Characterization of the clade 4 non-toxigenic C. difficile isolate L-NTCD03 carrying the cfr(B) gene
Britt Nibbering, Sam Nooij, Céline Harmanus, Ingrid M J G Sanders, Inez M Miedema, Quinten R Ducarmon, Rolf H A M Vossen, Susan L Kloet, Colleen K Ardis, Robert A Britton, Farnaz Yousefi, Jenna Bayne, Chandrashekhar Charavaryamath, Andy Law, Morgan L Murphy, Brett Sponseller

TL;DR
This study characterizes a non-toxigenic strain of Clostridioides difficile, L-NTCD03, which could be useful for developing new therapies.
Contribution
The study provides a detailed genomic and functional analysis of a non-toxigenic C. difficile strain carrying the cfr(B) gene.
Findings
L-NTCD03 is a non-toxigenic C. difficile strain resistant to multiple antimicrobials.
The cfr(B) gene in L-NTCD03 confers resistance to clindamycin, linezolid, retapamulin, and streptogramin A.
L-NTCD03 is a potential candidate for developing live biotherapeutic products.
Abstract
Clostridioides difficile infection (CDI) is a toxin-mediated gastro-intestinal disease. Yet, C. difficile is a phylogenetically diverse species that includes many non-toxigenic strains. In general, these are understudied, despite having significant potential impact for our understanding of the colonization process and as therapeutic modalities. Here, we present an in-depth characterization—including the complete genome sequence—of the non-toxigenic C. difficile strain L-NTCD03. This strain belongs to PCR ribotype 416, clade 4 and multilocus sequence type 39. It is resistant to multiple antimicrobials, but not those used for treatment of CDI. We validated the relevance of the cfr(B) gene from this strain in antimicrobial resistance to clindamycin, linezolid, retapamulin, and streptogramin A. We found the L-NTCD03 strain to be non-toxic in various assays. Altogether, L-NTCD03 is a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
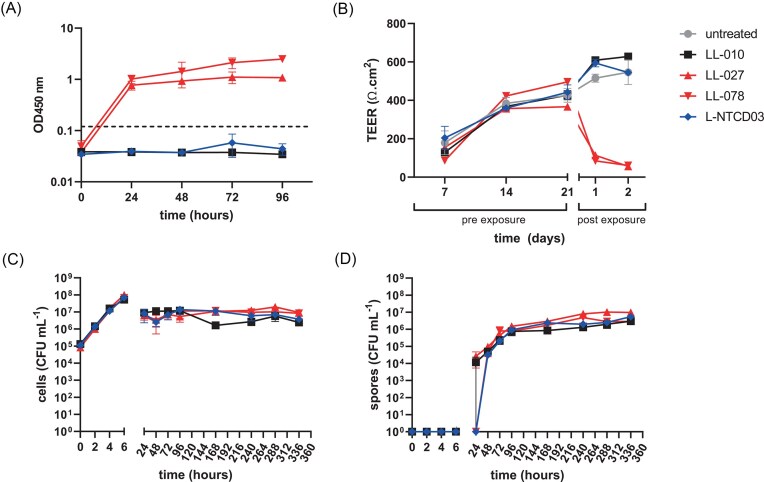 Figure 1
Figure 1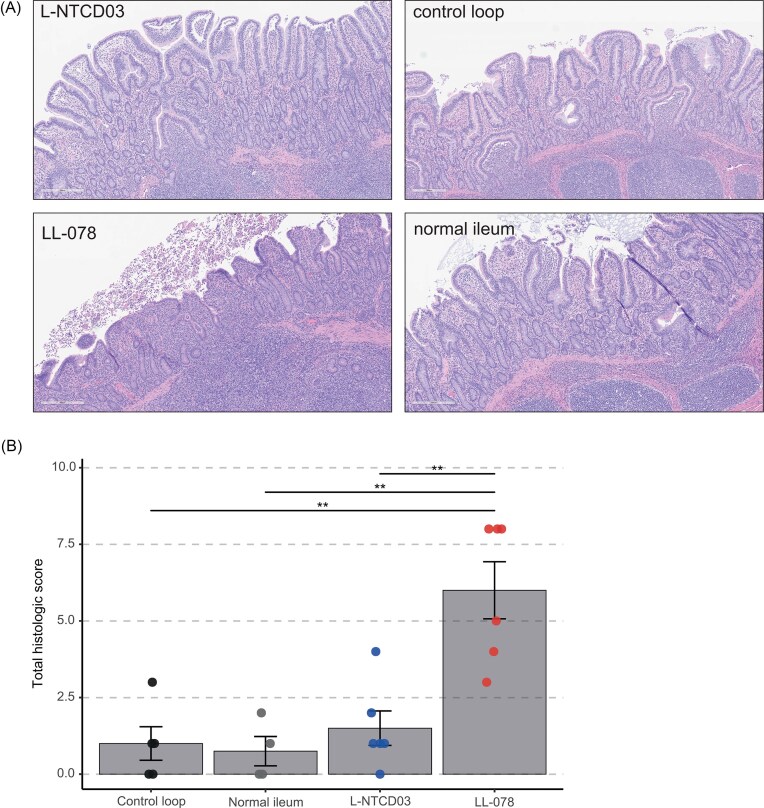 Figure 2
Figure 2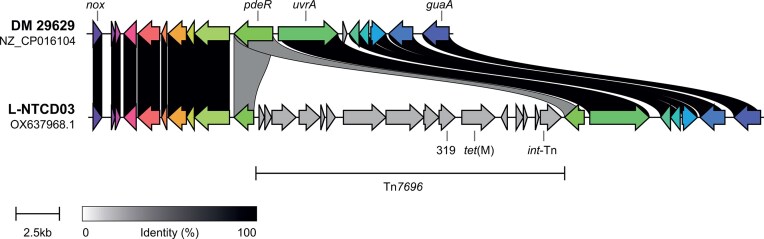 Figure 3
Figure 3| Oligonucleotide | Sequence (5’>3’) | Reference |
|---|---|---|
| 16S-F | TET | (Fawley et al. |
| 23S-R | CCCTGCACCCTTAATAACTTGACC | (Fawley et al. |
| NF793 | CACCTCCTTTTTGACTTTAAGCCTACGAATACC | (Fagan and Fairweather |
| NF794 | CACCGACGAGCAAGGCAAGACCG | (Fagan and Fairweather |
| oIMM-001 | CTACGGTACCCTCTACCTAATAGCCACAGGAG | This study |
| oIMM-002 | GATGGGATCCTCACTGCATCCGCAATTC | This study |
| oIMM-003 | GTTAATTGAGGAGTTTGGAGAATCG | This study |
| oIMM-004 | CACATCTGATAATGGGTAGCGTTC | This study |
| oWKS-1070 | GTCTTGGATGGTTGATGAGTAC | (Boekhoud et al. |
| oWKS-1071 | TTCCTAATTTAGCAGCAGCTTC | (Boekhoud et al. |
