Naked cuticle is essential for Drosophila wing development beyond Wingless signaling
Rui Wang, Ping Wang

TL;DR
The gene naked cuticle is crucial for Drosophila wing development, influencing multiple signaling pathways beyond its known role in Wnt signaling.
Contribution
This study reveals that Nkd integrates multiple signaling pathways during wing development, beyond its role in Wingless signaling.
Findings
Genetic disruption of nkd affects larval development and adult wing morphology.
High-throughput sequencing shows nkd influences multiple signaling pathways.
Nkd functions as an integrator of signaling pathways during wing development.
Abstract
The evolutionarily conserved gene naked cuticle (nkd) has been gone through studies in both invertebrate and mammalian model systems. Nkd proteins play an essential role in the development of Drosophila as a negative‐feedback regulator for Wingless signaling. In this research study, we showcase the multifaceted functions of Nkd in Drosophila wing development. The disturbance of nkd dosage genetically disrupts multiple biological processes in the larval stage and the morphologies of adult wings. Our results of high‐throughput sequencing indicate that nkd may have profound effects on Drosophila wing development involving more essential signaling pathways. Nkd may serve as a potential integrator of multiple signaling pathways during wing development, extending its functional repertoire beyond simple Wg inhibition. Naked cuticle (Nkd), a Wnt signaling inhibitor, assumes extensive roles in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
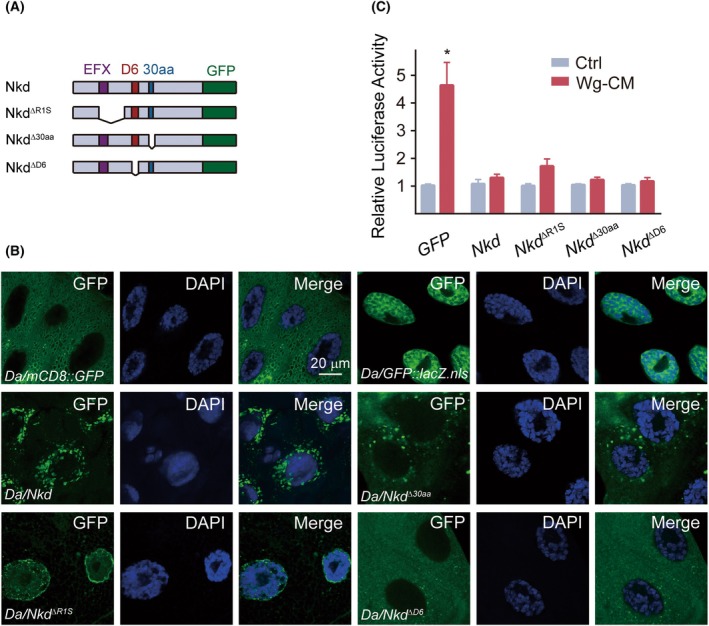 Fig. 1
Fig. 1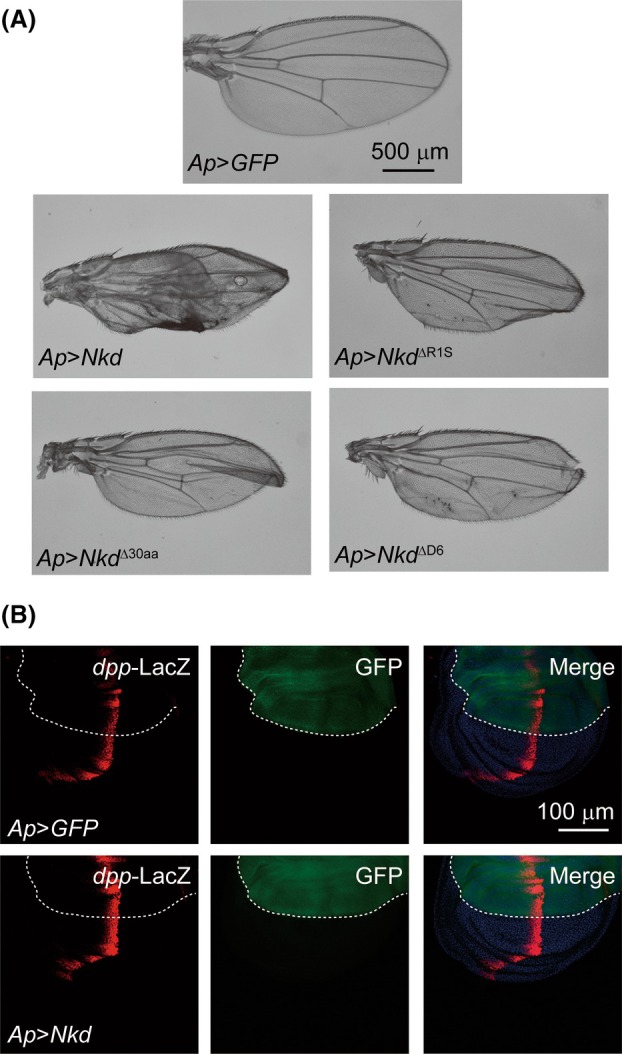 Fig. 2
Fig. 2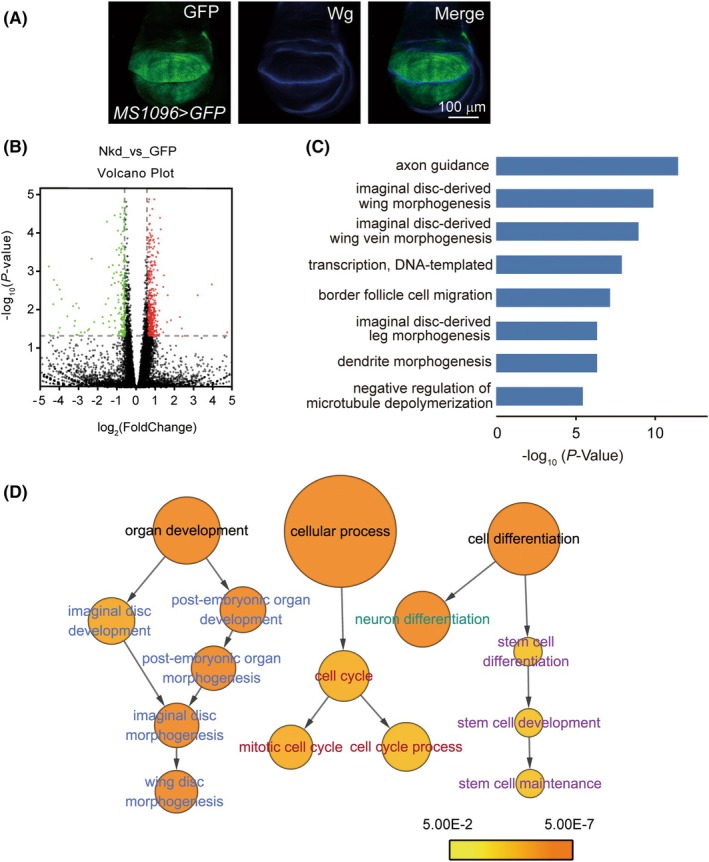 Fig. 3
Fig. 3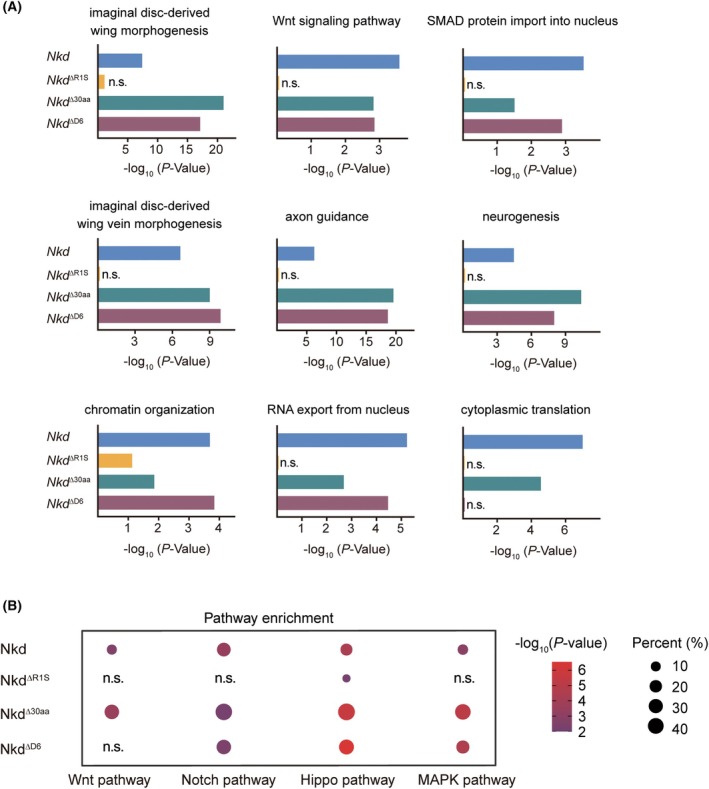 Fig. 4
Fig. 4| Term |
| Genes |
|---|---|---|
| Wnt signaling pathway | 0.002976 |
|
