The photoswitchable cannabinoid azo‐HU308 enables optical control of Ca2+ dynamics in INS‐1 β‐cells via off‐target effects on TRPC channels
Alexander E. G. Viray, James A. Frank

TL;DR
A light-sensitive cannabinoid compound triggers calcium influx in pancreatic cells through TRPC channels, not via its intended receptor.
Contribution
Azo-HU308 is a novel optical tool for controlling calcium levels in β-cells via TRPC channels, bypassing CB2 receptors.
Findings
UV-A activation of azo-HU308 induces repeatable Ca2+ transients in INS-1 β-cells.
The effect is mediated by TRPC channels and extracellular Ca2+ influx, not CB2 receptor activation.
Abstract
Intracellular Ca2+ regulates insulin secretion from pancreatic β‐cells and is influenced by cannabinoid signaling. However, the hydrophobicity and complex pharmacology of cannabinoid ligands prevent precision receptor targeting, limiting our understanding of their roles in modulating insulin release. Here, we use fluorescent Ca2+ imaging to examine how the light‐activatable CB2 receptor agonist azo‐HU308 modulates Ca2+ dynamics in INS‐1 β‐cells. UV‐A photoactivation of azo‐HU308 triggered robust, repeatable Ca2+ transients, and pharmacological profiling revealed this effect was independent of CB2 receptor activation but was instead mediated by extracellular Ca2+ influx through TRPC channels. These findings position azo‐HU308 as a novel optical tool for controlling β‐cell Ca2+ levels and highlight a non‐GPCR pathway by which synthetic cannabinoids can modulate Ca2+ dynamics in excitable…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
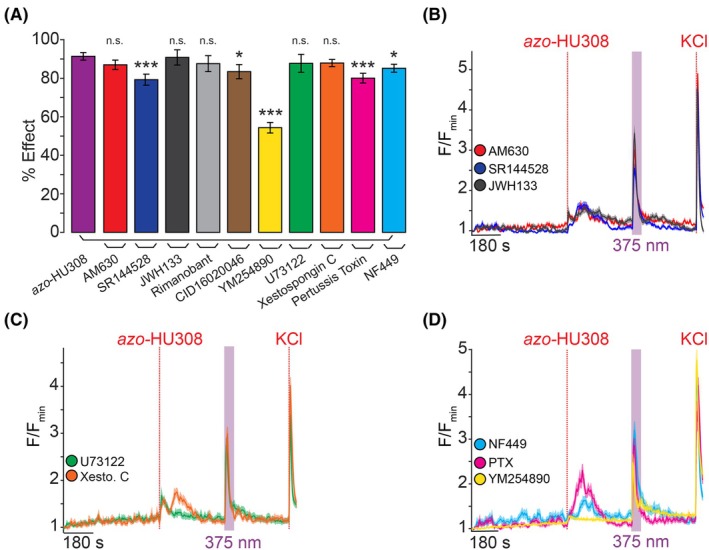 Fig. 2
Fig. 2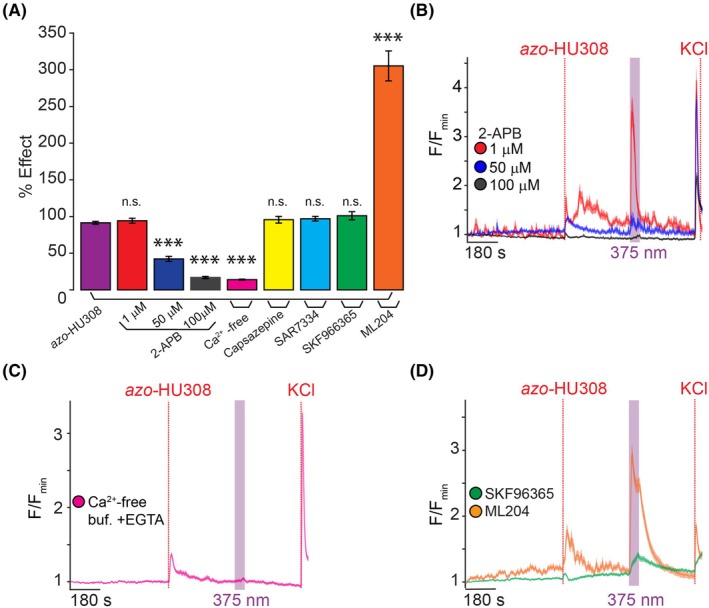 Fig. 3
Fig. 3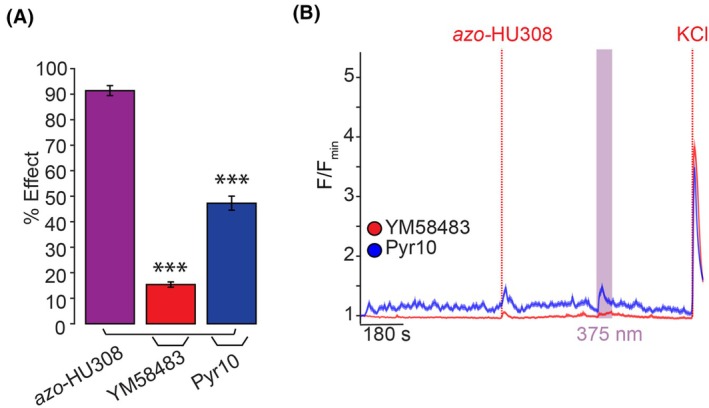 Fig. 4
Fig. 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPancreatic function and diabetes · Cannabis and Cannabinoid Research · Ion Channels and Receptors
Ca^2+^ regulates intracellular signaling and the secretion of hormones and neurotransmitters from excitable cells. In pancreatic β‐cells, Ca^2+^ controls the release of insulin, a key hormone that regulates glucose homeostasis [1]. After a meal, blood glucose is elevated and is transported into the β‐cell where it is metabolized to ATP. This closes ATP‐sensitive K^+^ channels (K_ATP_) and depolarizes the β‐cell, triggering Ca^2+^ influx through voltage‐dependent Ca^2+^ channels and subsequent insulin release. In addition, β‐cell Ca^2+^ is regulated by myriad receptor pathways, including ion channels and G protein‐coupled receptors (GPCRs). These Ca^2+^ fluctuations are oscillatory in nature, and their intensity and frequency correlate with insulin release [2]. Dysfunction in β‐cell Ca^2+^ signaling and glucose‐stimulated insulin secretion (GSIS) leads to type 2 diabetes mellitus (T2DM), a disease whose prevalence is rising throughout the world at epidemic rates. As such, novel approaches to modulate β‐cell Ca^2+^ levels and insulin release are urgently needed to address this epidemic.