| oWKS-1387 | CAGATGAGGGCAAGCGGATG | (Boekhoud et al. |
| oWKS-1388 | CGTCGGTGAGCCAGAGTTTC | (Boekhoud et al. |
| Antibiotic | MIC (mg/l) | EUCAST (T)ECOFF (mg/l) | Putatively associated AMR gene |
|---|---|---|---|
| Amoxicillin/clavulanic acid | 0.25 | – | – |
| Cefuroxime | >256 | – |
|
| Chloramphenicol | >256 | – |
|
| Ciprofloxacin | >32 | (32) |
|
| Clindamycin | >256 | 16 |
|
| Cotrimoxazole | 0.06 | – | – |
| Erythromycin | 48 | 4 |
|
| Fidaxomicin | 0.06 | 0.5 | – |
| Linezolid | 6 | – |
|
| Meropenem | 1 | (8) |
|
| Metronidazole | 0.06 | 2 | – |
| Moxifloxacin | 1 | 4 |
|
| Penicillin | 0.75 | – |
|
| Rifampicin | >32 | 0.004 |
|
| Tetracycline | 2 | (0.25) |
|
| Teicoplanin | 1 | – |
|
| Vancomycin | 0.38 | 2 |
|
| Plasmid | Antimicrobial MIC (mg/l) | |||
|---|---|---|---|---|
| Clindamycin | Linezolid | Retapamulin | Streptogramin A | |
| pAP24 (control) | 2 | 1 | 0.125 | 2 |
| pIMM03 ( | >256 | 12 | 16 | 16 |
- —Federation of European Microbiological Societies10.13039/501100000614
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsClostridium difficile and Clostridium perfringens research · Antimicrobial Resistance in Staphylococcus · Bacillus and Francisella bacterial research
Introduction
The Gram-positive anaerobic spore-forming bacterium Clostridioides difficile is best known as a nosocomial pathogen and C. difficile infection (CDI) is regarded as an important cause of healthcare-associated infectious diarrhea (Leffler and Lamont 2015, Smits et al. 2016). In this context, in particular strains belonging to the fluoroquinolone resistant PCR ribotype 027 (RT027) are associated with increased morbidity and mortality (Warny et al. 2005, He et al. 2013). Nevertheless, recent years have seen an increase in CDI cases that are not linked to classical risk factors such as antibiotic use or healthcare exposure, and are community-acquired (Smits et al. 2016, Viprey et al. 2022). Strains belonging to different types than RT027, such as those belonging PCR ribotype 078 (RT078), which also occur in many animal species, may be associated with community-acquired disease (Goorhuis et al. 2008, Viprey et al. 2022). Thus, C. difficile can be considered a quintessential One Health pathogen (Lim et al. 2020).
Considering its causative role in debilitating disease, much research has focused on the main virulence factors of C. difficile: toxin A (TcdA), toxin B (TcdB) and binary toxin (CDT) (Aktories et al. 2017, Kordus et al. 2022). Of note, whereas TcdA and/or TcdB is required for disease, CDT is mainly produced by a subset of toxigenic C. difficile strains, including those belonging to the epidemic types RT027 and RT078 (Smits et al. 2016).