| Notch signaling pathway | 0.006327 |
|
| Hippo signaling pathway | 0.001121 |
|
| MAPK signaling pathway | 0.002976 |
|
- —Fundamental Research Funds for the Central Universities10.13039/501100012226
- —National Natural Science Foundation of China10.13039/501100001809
- —Natural Science Foundation of Jiangsu Province10.13039/501100004608
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWnt/β-catenin signaling in development and cancer · Hippo pathway signaling and YAP/TAZ · Cancer-related gene regulation
The development of complex structures like the Drosophila wing requires the precise integration of multiple conserved signaling pathways. The interplay between these pathways ensures coordinated cell fate specification, proliferation, and tissue patterning. Therefore, identifying molecules that serve as integrators or nodes for cross talk between these pathways is crucial for understanding the fundamental principles of morphogenesis. Transducers, some of which serve as nodes for cross talk between pathways, are commonly essential for morphogenesis during development and elaborate tissue‐specific gene regulation. Given the crucial role of the Wnt pathway in stem cell development and maintenance [1, 2, 3], it is unsurprising that mutations in Wnt signaling transducers are frequently observed in diseases, including carcinomas. The most well‐characterized example is the APC mutation in colon cancer [4, 5, 6]. AXIN1, AXIN2, and β‐catenin mutations have also been reported in a variety of carcinomas as well [7, 8, 9, 10]. Several groups of molecules physiologically attenuate Wnt signaling through distinct mechanisms. Notably, negative‐feedback regulators, such as Notum, Nemo, and Naked cuticle (Nkd), ensure fine balancing of Wnt signaling [11, 12, 13].