Small‐molecule therapeutics that treat diabetes by promoting insulin release include sulfonylureas, which block K_ATP_ channels [3], and approaches that modulate the β‐cell's intracellular Ca^2+^ concentration ([Ca^2+^]i) through alternative receptor pathways, including GPCR targets, have gained significant therapeutic interest [4]. For example, the GPR40 agonist TAK‐875 improved glycemic control in patients but was pulled from clinical trials due to toxicity concerns [5]. More recently, glucagon‐like peptide‐1 receptor (GLP1R) agonists have demonstrated improved safety for treating metabolic disease and are increasingly being prescribed for T2DM [6]. Importantly, β‐cells express nearly 300 GPCRs [7], and these represent unexplored therapeutic targets that could eventually be leveraged as new therapeutic targets for T2DM.
One such GPCR family is the cannabinoid receptors (CBRs), which are activated by endocannabinoid lipids and phytocannabinoids found in marijuana. Cannabinoid CB1 and CB2 receptors (CB1R and CB2R) are expressed in the human pancreas and affect β‐cell excitability [8]. Additionally, noncanonical ‘orphan’ CBRs—such as GPR55 [9, 10]—affect insulin release from β‐cells in vitro and in rodent models [11]. Unfortunately, cannabinoid‐based therapeutics have had difficulty translating to the clinic, likely because we have a limited understanding of how CBRs affect β‐cell function. This is due to the hydrophobic nature of cannabinoid ligands, which are insoluble in physiological solutions and makes their application to cells slow and irreversible [12]. Cannabinoids also have complex pharmacology, and act through multiple GPCR and ion channel targets [13, 14]. To deconvolute the precise roles of CBR signaling in modulating β‐cell excitability and leverage this important signaling system for therapeutic development, we require tools that enable greater spatiotemporal resolution over cannabinoid signaling pathways.
Photopharmacology is an emerging approach to address these challenges by enabling optical control of biological processes with light‐sensitive small molecules, including photocaged and photoswitchable probes [15]. Photocaged ligands have their activity blocked by a photo‐labile protecting group, where a flash of light irreversibly releases the active compound. Photocages for Ca^2+^ and other ligands, including endocannabinoids [10, 16, 17], have been applied to optically control β‐cell [Ca^2+^]i [18]. Alternatively, photoswitchable probes are ligands that contain a photo‐isomerizable motif, most commonly an azobenzene, that switches between the trans‐ and cis‐conformations using two different wavelengths of light [19]. The change in molecular structure through cis/trans isomerization tunes the potency and affinity of the ligand, enabling reversible optical control of receptor activation with superior kinetics compared to standard pharmacology. Numerous photoswitches—including sulfonylureas or endogenous lipids—have been shown to enable dynamic manipulation of [Ca^2+^]i in cultured β‐cells and pancreatic islets [20, 21, 22, 23, 24]. Although our group and others have developed an array of photoswitchable cannabinoids to modulate both CB1R [25, 26, 27] and CB2R [28, 29], to date, they have not been applied to β‐cells.