Clostridioides difficile colonization does not always lead to disease. Importantly, a fraction of the population is asymptomatically colonized by toxigenic C. difficile (Crobach et al. 2018) and in particular in infants a high prevalence of C. difficile is noted without apparent disease (Ferretti et al. 2023, Mani et al. 2023). Additionally, C. difficile is not a universally pathogenic species; that is, non-toxigenic C. difficile (NTCD) strains exist that lack the genes encoding TcdA, TcdB, and CDT, such as those belonging to PCR ribotype 010 (RT010) (Braun et al. 1996, Monot et al. 2015, Smits et al. 2016). Though these isolates have not been found to induce disease in humans and animals, it has been noted that they might act as reservoirs for antimicrobial resistance genes. For instance, resistance to metronidazole is common in RT010 strains (Boekhoud et al. 2020, Boekhoud et al. 2021) and over half the non-toxigenic isolates from Southeast Asia appeared to carry antimicrobial resistance determinants on mobile genetic elements (Imwattana et al. 2020).
C. difficile is a phylogenetically diverse species with isolates falling into at least five classical clades and three so called cryptic clades, that could be considered a separate genomospecies (Knight et al. 2021). Epidemic strains fall generally into clade 2 (RT027 and related strains) and clade 5 (RT078 and related strains). NTCD have been reported in multiple clades, such as clade 1 (which includes RT010) (Baktash et al. 2022), clade 4 (Imwattana et al. 2020), clade C-I, and clade C-II (Ducarmon et al. 2023). Nevertheless, such strains are generally understudied compared to the toxigenic counterparts.
There is epidemiological and experimental evidence that NTCD carriage is associated with lower incidence of CDI, that NTCD colonization can prevent CDI disease and can reduce CDI recurrence in intestinal models, animals and humans (Natarajan et al. 2013, Gerding et al. 2018, Etifa et al. 2023). The strongest evidence for use of NTCD as a therapeutic intervention is for strain M3 (a RT010 isolate) in humans (Villano et al. 2012, Gerding et al. 2015, Sambol et al. 2023) and strain Z31 (RT009) in piglets (Oliveira Junior et al. 2019a,b).
In addition to potential use as a live biotherapeutic product, NTCD could provide a chassis for engineering of immune-epitopes and therefore serve as a potential vaccine (Wang et al. 2022a,b, Hughes et al. 2022), or be used to study aspects of CDI disease that are generally confounded by toxin-mediated effects, such as early colonization events.
In light of the possible applications of NTCD, and the relative paucity of well-characterized NTCD strains, we present here a detailed analysis of L-NTCD03, a clade 4 non-toxigenic C. difficile strain.
Materials and methods
Bacterial strains and growth conditions
L-NTCD03 was isolated from fecal material of a healthy elderly subject negative for HIV-1, HIV-2, HCV, HBV, as follows. Fecal material was stored in Stool Transport and Recovery (STAR) buffer and stored at −80°C. Ethanol treated material was inoculated into C. difficile enrichment modified broth (CDEB MOB; Mediaproducts NV, Groningen, The Netherlands, cat no. 21.1295). After 6 days of growth at 37°C under anaerobic conditions, the culture was plated onto C. difficile selective growth medium (CLO; VWR International, cat.no. 43 431). From this medium, a colony was streaked to singles on a Tryptic Soy Sheep Blood medium (TSS; bioMérieux, cat.no. 43 009), and growth was harvested into glycerol-containing broth (Tritium Microbiology, cat. no. I200.36.0001) for storage at −80°C.
Control strains belonging to PCR ribotype 027 (LL-027) and 078 (LL-078) were obtained from the Leeds–Leiden collection (Knetsch et al. 2012, Baktash et al. 2022).
Strains were cultured in simulated ileal effluent medium (SIEM) (Minekus et al. 1999, Wiese et al. 2022), Brain–Heart Infusion (BHI) broth, BHI supplemented with yeast extract (BHI-Y; BHI with 0.5% yeast extract) or C. difficile base medium supplemented with moxalactam and norfloxacin [CDMN; (Aspinall and Hutchinson 1992)] supplemented with 0.1% sodium taurocholate (CDMNT).
Molecular biology
A genomic region including cfr(B) and its native promoter was amplified by PCR using Q5 polymerase (NEB) and primers oIMM-01 and oIMM-02. The resulting PCR product was digested with KpnI and BamHI and cloned into similarly digested pAP24 (Oliveira Paiva et al. 2016). The resulting plasmid, pIMM03, was sequence verified using primers NF793, NF794 (Fagan and Fairweather 2011), oIMM-01, oIMM02, oIMM-03, and oIMM-04 and was introduced using standard laboratory methods into strain 630Δerm (van Eijk et al. 2015, Roseboom et al. 2023). Transconjugants were verified by PCR using primers oWKS-1070/oWKS-1071 (targeting chromosomal gluD), oWKS-1387 and oWKS-1388 (targeting plasmid-located traJ), and oIMM-01/oIMM-02 [targeting cfr(B)] using appropriate controls. All oligonucleotide sequences are listed in Table 1.