Endogenous nkd expression depends on wingless (wg) [12] and the recruitment of Wg‐dependent TCF to the nkd control region has been verified [14], providing evidence that nkd is a direct target gene of Wnt signaling. Later studies demonstrated that the inhibitory function of Nkd depends on specific motifs conferring membrane localization, Dishevelled (Dsh) binding, and two potential nuclear localization signals (NLSs) [15, 16, 17, 18, 19]. Aggregation of a conserved C‐terminal histidine cluster has been proven crucial for promoting Axin destabilization [20]. Nkd physically interacts with Dsh, which plays a vital role in both canonical and noncanonical Wnt pathways [21].
Beyond its role in development, the function of Nkd proteins is highly conserved and critically important in human disease. In mammalian species, including humans, there are two NKD homologs, NKD1 and NKD2 [22], which have emerged as important regulators in cancer biology. Studies have identified and suggested that both NKD1 [23, 24, 25] and NKD2 [26, 27, 28] are associated with several human cancers, including colorectal adenocarcinoma and non‐small‐cell lung cancer, by mutations and DNA hypermethylation mediated downregulation. Loss of either NKD1 or NKD2 would promote tumorigenesis and/or result in a poor prognosis of cancer. These studies lead to a potential diagnostic application that the methylation or expression of NKD1/2 might be used as a prognostic marker in cancer. These studies underscore the evolutionary significance of Nkd family proteins and highlight the necessity of understanding their precise molecular functions, which can be elegantly modeled in Drosophila. It is important to understand the pleiotropic role of nkd as the connection between nkd homologs and human diseases.
In this study, we demonstrate that nkd impacts multiple biological processes and mediates cross talk between Wnt/Wg signaling and other development‐related pathways during Drosophila wing development, largely dependent on Nkd‐Dsh interaction.
