual ants track the rate of traffic passing over them and leave the bridge when tactile stimulation drops below a certain threshold (Powell and Franks 2007; Garnier et al. 2013; McCreery et al. 2022). Although our data cannot quantitatively test these behavioral rules, the observed traffic dynamics suggest that A. glabrinotum may employ a similar process. We recorded two spontaneous bridge collapses, and the second event coincided with declines in local traffic and ant density (Fig. 1). This should also be tested with multiple observations. Among self-assembly behaviors in social Hymenoptera, bivouac formation is the most widespread, providing a temporary shelter for colony members (Anderson et al. 2002; Carlesso and Reid 2023). Both Old World and New World army ants form bivouacs, including several Aenictus species (Gotwald 1995). We also observed a self-assembling bivouac of A. glabrinotum at our field site, including the hanging chain (Text S1, Figs. 2 and S4, Video S2). The underlying behavioral process governing bivouac and bridge formation might be shared in Aenictus glabrinotum.
In conclusion, we document the first evidence of living bridge formation in Old World army ants. Although this behavior is well known in Eciton, its evolutionary origin remains unclear. Unique foraging behavior in army ants, such as mass raiding, is suggested to have continuously evolved from group foraging (Chandra et al. 2021). Yet, behavioral information within the clade of Old World army ants of the subfamily Dorylinae remains scarce. Although based on a single observation, documenting behaviors of understudied but evolutionarily important species is a valuable natural history, particularly by generating hypotheses for future comparative studies. Our findings underscore the continuing importance of natural history observations in revealing behavioral diversity and convergence among social insects.
Fig. 2A self-assembled chain formation similar to a living bridge by Aenictus glabrinotum. The photo was taken on June 6, 2023, on the same day as the bridge observations in the same field locality (see Text S1, Figure S4, and Video S2)
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
Supplementary Material 3