Antimicrobial susceptibility testing
To determine minimal inhibitory concentrations (MIC) of a panel of antibiotics gradient tests were used. This panel consisted of cotrimoxazol, linezolid, metronidazole (Liofilchem), amoxicillin/clavulanic acid, cefuroxime, ciprofloxacin, chloramphenicol, clindamycin, erythromycin, meropenem, moxifloxacin, penicillin, rifampicin, tetracyclin, teicoplanin, and vancomycin (bioMérieux). For this test, bacterial suspensions corresponding to a turbidity of 1.0 McFarland unit were plated on Brucella blood agar supplemented with 5 mg l^−1^ hemin and 1 mg l^−1^ vitamin K (bioMérieux) plates, unless indicated otherwise. MIC values were determined after 48 h of anaerobic incubation at 37°C as recommended by Clinical and Laboratory Standards Institute (CLSI), including relevant control strains (CLSI 2012). Fidaxomicin MIC was determined using the agar dilution method according to CLSI guidelines on fresh Brucella Blood Agar supplemented with 5 mg l^−1^ hemin and 1 mg l^−1^ vitamin K. Epidemiological cut-off values were used as defined by European Committee on Antimicrobial Susceptibility Testing (EUCAST 2025), when available.
MIC measurements for engineered strains (three independent transconjugants) were performed on BHI-yeast extract agar with either an E-test (clindamycin, linezolid) (bioMérieux or Liofilchem, respectively) or by incorporating retapamulin (Sigma CDS023386) or streptogramin A/virginiamycin M1 (Sigma V2753) at 0–128 mg/l into the agar.
PCR ribotyping
Capillary gel electrophoresis ribotyping of L-NTCD03 was performed according to standardized protocols (Fawley et al. 2015). In short, 2 µl of isolated DNA was added to 23 µl of the master mix which includes the 16S and 23S primers and HotStart Taq Master Mix (Qiagen). Using a MyCycler thermocycler (Biorad), the resulting DNA mix was subjected to an enzyme activation step at 95°C for 15 min followed by 24 cycles of amplification at 95°C for 1 min, 1 min at 57°C, and 1 min at 72°C. Finally, the DNA mix was subjected to 72°C for 30 min. The sample was prepared for analysis by adding PCR product as 1 µl of PCR product to 8.5 µl HiDi formamide (Applied Biosystems) and 0.5 µl of GeneScan™ 1200 LIZ marker (Thermo Fisher) and samples was denatured at 95°C for 5 min. To determine the ribotype, it was then analyzed using the ABI3500XL (capillary length 50 cm) and the resulting *.fsa files were compared to a reference database in BioNumerics (bioMérieux).
Toxin ELISA
To determine the productions of Toxins A and B, the C. difficile Tox A/B II^TM^ kit (Techlab) was used according to manufacturer’s protocol with a minor modification. In short, the supernatants that were collected at 0, 24, 48, 72, and 96 h from growth curves in SIEM pH=5.8 and pH=6.8 were filtered using a UNIFILTER microplate device (0.45 µm cellulose acetate filter, long drip, 96 wells, GE Health Care Life Sciences). The samples were applied to the filter plate and centrifuged at 3220 G for 5 min at 4°C. The filtrates were used in the analysis as described by manufacturer and the optical density was measured at 450 nm (OD_450_). A threshold value of OD_450_ of 0.12 was used determine whether toxins were present in the filtrate.
Cell lines and culture conditions
Human colorectal adenocarcinoma cells (Caco-2) were obtained from the American Type Culture Collection (ATCC-HTB-37) and cultured in minimum essential medium (MEM, Gibco) supplemented with 10% bovine fetal serum (10% FCS), 1% non-essential amino acids (NEAA, Lonza), 2 mM glutamine (Sigma Aldrich), 100 units/ml penicillin (Sigma Aldrich), and 100 units/ml streptomycin (Sigma Aldrich). Epithelial-like, mucin-producing, LS174T cells were obtained from Sigma Aldrich (87 060 401) and cultured in the medium described for Caco-2. Both cells were grown to 80% confluency before experimental conditions were prepared.
In vitro intestinal epithelial barrier model
The apical side of Thincert 12 well cell culture inserts (PET, pore size 0.4 µm, Greiner Bio-one) were coated with 250 µg/well purified human collagen type IV (OptiCol collagen Type IV, dissolved in 0.25% acetic acid, Cell Guidance Systems) and washed with sterile PBS after 1–2 h incubation.
To mimic the human colon environment, an 80:20 ratio mixture of Caco-2 and LS174T in MEM supplemented with 10% FCS, 1% non- NEAA, 2 mM glutamine, 100 U/ml penicillin, and 100 U/ml streptomycin (cell culture medium) was prepared. To function as controls, 100:0 and 0:100 ratio mixtures of Caco-2 and LS174T were prepared in cell culture medium. For all conditions, 1*10^5^ cells were seeded on the apical side of the PET membrane. To mimic the intestinal environment, medium was present in both the basal compartment (the well) and apical compartment (on top of the PET membrane) of the system. The systems were incubated at 37°C and 5% CO_2_. The cells in this model were allowed to grow and differentiate for 21 days during which the medium in both compartments was refreshed every other day.
To monitor the development of the in vitro intestinal epithelial barrier model, starting at T=0 the transepithelial electrical resistance (TEER) values were measured using EVOM-2 resistance meter (World Precision Instruments) every 7 days.