Materials and methods
Drosophila genetics
P[UAS‐GFP], P[UAS‐mCD8::GFP] [29], P[UAS‐GFP::lacZ.nls] [30], P[UAS‐Nkd ^ GFPC ^], P[UAS‐Nkd ^ ΔR1S/GFPC ^], P[UAS‐Nkd ^ Δ30aa/GFPC ^] [17], P[UAS‐Nkd ^ ΔD6/GFPC ^] [19], and P[dpp‐LacZ] [31] have been described previously and were obtained from the Bloomington Stock Center. All UAS‐Nkd transgenic lines were systematically outcrossed into a common, uniform genetic background for at least five generations. Following outcrossing, stable stocks for each transgene were established through re‐balancing with the appropriate chromosome balancers.
All lines were cultured with standard medium at 25 °C. Experiments with Daughterless‐Gal4, Engrailed‐Gal4, Apterous‐Gal4, and MS1096‐Gal4 were carried out at 25 °C. Larvae were dissected 120 h after egg laying. Adult flies were frozen overnight and photographed with a Nikon ShuttlePix P‐400Rv microscope.
Antibodies
Immunostaining of imaginal disks was performed as described [32] using the following antibodies: anti‐β‐galactosidase antibodies (1 : 500, Cappel, MP Biomedicals, Santa Ana, CA, USA) and anti‐Wingless (1 : 200, Developmental Studies Hybridoma Bank at the University of Iowa, USA).
Vectors
Nkd alleles expression vectors were generated by subcloning Nkd fragments with GFP tags at the N terminus into a pAc5.1 expression vector (Invitrogen, Carlsbad, CA, USA). The primer sequences used for plasmid construction are listed below:
Nkd‐full forward: 5′‐GGTACCATGGCGGGTAACATTGTCAAATG‐3′, Nkd‐full reverse: 5′‐CTCGAGGACATCCTGCTGCTCCTTG‐3′, Nkd‐del‐R1S forward: 5′‐CTGGAGGAATTCTCCCGGGCGGAGCAGTGC‐3′, Nkd‐del‐R1S reverse: 5′‐CTCCGCCCGGGAGAATTCCTCCAGTCGGAT‐3′, Nkd‐del‐D6 forward: 5′‐AACCATCAAATGGCCTGCCCGAATCGCCAT‐3′, Nkd‐del‐D6 reverse: 5′‐ATTCGGGCAGGCCATTTGATGGTTCGGGTG‐3′, Nkd‐del‐30aa forward: 5′‐CGAAAATCGGCCGGAAAACCCCAAGCCAAT‐3′, Nkd‐del‐30aa reverse: 5′‐TTGGGGTTTTCCGGCCGATTTTCGCTGCTG‐3′.
Drosophila cell culture
Kc167 (Kc) cells were routinely cultured in Schneider's Drosophila media (Invitrogen) containing 5% Fetal Bovine Serum (FBS) at 25 °C. Transient transfections were carried out with X‐tremeGENE HP DNA Transfection Reagent (Roche Applied Science, Penzberg, Germany) according to the manufacturer's instructions.
For the Wnt response element (WRE) reporter assay, a mixture of DNA containing 100 ng SuperTopFlash (STF) vectors, 200 ng of ectopic Nkd alleles expression vectors, and 1 ng pAc‐lacZ (Invitrogen) was co‐transfected into 10^6^ cells. Transfection efficiency was normalized using the pAc‐lacZ β‐galactosidase activities. Luciferase and β‐galactosidase activities were assayed using the Tropix Luc‐Screen and Galacto‐Star kits (Applied Biosystems, Carlsbad, CA, USA) and quantitated with a Chameleon plate luminometer (Hidex Personal Life Science, Turku, Finland).
pTub‐wg S2 cells were kindly provided by Dr Roel Nusse from Stanford University. pTub‐wg S2 cells were cultured in Schneider's Drosophila media (Invitrogen) containing 10% FBS at 25 °C. Wg‐conditioned media (Wg‐CM) was concentrated and stored as described [14]. Kc cells were treated with Wg‐CM for 12 h prior to harvesting.