Recently, our group developed a photoswitchable ligand based on the CB2R‐selective synthetic cannabinoid, HU308 [30]. We incorporated an azobenzene into HU308's alkyl chain to afford a photoswitchable ligand, azo‐HU308‐3 (now coined azo‐HU308) (Fig. 1A). Using a Ca^2+^ imaging assay in AtT‐20 cells overexpressing CB2R [AtT‐20(CB2) cells], we demonstrated that azo‐HU308 enables reversible, optical control of [Ca^2+^]i when photoswitched to cis, and that the mechanism proceeded by CB2R and phospholipase C, and subsequent Ca^2+^ release from intracellular stores. As a follow‐up to this study, here we present the application of azo‐HU308 to optically control [Ca^2+^]i in cultured pancreatic β‐cells. We show that azo‐HU308 robustly elevates intracellular Ca^2+^ in β‐cells when isomerized to cis with UV‐A light and permits multiple rounds of stimulation. Using a pharmacological screen to determine the mechanism of action, we observed that the Ca^2+^ influx was not CB2R‐mediated, but rather involves TRPC ion channels and Ca^2+^ influx across the plasma membrane. This study presents a novel tool to optically modulate β‐cell Ca^2+^ dynamics and sheds light on a new mechanism by which synthetic cannabinoids may modulate Ca^2+^ influx through non‐GPCR targets.
Materials and methods
Compounds
Azo‐HU308 (azo‐HU308‐3) was synthesized as previously described [31]. HU308 (Cayman Chemical, Ann Arbor, MI, USA; #90086), AM630 (Cayman Chemical, #1006974), SR 144528 (Cayman Chemical, #9000491), JWH 133 (Cayman Chemical, #10005428), Rimonabant hydrochloride (Sigma‐Aldrich, St. Louis, MO, USA; #SML0800), CID16020046 (Tocris, Bristol, UK; #4959), Xestospongin C (Cayman Chemical, #64950), U73122 (Tocris, #1268), YM254890 (Tocris, #7352), Pertussis Toxin (Cayman Chemical, #19546), NF449 (Cayman Chemical, #13324), 2‐APB (Tocris, #1224), Capsazepine (Cayman Chemical, #10007518), SAR7334 hydrochloride (Cayman Chemical, #28292), SKF 96365 hydrochloride (Cayman Chemical, #10009312), ML‐204 (Cayman Chemical, #15626), YM‐58483 (Cayman Chemical, #13246), and Pyr 10 (Tocris, #6941) were obtained from commercial suppliers. Compounds were dissolved as stock solutions in DMSO, aliquoted, and stored at −20 °C until use.
Cell culture media and solutions
INS‐1 media contains: RPMI 1640 with l‐glutamine (Gibco, Waltham, MA, USA; #11875‐093) with 10% FBS, 1 : 100 Penicillin‐strep (5000 units·mL^−1^, Gibco, #15070), and 10 mm HEPES (Fisher, Waltham, MA, USA; #BP310‐500), 1 mm sodium pyruvate (Alfa Aesar, Ward Hill, MA, USA; #A11148). INS‐1 media was filtered and distributed into 50 mL aliquots. 50 μm 2‐mercaptoethanol diluted in H_2_O (BME, Sigma, #M3148) was added fresh to each 50 mL aliquot prior to use.
INS‐1 imaging buffer contains (in mm): 185 NaCl, 1.2 CaCl_2_, 1.2 MgCl_2_, 1.2 K_2_HPO_4_, 20 HEPES. Adjusted to pH 7.4 with NaOH. d‐glucose was supplemented at 20 mm.
Phosphate buffered saline (PBS) contains (in mm): 320 Na_2_HPO_4_ (Fisher, #BP332‐500), 80 Na(PO_4_H_2_)·H_2_O (Fisher, #S369‐1). Adjusted pH to 7.4 with NaOH.
Cell culture
INS‐1 832/13 cells [32] were grown in INS‐1 media and incubated at 37 °C and 5% CO_2_. Cells were used between passages 65 and 80. For live‐cell Ca^2+^ imaging, INS‐1 cells were plated at a density of 100,000 cells per well on 8‐well glass bottom chambered coverslips (Ibidi, Graefelfing, Germany; #0827‐90 or Cellvis, Mountain View, CA, USA; #C8‐1.5H‐N). 18–24 h later, the cells were starved in Opti‐MEM™ (250 μL) for 2 h before adding the transfection mixture containing (per well): 50 μL Opti‐MEM™, 1 μL Lipofectamine‐2000 (Fisher Scientific, #11668019), and 250 ng R‐GECO cDNA. The cells were then incubated at 37 °C and 5% CO_2_ for 18–24 h before exchanging the transfection mixture with INS‐1 media. Microscopy experiments were performed 60–72 h post transfection.