On day 21, the medium in the apical compartment of the system was replaced with a 10 times diluted, in cell culture medium, supernatant harvested at 72 h in a growth curve of all of the strains in BHI-y medium. This supernatant was incubated in the apical compartment for 1 h at 37°C and 5% CO_2_, removed afterwards and replaced with cell culture medium. In every condition, a control well was exposed to calcium free medium (SMEM, Gibco) to determine as a positive control. TEER measurements were performed at 4 (only SMEM condition), 24, and 48 h post-exposure. Morphology was assessed under light microscopy every 24 h.
Swine IsoLoop model
We compared the effect of L-NTCD03 and LL-078 on intestinal mucosa in vivo in the pig IsoLoop model, a translational surgical in situ ileal loop platform for host-microbiome-pathogen interface (Bayne et al. 2025). Twelve hours before the start of the surgery, 1 ml was drawn from a culture in the exponential growth phase of either L-NTCD-03 or LL-078 in BHI-y and inoculated into fresh, pre-reduced CDMNT medium and grown anaerobically at 37°C. Ten milliliter of culture was aliquoted per loop and stored anaerobically at room temperature until injection was required.
Three-month-old, female pigs (Sus scrofa, mixed breed, 50–60 kg) were used under USDA pain category D, per Institutional Animal Care and Use Committee (IACUC) protocol IACUC-22–066 essentially as described (Bayne et al. 2025).
In short, the pig was placed in dorsal recumbency under general anesthesia. Ileal loops were rinsed with sterile saline solution before treatment with an antibiotic cocktail (60 mg/l norfloxacin; 160 mg/l moxalactam) for 30 min. After a saline rinse, each independent loop except for the most aboral segment was inoculated using a tuberculin needle full thickness into the lumen. Upon recovery, animals were housed under standard husbandry conditions with regular veterinary assessments to ensure welfare. The loops were sampled at euthanasia by cutting the mesentery and immediately cooling the intestines on ice. On ice, the loops were cut open one by one and a piece of tissue, including intestinal content, was fixed in 10% neutral buffered formalin (VWR) before embedding in paraffin.
Five micrometre thick tissue sections were stained with hematoxylin and eosin. Microscopic analysis of histopathologic features was conducted by a board-certified pathologist, blinded to sample identities. The scoring rubric, including descriptive statistics applied, is provided as Supplementary Information, and raw data of the scoring is included as Supplementary Table 1. Scores are reported as total histological score per ileal loop (range 0–20), computed as the sum for four 0–5 ordinal components (epithelial necrosis, mucosal/lamina propria edema, crypt abscess, and neutrophils in villi/lamina propria). Statistical analyses of histological scores were performed in Python (NumPy/SciPy); figures were generated with matplotlib. All tests were two sided with α = 0.05.
DNA extraction and genome sequencing
To extract pure high-molecular weight total DNA, a two-fold serial dilution of L-NTCD03 in BHI-Y medium was prepared. After overnight incubation at 37°C in an anaerobic environment, 4 ml of culture with an optical density at 600 nM between 0.6 and 0.8 was spun down and DNA was extracted from the cell pellet using the 500/G genomic tip kit (Qiagen), according to the instructions of the manufacturer. Quantity and quality of the DNA was verified by gel electrophoresis on a 0.8% agarose gel, using the Qubit dsDNA high-sensitivity kit (Qubit) and a Femto Pulse system (Agilent), according to the manufacturer’s protocols.
PacBio sequencing libraries were generated according to the manufacturer’s multiplexed microbial library preparation protocol, part number 101–696-100, version 7, July 2020 release (Pacific Biosciences) using the SMRTbell Express Template Prep Kit v2.0 with the following modifications: genomic DNA was sheared using Speed 34 on the Megaruptor 3 (Diagenode) and an additional size selection step of 6–50 kb fragments on the Blue Pippin (Sage Science) was included for the final SMRT bell library. The libraries were sequenced on a Sequel II platform (Pacific Biosciences) using the Sequel II Binding Kit v2.0, Sequencing Primer v4, Sequencing Kit v2.0, and a 30hr movie time.
Genome annotation and bioinformatics analyses
De novo genome assembly was carried out using Hifiasm (version 0.16.1-r375; parameters “–hg-size 4m”) (Cheng et al. 2021) and the start position was set to dnaA using Circlator fixstart (version 1.5.5 using default parameters) (Hunt et al. 2015). Completeness check was performed using Busco (version 5.2.2; parameters “-m genome -l clostridia_odb10”) (Manni et al. 2021) and assembly statistics were calculated and visualized with Quast (version 5.0.2; using NZ_CP076401 as reference genome) (Gurevich et al. 2013). Finally, Prokka (version 1.14.6; parameters “–genus Clostridioides –species difficile –compliant”) (Seemann 2014) was used to annotate the genome.
A database of AMR genes was constructed using the AMRFinder plus tool v3.10.23 (parameters “-n” “–organism C. difficile”, “—plus”) to identify antimicrobial resistance (AMR) genes (Feldgarden et al. 2021). Minimal “% coverage of reference sequence” and “% Identity to refence sequence” were set to 60% and 90% %, respectively.
BacAnt (Hua et al. 2021) and MobileElementFinder (Johansson et al. 2021) were used to identify putative transposable elements. progressiveMauve was used to align genome sequences (Darling et al. 2010).