RNA sequencing analysis
Each RNA sample was prepared in biological duplicate from ~ 70 wing imaginal disks of late third larvae expressing GFP or Nkd recombinant proteins under the control of MS1096‐Gal4, which has been shown to express GAL4 in the central pouch region of wing disks [33, 34, 35]. Total RNA was extracted using TRIzol (Invitrogen). RNA‐seq libraries were generated using NEBNext UltraTM RNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA) following manufacturer's recommendations. RNA‐seq was performed by using Illumina NovaSeq 6000 platform and 150‐bp paired‐end reads were generated. Paired‐end clean reads were aligned to the fly reference genome (BDGP6) with hisat2 (version 2.2.1) [36]. The aligned reads were used to quantify mRNA expression by using featureCounts. The deseq2 package in r [37] was used to compute normalized fold change and adjusted P value for each gene, comparing Nkd recombinant protein‐expressing wing disks to the GFP‐expressing controls. Those genes with |fold change| > 1.5 and adjusted P value < 0.05 were considered to be differentially expressed. For bioinformatic analyses, the volcano plot showing differentially expressed genes (DEGs) was generated with ggplot2 package in r. Gene Ontology (GO) enrichment analysis of those DEGs was performed using david v6.8 [38, 39] and cytoscape v3.6.1 [40] with a bingo plugin (v3.0.3) [41, 42]. Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses were carried out with online software kobas 3.0 [43]. The RNA‐seq reads used for this study have been submitted to the National Center for Biotechnology Information (NCBI) under Database ID: GSE167145.
Results
Overexpression of mutant Nkd proteins suppresses Wg pathway in Kc167 cells
In order to gain further insight into the Nkd function and subcellular localization, we sought to use several published UAS alleles of nkd, including a full‐length wild‐type Nkd, P[UAS‐Nkd] [17], and three mutant alleles of Nkd. Nkd ^ΔR1S^ lacks the Dsh binding region [16]. Nkd ^Δ30aa^ and Nkd ^ΔD6^ have potential nuclear localization signals (NLSs) deleted [17, 19] (Fig. 1A). Using the Drosophila salivary gland system, we examined the subcellular localization of Nkd mutants. These mutants were ubiquitously overexpressed using the Daughterless‐Gal4 (Da‐Gal4) driver. Membrane‐localized and nuclear‐localized GFP served as controls (Fig. 1B). Wild‐type Nkd accumulated around the nuclear periphery with partial intranuclear puncta. Both Nkd ^Δ30aa^ and Nkd ^ΔD6^ localized cytoplasmically, while Nkd ^ΔR1S^ predominantly accumulated in the nucleus. This indicates that Nkd nuclear localization requires both putative NLSs [19]. Notably, the Nkd ^ΔR1S^ mutant exhibits preferential nuclear accumulation. Due to limitations of larval salivary glands (polytene, nondiploid cells) for functional insights, we generated Nkd mutant constructs based on in vivo fragments and investigated their function in Drosophila Kc cells via reporter assays.
Nuclear localization of Nkd in the salivary gland requires the D6 motif and a 30 amino acid fragment. (A) A diagram showing the domain structures of ectopic Nkd mutants to be overexpressed in flies. (B) Representative confocal images of the late third instar salivary gland cells with genotypes as indicated. Nuclei are stained with DAPI (blue). Scale bar is 20 μm. (C) SuperTopFlash reporter assay measuring Wg pathway activity. Kc cells treated with Wg‐CM show suppressed luciferase activity across all Nkd mutants. Experiments were performed in triplicate and repeated at least three times. The data are expressed as mean ± SEM. * indicates P < 0.05 compared with the control group, as analyzed by ANOVA, followed by Tukey's multiple comparison post hoc test.
The WRE serves as an indirect indicator of cellular Wg signaling intensity. SuperTopFlash (STF) comprises 12 tandem TCF‐binding sites [14]. Upon Wg pathway activation, this reporter plasmid drives luciferase expression, allowing semi‐quantitative measurement of Wg signaling via luciferase activity. Treatment with Wg‐conditioned media (Wg‐CM) activated the Wg pathway in GFP‐overexpressing control cells. Reporter assays revealed that all Nkd mutants suppressed Wg signaling upon Wg‐CM stimulation (Fig. 1C).