Confocal microscopy
Live cell imaging was performed on an Olympus Fluoview 1200 laser scanning confocal microscope at 37 °C and 5% CO_2_. Videos were acquired with a 20×/0.75 NA objective (Olympus UPlanSApo, Center Valley, PA, USA) at 512 × 512‐pixel resolution and a scan rate of 4 s/frame. R‐GECO excitation was performed with a 559‐nm laser at low laser power (1–3%), and emission was collected at 570–670 nm. Compounds stock solutions (1000× in DMSO) were incubated for 10 s with 10% Pluronic F‐127 (Tocris #6253) in DMSO (1 : 1 v:v ratio with compound solution). The mixture was then diluted 1000× with imaging buffer and pipetted directly into the imaging well. Vehicle controls were performed using an identical procedure, adding a final concentration of DMSO (0.2 vol%) and Pluronic F‐127 (0.01 vol%). For co‐application experiments with receptor antagonists, the drugs were applied to the imaging buffer and fluorescent images were acquired immediately upon thermal equilibration of the plate in the environment chamber, typically ~ 10 min after application.
Photoswitching was performed with a 375‐nm laser (PicoQuant, Berlin, Germany; PDL 800‐D, ~ 95 μW output from the objective) triggered using the quench function in the olympus software. KCl (30 mm) was added at the end of each experiment to elicit maximum R‐GECO fluorescence intensity for normalization. Fluorescence intensity values in each experimental video were collected and analyzed with imagej [33] and the data were processed and plotted in Excel and matlab using in‐house written scripts (available on request). Oval‐shaped regions were manually drawn over each cell to measure the fluorescence in imagej.
Data analysis and statistical methods
All data are presented as mean ± SEM For Ca^2+^ imaging experiments, the figure captions report the total number of individual cells (N) and the number of independent trials (T, biological replicates). Statistical significance was assessed using the Data Analysis package in matlab (Mathworks, Natick, MA, USA). For comparison between two groups, a Student's paired t‐test (two‐sided) was used, with the significance threshold placed at *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant.
For bar graph analysis of R‐GECO intensity (e.g., Fig. 1E), the photoswitching ‘Percent effect (%)’ values were calculated as change in fluorescence intensity within 60 s after addition of the trans isomer; or for cis‐, the maximum intensity during irradiation. These values are normalized to the range calculated between minimum baseline fluorescence intensity (first 50 frames) and the maximum intensity generated after KCl addition. These values were calculated for each cell, then calculated as a mean across all cells and plotted as a bar graph for comparison between conditions.
Results
Azo‐HU308 enables reversible optical control of Ca2+ influx in INS‐1 pancreatic β‐cells
Motivated by studies that showed that CBR signaling modulates Ca^2+^ in β‐cells [17, 34] and our finding that azo‐HU308 reversibly controls Ca^2+^ levels via CB2R and Ca^2+^ release from intracellular stores [31], we asked whether azo‐HU308 could optically modulate [Ca^2+^]i in cultured β‐cells. We chose the rat insulinoma INS‐1 832/13 (INS‐1) cells because they exhibit glucose‐stimulated Ca^2+^ oscillations that couple to insulin release [32], and little is known about CBR signaling in INS‐1 cells. INS‐1 cells were transfected with the genetically encoded Ca^2+^ sensor R‐GECO [35], which allowed us to monitor [Ca^2+^]i levels in real time by fluorescence microscopy (Fig. 1B, top). We observed robust Ca^2+^ oscillations when the cells were exposed to elevated glucose (20 mm) solution (Fig. 1C). Application of trans‐azo‐HU308 did not affect [Ca^2+^]i (Fig. 1C and 1D, red); but strikingly, when we irradiated the cells with 375 nm UV‐A light (~ 95 μW laser irradiation) to isomerize azo‐HU308 to cis, we observed a robust [Ca^2+^]i burst across the cells (Fig. 1B bottom, C, D, red, E, Fig. S1A and Table S1). The Ca^2+^ increase was intense but short‐lived and decayed rapidly before the photostimulation period was finished. The effect of azo‐HU308 was dose‐sensitive, and the [Ca^2+^]i‐stimulating effect continued to increase up to 20 μm (Fig. S1B,C and Table S2). In control experiments, the parent molecule HU308 [30] caused a similar Ca^2+^ response immediately upon addition. The Ca^2+^ response induced on HU308 application quickly died off, and there was no further Ca^2+^ effect induced by 375 nm irradiation, confirming that the parent compound HU308 is not light‐sensitive (Fig. 1E, Fig. S2A–D and Table S1). In vehicle control experiments, where DMSO (0.1%) was added in the absence of azo‐HU308, there was no change to [Ca^2+^]i upon addition or irradiation (Fig. 1D, black). This confirms that the effect was not directly caused by the irradiation or an artifact of compound addition.