Multi-locus sequence typing (MLST) was performed using mlst (v2.19.0) with the PubMLST C. difficile database updated to October 21, 2021, (Jolley et al. 2018) and verified on December 1, 2025.
Genomic comparisons were made in Geneious R10 (BioMatters Ltd), and figures for publication were prepared using CAGECAT (van den Belt et al. 2023) and Adobe Illustrator 26.3.1.
Results
L-NTCD03 belongs to a rare PCR ribotype, RT416
As part of a study investigating carriage of multidrug resistant organisms and C. difficile in nursing homes (Ducarmon et al. 2021), fecal material was obtained from a healthy elderly subject. C. difficile was cultured from this material as described in materials and methods. The resulting isolate was negative in a multiplex PCR that targets the toxin genes tcdA, tcdB, cdtA, and cdtB (ECDC 2018), suggesting that the isolate was non-toxigenic. Hereafter, the isolate is referred to as L-NTCD03.
We performed capillary electrophoresis PCR ribotyping (Fawley et al. 2015) of L-NTCD03 through the Dutch Expertise Center for C. difficile, hosted at the LUMC. We initially failed to obtain a ribotype assignment, as the banding pattern was not present in our database. The *.fsa files were shared with the C. difficile Ribotyping Network Laboratory (CDRN; PHE Microbiology Services, Leeds, UK), and they determined the isolate to belong to PCR ribotype 416. Since this identification, no other isolates were identified with the same ribotype in the Dutch National Expertise Center for C. difficile infections. This suggests a prevalence in the Netherlands that is below 0.01% on the basis of the number of strains typed since then.
L-NTCD03 is susceptible to clinically used antimicrobials
To determine whether L-NTCD03 the minimal inhibitory concentration for different antimicrobials, we performed antimicrobial susceptibility testing using agar dilution (for fidaxomicin) or gradient test (amoxicillin/clavulanic acid, cefuroxime, chloramphenicol, clindamycin, cotrimoxazol, erythromycin, linezolid, metronidazole, meropenem, moxifloxacin, penicillin, rifampicin, tetracyclin, teicoplanin, and vancomycin) methods on Brucella Blood Agar, as recommended by CLSI (CLSI 2012). Out of the 17 antimicrobials tested, L-NTCD03 demonstrated elevated MICs for cefuroxime, chloramphenicol, ciprofloxacin, erythromycin, linezolid, rifampicin, and tetracycline (Table 2). In contrast, the strain is susceptible to antimicrobials that are used in CDI therapy—fidaxomicin, vancomycin, and metronidazole—as well as several others (Table 2). As Brucella Blood Agar is a suboptimal medium for determining resistance to metronidazole, we also retested the isolate on recently recommended Fastidious Anaerobe Agar supplemented with horse blood (Freeman et al. 2025); this confirmed susceptibility to metronidazole (data not shown). These results demonstrate that L-NTCD03 would likely be suppressed or eradicated using standard antimicrobial therapy for CDI, but not by certain other antimicrobials that increase susceptibility for CDI.
Supernatant from L-NTCD03 does not contain detectable toxins A and B
To confirm the absence of toxins, as suggested by the multiplex PCR result, we assessed the presence of TcdA and TcdB in culture supernatant using an enzyme-coupled immune assay (Techlab). We used a medium that mimicks the physiological conditions in the gut: simulated ileal effluent medium, SIEM (Minekus et al. 1999, Wiese et al. 2022).
During 96 h of growth in SIEM, toxin levels were below the limit of detection for all supernatants derived from L-NTCD03, as well as for the reference non-toxigenic strain LL-010 (Fig. 1A). In contrast, toxins were readily detected in culture supernatants derived from the toxigenic reference strains LL-027 and LL-078 (Fig. 1A).
L-NTCD03 is a non-toxigenic C. difficile strain. A. L-NTCD03 does not show production of toxins in an ELISA assay. The dotted line indicates the threshold value indicated by the manufacturer for defining toxin production. B. Culture supernatant from L-NTCD03 does not lead to a reduction in transepithelial resistance as measured in a Caco2/LS174T cellular model that was matured for 21 days (pre-exposure) before applying culture supernatant (post exposure). C. Growth of L-NTCD03 is similar to other C. difficile strains. Colony forming units (CFU) per milliliter culture were determined by a droplet-plating method. D. Spore formation of L-NTCD03 is similar to other C. difficile strains. Spores were enumerated after ethanol treatment using a droplet plating method. Values indicated are the mean and standard error of the mean of n=3 biological replicates. LL-010 is a non-toxigenic control strain and LL-027 and LL-078 are toxigenic controls.
Supernatant from L-NTCD03 does not affect transepithelial electrical resistance
Though our results clearly indicate the absence of TcdA and TcdB, we considered that there could be other factors that affect epithelial integrity, not detected in our assays so far. For this reason, we assessed the effect of culture supernatant derived from L-NTCD03, LL-010, LL-027, and LL-078 in an in vitro model for epithelial integrity. In this model, a mixture of Caco2 and mucin-producing LS174T cells is grown in a transwell system and the transepithelial electrical resistance (TEER), a proxy for epithelial integrity, is measured using electrodes.