However, reporter assays may arise discrepancies with variable expression levels and differential cellular contexts. Adult wing phenotypic analysis in vivo may yield more biologically meaningful insights.
Nkd overexpression modulates wing morphogenesis
The crumpled wings and necrotic‐like appearance were frequently observed when either Nkd or mutant alleles were driven by Apterous‐Gal4 (Ap‐Gal4) in the ventral wing pouch (Fig. 2A). In addition to morphological alterations, the wings of mutant flies exhibited size reduction compared with those of Ap‐Gal4 > GFP controls.
Nkd overexpression modulates wing morphogenesis without altering Dpp expression pattern. (A) Representative images of female adult wings from flies with genotypes as indicated. Scale bar is 500 μm. (B) Representative confocal images of the third instar larval wing disks with antibodies against β‐galactosidase (LacZ, red) with genotypes as indicated. Dorsal/ventral boundaries are marked by the white dotted lines. Nuclei stained with DAPI (blue). Scale bar is 100 μm.
During development, graded morphogens regulate gene expression across fields of cells in a concentration‐dependent manner [44]. Ectopic overexpression or insufficient expression of Decapentaplegic (Dpp) correspondingly alters imaginal disk size [45]. Given the established correlation between Dpp signaling strength and wing size, we investigated whether observed wing size variations result from altered Dpp expression levels. In our experiments, ectopic Nkd expression in the Drosophila wing disk induced no significant change in Dpp expression pattern (Fig. 2B). This suggests that Dpp occupies a position upstream of Nkd within the wing developmental regulatory network. These results prompted us to hypothesize that the role of nkd function in wing development might be beyond Wingless signaling, consistent with our understanding that Drosophila wing development is orchestrated by many signaling pathways.
Overproduction of Nkd disrupts transcriptional networks in wing disks
To explore the nkd function in the wing at a molecular level, we examined the whole transcriptional profiles of wing disks expressing Nkd mutants and GFP controls by performing RNA sequencing (RNA‐seq) analysis. MS1096‐Gal4 was reported to express GAL4 in the central pouch region of third instar larval wing disks [33, 34, 35] (Fig. 3A), which could induce Nkd expression in a broader manner than En‐Gal4 and Ap‐Gal4 do. RNA samples were extracted from dissected wing primordia expressing Nkd proteins driven by MS1096‐Gal4 (see Materials and methods).
Overproduction of Nkd in wing disks affects biological processes in larvae. (A) Representative confocal images of the late third instar wing imaginal disks stained with antibodies against Wingless (Wg, blue) with genotypes indicated. Scale bar is 100 μm. (B) The differentially expressed genes (DEGs) were illustrated in volcano plots (|fold change| > 1.5 and adjusted P value < 0.05). Data points in red represent upregulated and green represent downregulated genes. Genes without any significant difference are in black. (C) Functional annotation clustering analysis of differentially expressed genes derived from Nkd overexpression was performed using DAVID. (D) Gene Ontology (GO) charts for Biological Processes of DEGs between Nkd group and GFP control, analyzed and visualized as directed acyclic graphs by Cytoscape. GO categories are shown in circles with the area proportional to the ratios of observed gene frequencies over expected ones and the intensities of the orange color gradient correlate with minus Log10(P values).
The DEGs between Nkd group and GFP control (Nkd_vs._GFP) include 418 upregulated genes and 254 downregulated genes (|fold change| > 1.5 and adjusted P value < 0.05) (Fig. 3B). Overexpression of Nkd significantly impacts the expression of multiple genes involved in wing morphogenesis. Functional annotation clustering analysis revealed significant enrichment of biological processes critical for Drosophila wing development, including imaginal disk‐derived wing morphogenesis and wing vein specification. Key transcriptional regulatory pathways, such as DNA‐templated transcription, were concurrently enriched, suggesting coordinated genetic control of wing patterning (Fig. 3C). Gene Ontology analysis performed by Cytoscape detailed these DEGs in some interesting biological processes, such as postembryonic organ development, cell cycle, neuron differentiation, and stem cell maintenance (Fig. 3D). These processes play crucial roles in Drosophila wing development, determining wing shape, vein patterning, and transcriptional regulation.