To test the reversibility of azo‐HU308's effects, 375‐nm photostimulation was repeated over six cycles. We observed a Ca^2+^ increase on each irradiation; however, the magnitude decayed on each subsequent stimulation, which indicates that receptor desensitization was likely occurring (Fig. 1F). To highlight the outstanding spatial precision of azo‐HU308, we applied targeted illumination of three different square regions of interest on the plate (Fig. 1G). We observed [Ca^2+^]i increases only in the cells that were irradiated, while cells that were outside the irradiation area did not respond (Fig. 1H). Combined, these results demonstrate that azo‐HU308 is a photoswitchable ligand that robustly stimulates Ca^2+^ in INS‐1 β‐cells when activated with UV light and isomerized to cis.
Azo‐HU308's effect on β‐cell Ca2+ is not mediated by CB2R
After verifying the robust effect of cis‐azo‐HU308 stimulating INS‐1 cell [Ca^2+^]i, we wanted to determine its mechanism of action. Our previous study showed that azo‐HU308 and HU308 elevate [Ca^2+^]i in AtT‐20(CB2) cells through CB2R and phospholipase C (PLC)‐mediated Ca^2+^‐release from intracellular stores [31]. Therefore, we co‐applied CB2R antagonists alongside azo‐HU308 to verify the contributions of CB2R signaling. To our surprise, azo‐HU308 still caused a large Ca^2+^ response in INS‐1 cells upon irradiation, even when co‐applied with three different CB2R antagonists—AM630, SR144528, and JWH133 (Fig. 2A,B and Table S3) [36, 37, 38]. Although HU308 is touted as a selective ligand for CB2R [30], it is possible that our probe has off‐target effects at other CBRs. INS‐1 cells express multiple CBRs including CB1R and GPR55 [10, 34]. However, co‐application of the CB1R antagonist rimonabant [39], or the GPR55 antagonist CID16020046 [40] failed to block azo‐HU308's photoswitching effect (Fig. 2A, Fig. S3A,B and Table S3). These results suggest that the effects of azo‐HU308 in β‐cells are not mediated by CB2R or through off‐target CBR pathways.
*Azo‐HU308's photoswitching effect in INS‐1 cells is not mediated by CB2R or G protein signaling. INS‐1 cells were transfected with R‐GECO and Ca2+ levels recorded by confocal microscopy. Azo‐HU308 (20 μm) and 375 nm light were applied in conjunction with various signaling antagonists. Shown are: (A) Summary bar graph showing that the % Ca2+ effect for azo‐HU308 photoswitching in the presence of CB2R inhibitors AM630 (20 μm; N = 151, T = 3), SR144528 (20 μm; N = 159, T = 3), and JWH133 (32 μm; N = 166, T = 3), which failed to block azo‐HU308's effect. Similar results were observed with the CB1R inhibitor Rimanobant (2 μm; N = 103, T = 2), GPR55 inhibitor CID16020047 (20 μm; N = 104, T = 2), PLC inhibitor U73122 (10 μm; N = 89, T = 2), or the IP3R inhibitor Xestospongin C (1 μm; N = 152, T = 3). Inhibitors for G protein signaling, including Gs‐protein inhibitor NF449 (10 μm; N = 173, T = 3), Gi/o‐protein inhibitor pertussis toxin (100 ng·mL−1; N = 135, T = 4), and Gq/11 protein inhibitor YM524890 (10 μm; N = 120, T = 3) also failed to block azo‐HU308 photoswitching. (B) Averaged Ca2+ traces for CB2R inhibitors AM630 (red), SR144528 (blue), and JWH133 (black). (C) Average traces for U73122 (green) and Xestospongin C (orange). (D) Average Ca2+ traces for G protein inhibitors: Gs‐inhibitor NF449 (cyan), Gi/o‐inhibitor pertussis toxin (magenta) and Gq/11‐inhibitor YM254890 (yellow). Error bars = mean ± SEM. *P < 0.05, **P < 0.01, **P < 0.001, ns, not significant. Student's t‐test.
Next, we asked whether PLC and Ca^2+^‐release from internal stores were being stimulated by cis‐azo‐HU308, as found in our previous study [31]. Surprisingly, both the PLC inhibitor U73122 and the inositol 1,4,5‐trisphosphate receptor (IP_3_R) inhibitor Xestospongin C failed to block the photoswitching effect (Fig. 2A,C), indicating that an entirely different Ca^2+^ signaling mechanism was involved [41, 42]. We tested several inhibitors of G protein signaling. The G_q/11_ G protein inhibitor YM254890 produced only a modest inhibition of the photoswitching effect, but the response was still largely intact (Fig. 2A,D, yellow) [43]. We applied the G_s_ protein inhibitor NF449 (Fig. 2A,D, cyan) and the G_i/o_ protein blocker pertussis toxin (PTX, Fig. 2A,D, magenta) [44, 45]. Both drugs caused only a small reduction of cis‐azo‐HU308's effect on [Ca^2+^]i. Combined, these results suggest that the mechanism of action for azo‐HU308's effect on [Ca^2+^]i release in INS‐1 cells was independent of CB2R, other CBRs, or even GPCR activation entirely. These results suggest an entirely unique mechanism compared to our previous study in CB2R‐overexpressing AtT‐20 cells.