The TEER values gradually increased over the first 14 days of incubation, consistent with cells growing to confluency, and stabilized between days 14 and 21 (Fig. 1B). When filtered supernatants from 72 h-cultures in BHI were applied to the cell layer, no drop in TEER was observed for the non-toxigenic strains (LL-10 and L-NTCD03), whereas LL-027 and LL-078 derived supernatants resulted in a significant decrease (Fig. 1B). Similar results were obtained when only LS174T cells or Caco2 cells were used (data not shown). This shows that no other factors are produced by the non-toxigenic C. difficile strains that affect TEER in this cellular model.
L-NTCD03 grows similar to other isolates in a simulated ileal effluent medium
To assess the growth and sporulation of L-NTCD03 in relation to well-characterized common subtypes of C. difficile in SIEM, we monitored the total number of colony forming units (CFU) per milliliter (ml) over a period of 14 days. Spore formation was monitored by enumeration of ethanol resistant CFUs.
Growth of L-NTCD03 was indistinguishable from LL-010, LL-027, and LL-078 for the first 6 h, during which the number of CFU/ml increased ∼3 log (Fig. 1C). During stationary phase, we observed a lower total cell count of ∼10^7^ CFU/ml, for all strains (Fig. 1C).
No spore production was found during the first 6 h of growth in SIEM (Fig. 1D). For LL-010 and LL-027 spores were detectable at 24 h, but for L-NTCD03 and LL-078 spores were first found at the 48 h-timepoint. Beyond this point, sporulation dynamics were comparable between all strains Fig. 1D).
Thus, under the conditions tested, L-NTCD03 shows robust growth and sporulation, in line with other C. difficile strains.
Phenotype microarrays show a common carbon utilization profile for L-NTCD03
In order to assess the carbon utilization by L-NTCD03 we performed phenotype microarrays on Biolog PM1 and PM2 plates, and analyzed the results using AMIGA software, as described previously (Midani et al. 2021). We used an arbitrary cut-off of 1.5-fold increase in or a two-fold decrease in optical density to define differential growth. We observed stimulated growth on the carbon sources N-Acetyl-D-Glucosamine, D-Mannose, D-Mannitol, D-Fructose, alpha-D-Glucose, N-AcetylNeuraminic Acid, D-Melezitose, D-Tagatose, and D-Glucosamine (Supplemental Table S1). A decreased cell density was observed in the presence of Glyoxylic Acid and Glycyl-LGlutamic Acid (Supplemental Table S1).
In parallel experiments, we observed similar profiles for representative strains from PCR ribotype 010, 027, and 078 (data not shown) (Collins et al. 2018, Midani et al. 2025). We conclude that L-NTCD03 carbon utilization is similar to that of other C. difficile isolates.
L-NTCD03 does not induce C. difficile-associated intestinal lesions in the swine IsoLoop model
We have previously demonstrated the utility of the swine IsoLoop model for assessing C. difficile–associated lesions in a translationally relevant gut microbiota–pathogen interface (Bayne et al. 2025). Using this platform, we compared the effects of L-NTCD03 in relation to the effects induced by the toxigenic reference strain LL-078.
We inoculated multiple loops in the same animal (one animal per strain, either LL-078 or L-NTCD03), and compared the histology to control loops without C. difficile and normal ileum (Fig. 2). Loops inoculated with LL-078 showed signs of inflammation, whereas those inoculated with L-NTCD03 were comparable to the control loop (Fig. 2A). The total histological score (further detailed in the Methods section of this manuscript) of samples derived from loops inoculated with LL-078 were significantly higher than those derived from, whereas the histology of L-NTCD03 derived samples was comparable to that of control loops (Fig. 2B and Supplemental Table 1).
L-NTCD03 does not induce pathology in the swine IsoLoop model. A. Representative images from histological analyses. Tissues samples were formalin fixed and paraffin embedded, sectioned and hematoxylin and eosin stained. No significant C. difficile-associated lesions are observed in the control loop (an ileal loop that was antibiotic-treated but not inoculated with C. difficile), normal ileum (not a loop but control for surgical integrity of the loops) and the loop inoculated with L-NTCD03. In contrast, the loop inoculated with LL-078 (toxigenic strain, positive control for C. difficile pathology) showed blunting of the villi with epithelial necrosis, neutrophilic infiltration of lamina propria, and crypt abscesses. B. Combined histological scores for the different samples. A bar plot depicting the mean histological score with standard error of the mean is given, with individual datapoints depicted. Prespecified contrasts (see Supplementary Information) are shown with brackets, with asterisks indicating the raw P-value magnitude, for readability (**: P < 0.01), while significance is determined by Holm adjustment across the three prespecified tests.
L-NTCD03 is an ST39 (clade 4) isolate carrying a cfr(B) resistance gene
Clostridioides difficile is a phylogenetically diverse species, with isolates falling into at least five classical (CDI-associated) clades and three so called cryptic clades, that have been proposed to form separate genomospecies (Knight et al. 2021). To place the L-NTCD03 in phylogenetic context, we reconstructed the complete genome sequence of the strain using Pacific Biosciences circular consensus sequencing (BioProject PRJEB62683). After assembly, the resulting genome showed a single circular contig of 4 318 385 bp—indicating the absence of extrachromosomal elements—and an average [G+C]-content of 28,91%. Reference assembly of short-read (Illumina) sequence data did not identify any nucleotide variants (data not shown), underscoring the quality of the assembly.