R1S region is essential for Nkd‐mediated signaling cross talk
For better understanding of Nkd function in biological processes, we analyzed DEGs between different Nkd mutant groups and GFP control with the same threshold. We processed DEGs in each group with david functional category analysis and found that these genes were enriched in imaginal disk derived wing morphogenesis, neurogenesis as well as other biological processes, including chromatin organization (Fig. 4A), which suggests that Nkd participates in wing development from many directions directly or indirectly.
Overproduction of Nkd in wing disks disturbs multiple biological processes and signalings involved in Drosophila development. (A) Top biological process terms enriched in DAVID functional Category analysis of DEGs in different groups. n.s., not significant. (B) The bubble chart showing enriched DEGs in KEGG signaling pathway for different groups. The color and size of the bubble represent P value and the percentage of differentially expressed genes enriched in the pathway, respectively.
The expression of some key components in Wnt/Wg signaling and/or other pathways has changed significantly when Nkd is overproduced. KEGG pathway analysis revealed that these differentially expressed genes were highly involved in several critical pathways, including Hippo, Notch, mitogen‐activated protein kinase (MAPK), and Wnt signaling pathways (Fig. 4B and Table 1). While Nkd ^ΔR1S^ overexpression altered Hippo signaling, it minimally affected Wnt, Notch, and MAPK pathways or processes like wing morphogenesis compared with wild‐type Nkd (Fig. 4B). This result could be attributed to the limited number of DEGs in the Nkd^ΔR1S^_vs._GFP group (116 DEGs). These data prompt us to the conclusion that the R1S region is crucial for Nkd in development‐related signaling pathways, not merely Wnt signaling. The mediated role of Nkd in cross talk between Wnt/Wg signaling and other development‐related pathways in Drosophila largely depends on the R1S region.
Discussion
Wnt signaling is most famous for the ability to shape growing tissues while inducing proliferation in development [1, 3]. Wnt signaling should be finely balanced as it is crucial for the development and maintenance of stem cells. Nkd proteins play an essential role in Wnt/Wg signaling as a negative‐feedback regulator.
Our research demonstrates that Nkd exerts pleiotropic effects on Drosophila wing development, extending beyond its canonical role as a Wnt/Wg signaling inhibitor. Nkd overexpression induced wing size reduction and crumpling, which indicates there are more features for nkd or its homologs beyond inhibiting Wg signaling [46, 47, 48, 49].
Interestingly, while ectopic Nkd expression induced a pronounced reduction in wing size, it did not alter the expression pattern of Dpp, a key morphogen governing wing growth. This indicates that the size defect is not mediated through the canonical Dpp signaling gradient. Instead, we propose that the observed growth impairment likely stems from the broad transcriptional perturbations revealed by our RNA‐seq analysis. Specifically, the significant enrichment of DEGs related to cell cycle suggests a mechanism whereby Nkd overexpression may directly impact cell proliferation or survival. Furthermore, the profound disruption of the Hippo signaling pathway presents a particularly compelling alternative mechanism, as Hippo is a well‐established central regulator of organ size that acts in parallel to Dpp. Future studies quantifying cell number, cell size, and apoptosis in the wing disk upon Nkd overexpression will be crucial to precisely distinguish between these possibilities.
RNA‐seq analysis revealed widespread transcriptional perturbations in wing disks overexpressing Nkd, with significant enrichment of genes linked to Hippo, Notch, MAPK, and Wnt pathways. Beyond its developmental implications, our work provides new mechanistic insights into the tumor‐suppressive role of Nkd homologs in human cancers. The observation that Nkd‐mediated cross talk critically depends on the R1S region redefines Nkd proteins from mere Wnt pathway inhibitors to potential signaling integrators. This expanded role has direct implications for diagnostic and therapeutic strategies. For diagnosis, the downregulation or mutation of NKD1/2, as observed in carcinomas, likely leads to a concurrent dysregulation of a network of oncogenic pathways [23, 24, 25, 26, 27, 28]. This could explain the association of NKD loss with poor prognosis and suggests that NKD status could serve as a more comprehensive biomarker for tumor multi‐pathway activity. Therapeutically, our data suggest that restoring the function of NKD may represent a novel strategy to simultaneously dampen multiple oncogenic signals. Future efforts could focus on identifying small molecules that stabilize the NKD‐DVL interaction or otherwise enhance NKD integrative activity, offering a potential multi‐pathway intervention for Wnt‐dysregulated cancers.