Azo‐HU308's photoswitching effect is modulated by ion channel targets
Besides GPCRs, pancreatic β‐cell [Ca^2+^]i is affected by numerous transmembrane ion channels, including transient receptor potential (TRP) channels [46]. Based on the rapid and transient nature of azo‐HU308's effect, we hypothesized that these may be involved as an unintended receptor target. To this end, we tested the effect of 2‐aminoethoxydiphenyl borate (2‐APB), which is a nonselective TRP channel inhibitor and IP_3_ receptor modulator [47], and observed a dose‐dependent inhibition of azo‐HU308 photoswitching (Fig. 3A,B and Table S4). At low concentrations of 2‐APB (1 μm, red), we observed a potentiation of the effect of trans‐azo‐HU308, as a larger [Ca^2+^]i increase was observed upon probe addition, while the cis‐effect remained constant. At a mid‐range concentration (50 μm, blue), the photoswitching effect was significantly reduced, and at the highest concentration (100 μm, black), the effect of trans‐ and cis‐azo‐HU308 was completely abolished. We then conducted our Ca^2+^ imaging experiments in Ca^2+^‐free extracellular solution (with added ethylene glycol‐bis(β‐aminoethyl ether)‐N,N,N′,N′‐tetraacetic acid (EGTA)), where we observed that azo‐HU308's photoswitching effect on Ca^2+^ was completely abolished (Fig. 3A,C and Table S4). Combined, these results indicate that the Ca^2+^ response of cis‐azo‐HU308 is mediated by ion channels expressed on the INS‐1 cell surface, and that the rise in [Ca^2+^]i requires extracellular Ca^2+^ influx.
*Azo‐HU308's effect on Ca2+ is modulated by pharmacological inhibitors of TRP channels. INS‐1 cells were transfected with R‐GECO and Ca2+ levels recorded by confocal microscopy. Azo‐HU308 (20 μm) was applied in conjunction with various antagonists. (A) Summary bar graph showing a dose‐responsive photoswitching effect on Ca2+ with 2‐APB at different concentrations (1 μm; N = 100, T = 2), (50 μm; N = 93, T = 2), and (100 μm; N = 96, T = 2), and the response was completely abolished by the removal of extracellular Ca2+ using EGTA (0.1 mm) in the buffer (N = 200, T = 4). The TRPV1 blocker Capsazepine (5 μm; N = 119, T = 2), TRPC6 blocker SAR7334 (20 μm; N = 164, T = 3), broadband TRPC blocker SKF963665 (20 μm; N = 151, T = 3), and TRPC4 blocker ML204 (100 μm; N = 133, T = 3) failed to prevent the photoswitching effect. (B) Averaged Ca2+ traces showing dose‐dependent inhibition of photoswitching response with 2‐APB at 1 μm (red), 50 μm (blue), and 100 μm (black). (C) Averaged Ca2+ trace showing that the photoswitching effect requires an extracellular source of Ca2+ (magenta), as azo‐HU308 photoswitching had no effect in the presence of EGTA. (D) Averaged Ca2+ traces showing the effect of SKF96365 (green); and that ML204 (orange) potentiated photoswitching. Error bars = mean ± SEM. *P < 0.05, **P < 0.01, **P < 0.001, ns, not significant. Student's t‐test.
We then set out to determine the specific ion channels involved. INS‐1 cells express a variety of TRP channels, which are known to respond to lipid agonists [46, 48, 49, 50]. Transient receptor potential vanilloid 1 (TRPV1) is expressed in INS‐1 cells [51] and has been shown to be activated by lipids. However, when INS‐1 cells were pretreated with the TRPV1 antagonist capsazepine [52], azo‐HU308 photoswitching effects remained intact (Fig. 3A and Fig. S4A), ruling out TRPV1's role. Next, we set our sights into TRPC channels. Human and murine pancreatic islets and β‐cells, including INS‐1 cells, express multiple TRPC channels, which are important for store‐operated Ca^2+^ homeostasis and can regulate insulin secretion [53, 54]. Gratifyingly, when we pretreated with the broadband TRPC inhibitor SKF96365, there was a reduction in amplitude to the photoswitching effect of azo‐HU308 (Fig. 3A,D, green) [55]. We then applied more selective antagonists. Interestingly, co‐application of the TRPC4 inhibitor ML204 potentiated the photoswitching response (Fig. 3A, D, orange). Additionally, the TRPC6 inhibitor SAR7334 failed to block the photoswitching response (Fig. 3A and Fig. S4B) [56, 57].
TRPC channels mediate azo‐HU308's photoswitching effect on Ca2+ in INS‐1 cells
A recent study has shown TRPC channels' expression and regulation of insulin secretion from human and murine islets [58], and our broadband TRPC inhibitor SKF96365 reduced the amplitude of azo‐HU308 photoswitching effects in INS‐1 cells. Therefore, we focused on TRPC channels using blockers that have greater selectivity. Intriguingly, treatment with YM58483—which blocks TRPC3 and TRPC5 channels [59]—completely abolished the [Ca^2+^]i response caused by cis‐azo‐HU308 (Fig. 4A,B, red and Table S5). Next, we treated INS‐1 cells with the selective TRPC3‐specific inhibitor Pyr10 [60]. In this case, we saw a ~ 50% reduction in the Ca^2+^ response caused by photoswitching (Fig. 4A,B, blue and Table S5), demonstrating that TRPC3 may be involved but is not solely responsible for the effect. With both YM58483 and Pyr10 effectively reducing the azo‐HU308‐mediated Ca^2+^ response during irradiation, these results suggest that the proposed target of our photoswitch is likely through TRPC channels expressed in INS‐1 cells.
*TRPC channels are a target for azo‐HU308 in INS‐1 cells. INS‐1 cells were transfected with R‐GECO and Ca2+ levels recorded by confocal microscopy. Azo‐HU308 (20 μm) was applied in conjunction with TRPC channel antagonists. (A) Summary bar graph showing that TRPC channel inhibitors YM58483 (20 μm; N = 174, T = 4) and Pyr10 (15 μm; N = 200, T = 4) significantly reduced the photoswitching effect of azo‐HU308. (B) Averaged Ca2+ trace showing that treatment with the TRPC3 and TRPC5 inhibitor YM58483 (red) eliminated the Ca2+ photoswitching response, while the TRPC3‐selective antagonist Pyr10 (blue) reduced the Ca2+ photoswitching response by ~ 50%. Shaded error bars = mean ± SEM. *P < 0.05, **P < 0.01, **P < 0.001, ns, not significant. Student's t‐test.
Discussion
This study demonstrates that the photoswitchable cannabinoid azo‐HU308 robustly stimulates [Ca^2+^]i in INS‐1 β‐cells when activated by UV light and that the probe is more active in the cis‐configuration. While our first study using azo‐HU308 in CB2R‐overexpressing AtT20 cells showed that the probe triggers a prolonged Ca^2+^ response via Ca^2+^‐induced Ca^2+^‐release after CB2R and PLC activation [31], here an extensive pharmacological screen ruled out this mechanism of action. Notably, co‐application of three different CB2R antagonists—AM630, SR144528, and JWH130—failed to reduce azo‐HU308's effect, ruling out a CB2R‐mediated mechanism. In INS‐1 cells, the intense and short‐lived Ca^2+^ increase induced by cis‐azo‐HU308 is initiated solely by the influx of extracellular Ca^2+^, suggesting that an ion channel mechanism is involved. Our results also suggest that the plasma membrane ion channel target rapidly desensitizes, which led us to investigate the role of TRP ion channels. β‐cells express a multitude of TRP channels that permit the flux of Ca^2+^ ions into the cell, and we observed that the effect of azo‐HU308 was modulated by several TRP ion channel modulators, including classical nonselective TRP channel blockers such as 2‐APB and SKF96365. Gratifyingly, selective inhibitors for TRPC channels blocked azo‐HU308's effect on Ca^2+^, with the most significant inhibition coming from the TRPC3 and TRPC5 antagonist YM58483. These pharmacological results suggest that TRPC channels may play a role in mediating azo‐HU308's action on INS‐1 Ca^2+^ signaling.
A limitation of our study is that we have not ruled out that azo‐HU308 activates CB2R in INS‐1 cells, but only that CB2R is not responsible for the observed Ca^2+^ response. Studies have shown that CB2R are expressed in various β‐cell lines and islets [61], yet CB2R activation may cause signaling through other pathways, such as cAMP or MAP‐kinase signaling, that are not measured in this study. Future efforts to characterize the effect azo‐HU308 in INS‐1 cells on these different effector pathways may be warranted; however, our results suggest that the off‐target effects of azo‐HU308 prevent it from being a useful tool to study selective CB2R signaling in INS‐1 β‐cells. A second limitation to our study was that it is performed in an in vitro system, using only a monolayer of INS‐1 β‐cells. Future studies to investigate the action of azo‐HU308 on more physiologically relevant systems, such as human β‐cells or cultured islets that retain the 3D islet architecture and other endocrine cell types, may produce different results due to differential CBR expression and intercellular signaling between different islet cell populations. Additionally, integrating these findings with transcriptomic [62] or proteomic [63] analyses of CBR and TRP channel expression in different β‐cell systems may help clarify why significant discrepancies have been reported regarding cannabinoid actions in β‐cells.