It is important to note that the broad transcriptional changes observed upon Nkd overexpression could encompass both direct regulatory targets and secondary consequences of developmental disruption. Future studies employing tissue‐specific knockdown and direct measurement of protein levels of key pathway components will be essential to validate and extend these transcriptomic findings. The DEGs identified here should be considered as a resource that defines the ‘Nkd‐overexpression signature’ and implicates specific pathways. Future work employing tissue‐specific, acute protein degradation systems combined with assays for direct binding will be essential to disentangle the direct targets from the secondary network effects and precisely map the domains responsible for specific interactions. Although variable expression levels of Nkd transgenes preclude quantitative comparison between mutants, the Nkd^ΔR1S^ allele uniquely failed to perturb Notch/MAPK pathways (Fig. 4B). This specific phenotype, which coupled with its nuclear accumulation, highlights the R1S region as a critical determinant for cross talk. The conserved role of R1S in Dsh binding [15, 16] suggests that the integrator function of Nkd may be evolutionarily wired, positioning it as a therapeutic node for Wnt‐dysregulated diseases.
Our data propose that Nkd homologs may serve as signaling nodes integrating multiple pathways. The multifaceted role of Nkd in Drosophila wing development highlights its capacity to integrate Wnt signaling with other pathways. These findings expand the functional repertoire of Nkd beyond canonical Wnt inhibition and provide a framework for investigating Nkd homologs in developmental and disease contexts.
Conflict of interest
The authors declare no conflict of interest.
Author contributions
RW conceived and designed the project. RW and PW conducted experiments and analyzed results. RW wrote the paper. Both authors reviewed and approved the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Loh KM , van Amerongen R and Nusse R (2016) Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Dev Cell 38, 643–655.27676437 10.1016/j.devcel.2016.08.011 · doi ↗ · pubmed ↗
- 2Beumer J and Clevers H (2024) Hallmarks of stemness in mammalian tissues. Cell Stem Cell 31, 7–24.38181752 10.1016/j.stem.2023.12.006PMC 10769195 · doi ↗ · pubmed ↗
- 3Nusse R and Clevers H (2017) Wnt/β‐catenin signaling, disease, and emerging therapeutic modalities. Cell 169, 985–999.28575679 10.1016/j.cell.2017.05.016 · doi ↗ · pubmed ↗
- 4Kinzler KW and Vogelstein B (1996) Lessons from hereditary colorectal cancer. Cell 87, 159–170.8861899 10.1016/s 0092-8674(00)81333-1 · doi ↗ · pubmed ↗
- 5Wood LD , Parsons DW , Jones S , Lin J , Sjöblom T , Leary RJ , Shen D , Boca SM , Barber T , Ptak J et al. (2007) The genomic landscapes of human breast and colorectal cancers. Science 318, 1108–1113.17932254 10.1126/science.1145720 · doi ↗ · pubmed ↗
- 6Johnson DE and Disis ML (2025) Vaccines for cancer interception in familial adenomatous polyposis. Front Immunol 16, 1525157.39944699 10.3389/fimmu.2025.1525157 PMC 11813938 · doi ↗ · pubmed ↗
- 7Liu W , Dong X , Mai M , Seelan RS , Taniguchi K , Krishnadath KK , Halling KC , Cunningham JM , Boardman LA , Qian C et al. (2000) Mutations in AXIN 2 cause colorectal cancer with defective mismatch repair by activating beta‐catenin/TCF signalling. Nat Genet 26, 146–147.11017067 10.1038/79859 · doi ↗ · pubmed ↗
- 8Morin PJ , Sparks AB , Korinek V , Barker N , Clevers H , Vogelstein B and Kinzler KW (1997) Activation of beta‐catenin‐Tcf signaling in colon cancer by mutations in beta‐catenin or APC. Science 275, 1787–1790.9065402 10.1126/science.275.5307.1787 · doi ↗ · pubmed ↗