In summary, we demonstrate that azo‐HU308 can optically stimulate Ca^2+^ levels in INS‐1 β‐cells with high spatiotemporal precision through a TRPC ion channel mechanism, revealing a non‐GPCR pathway activated by HU308 and its photoswitchable analog. While unexpected, cannabinoid ligands have been known to activate TRP channels in the TRPV, TRPM, and TRPA families [64, 65]. Our finding that (azo‐)HU308 can also activate TRPC channels underscores the importance of thoroughly characterizing the pharmacology of new chemical probes across diverse experimental systems, particularly at high concentrations, where ligand behavior can vary significantly. Overall, azo‐HU308 provides a new tool to investigate cannabinoid‐mediated activation of TRPC channels in β‐cells, and will advance our understanding of how natural and synthetic cannabinoid ligands influence β‐cell excitability and Ca^2+^ homeostasis.
Conflict of interest
The authors declare no conflict of interest.
Author contributions
JAF conceived of the project, provided funding, and oversight of the project. AEGV performed imaging experiments, data analysis, and figure preparation. AEGV and JAF contributed to the writing of the manuscript.
Supporting information
Fig. S1. azo‐HU308 increases Ca^2+^ transient in INS‐1 cells with stimulated by UV‐A light. Fig. S2. Nonphotoswitchable HU308 addition causes a short Ca^2+^ transient in INS‐1 cells. Fig. S3. azo‐HU308's photoswitching effect on Ca^2+^ is not CB1‐ or GPR55‐mediated. Fig. S4. azo‐HU308's photoswitching effect is not TRPV1‐ or TRPC6‐mediated. Table S1. Statistical significance calculations for Fig. 1E. Table S2. Statistical significance calculations for Fig. S1C. Table S3. Statistical significance calculations for Fig. 2A. Table S4. Statistical significance calculations for Fig. 3A. Table S5. Statistical significance calculations for Fig. 4A.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Klec C , Ziomek G , Pichler M , Malli R and Graier WF (2019) Calcium signaling in ß‐cell physiology and pathology: a revisit. Int J Mol Sci 20, 6110.31817135 10.3390/ijms 20246110 PMC 6940736 · doi ↗ · pubmed ↗
- 2Rutter GA and Hodson DJ (2015) Beta cell connectivity in pancreatic islets: a type 2 diabetes target? Cell Mol Life Sci 72, 453–467.25323131 10.1007/s 00018-014-1755-4PMC 11113448 · doi ↗ · pubmed ↗
- 3Nauck MA , Wefers J and Meier JJ (2021) Treatment of type 2 diabetes: challenges, hopes, and anticipated successes. Lancet Diabetes Endocrinol 9, 525–544.34181914 10.1016/S 2213-8587(21)00113-3 · doi ↗ · pubmed ↗
- 4Daly A and Hovorka R (2021) Technology in the management of type 2 diabetes: present status and future prospects. Diabetes Obes Metab 23, 1722–1732.33950566 10.1111/dom.14418 PMC 7611289 · doi ↗ · pubmed ↗
- 5Marcinak JF , Munsaka MS , Watkins PB , Ohira T and Smith N (2018) Liver safety of Fasiglifam (TAK‐875) in patients with type 2 diabetes: review of the global clinical trial experience. Drug Saf 41, 625–640.29492878 10.1007/s 40264-018-0642-6 · doi ↗ · pubmed ↗
- 6Hodson DJ , Tarasov AI , Gimeno Brias S , Mitchell RK , Johnston NR , Haghollahi S , Cane MC , Bugliani M , Marchetti P , Bosco D et al. (2014) Incretin‐modulated Beta cell energetics in intact islets of Langerhans. Mol Endocrinol 28, 860–871.24766140 10.1210/me.2014-1038 PMC 4042069 · doi ↗ · pubmed ↗
- 7Röthe J , Kraft R , Schöneberg T and Thor D (2020) Exploring G protein‐coupled receptor signaling in primary pancreatic islets. Biol Proced Online 22, 4.32082084 10.1186/s 12575-019-0116-y PMC 7023723 · doi ↗ · pubmed ↗
- 8Veilleux A , Di Marzo V and Silvestri C (2019) The expanded endocannabinoid system/Endocannabinoidome as a potential target for treating diabetes mellitus. Curr Diab Rep 19, 117.31686231 10.1007/s 11892-019-1248-9 · doi ↗ · pubmed ↗