We derived the seven-gene multilocus sequence type of our isolate using PubMLST (Griffiths et al. 2010, Jolley et al. 2018). L-NTCD03 was found to belong to ST39 (adk 3, atpA 7, dxr 10, glyA 8, recA 7, sodA 2, tpi 10), which falls into clade 4. A core genome phylogenetic tree of representative isolates of different C. difficile clades using Panaroo and IQTree confirmed this placement (data not shown) (Minh et al. 2020, Tonkin-Hill et al. 2020, Baktash et al. 2022).
We performed automated annotation of the genome sequence using Prokka (Seemann 2014). This resulted in 3844 predicted coding sequences, 88 tRNAs, 34 rRNAs, 1 tmRNA and eight repeat regions. As expected, no complete genes corresponding to the toxin genes tcdA, tcdB, cdtA, and cdtB were detected; in stead the PaLoc insertion site conforms to what has been described for clade 4 strains *(*115 bp insertion) (Dingle et al. 2014).
We used AMRfinderPlus to identify antimicrobial resistance genes in the L-NTCD03 genome (Feldgarden et al. 2022). We found 14 putative antimicrobial resistance genes, which are associated with 12 different classes of antibiotics (Supplemental Table 2); ten of these might be linked to the phenotypically observed reduced susceptibility/resistance (Table 2).
Though cfr(B) has been implicated in antimicrobial resistance of C. difficile to PhLOPSa (phenicol, lincosamide, oxaxolidinone, pleuromutilin, and streptogramin A) antimicrobials (Marin et al. 2015, Stojkovic et al. 2019), a direct causal role for this gene in resistance to these antimicrobials has not been shown to date. Therefore, we cloned the cfr(B) gene under its native promoter in a shuttle plasmid and introduced this plasmid into the commonly used C. difficile laboratory strain 630Δerm (Hussain 2005, van Eijk et al. 2015). We assessed antimicrobial susceptibility using gradient tests and found that introduction of cfr(B) increased the minimal inhibitory concentration for clindamycin, linezolid, retapamulin, and streptogramin A (Table 3) in comparison to a control strain carrying a similar plasmid that carried the reporter gene sluc^opt^ (Oliveira Paiva et al. 2016) rather than cfr(B). The MIC increased >128-fold for clindamycin (from 2 mg/l to >256 mg/l), 12-fold for linezolid (from 1 mg/l to 12 mg/l), 64-fold for retapamulin (from 0.125 mg/l to 16 mg/l), and 8-fold for streptogramin A (from 2 mg/l to 16 mg/l) in the presence of cfr(B). We observed no change in MIC for vancomycin and metronidazole (data not shown). This result indicates that cfr(B) can indeed confer resistance to lincosamide, oxazolidinone, pleuromutilin, and streptogramin antimicrobials in C. difficile, consistent with expectations (Marin et al. 2015, Stojkovic et al. 2019).
The genome of L-NTCD03 harbors several putative mobile genetic elements
The BacAnt annotation server (Hua et al. 2021) identified two putative transposable elements, with homology to Tn4451 (GenBank U15027; 94.57% identity and 100% coverage; bp 524 023–530 368) and Tn5405 (GenBank U73027; 99.84% identity and 95.97% coverage; bp 551 080–553 654). Of note, the catP gene is contained on the former, suggesting a potential for transmission of this resistance determinant. MobileElementFinder (Johansson et al. 2021), in addition to Tn4551, identifies a chromosomal region with homology to Tn6009 (GenBank EU399632; identity 99.95% and coverage 100%; bp 268 640–270 528). Comparison with a closely related clade 4 genome (DSM 29629, GenBank NZ_CP016104) reveals that this is in fact a 17.9-kb transposon highly similar to Tn916, which has inserted in a gene (pdeR) encoding a GGDEF domain containing protein (in L-NTCD03 this gene is split into two ORFs: LLHAOGBK_00 311 and LLHAOGBK_00 328) (Fig. 3). The transposon (GenBank OR192611) is also found in several Streptococcus and Enterococcus genome assemblies (data not shown), and was assigned the number Tn7696 by the transposon registry (Tansirichaiya et al. 2019). Compared to Tn916, Tn7696 has a 120-bp in-frame deletion in the gene encoding a putative endopeptidase p60 precursor (ORF14 in Tn916, LLHAOGBK_00 319 in L-NTCD03) as well as several SNPs.
L-NTCD03 harbors the novel transposon Tn7696. Comparison of a region of the genome of L-NTCD03 with the genome of the closely related clade 4 strain DSM 29629 (GenBank NZ_CP016104). The insertion of Tn7696 into the pdeR gene (LLHAOGBK_00 328 and LLHAOGBK_00 311/CDIF29629_RS01565) is evident. Tn7696 harbors the tetracycline resistance gene tet(M) and an integrase (int-Tn), like Tn916. The gene with locus tag LLHAOGBK_00 319 (see main text) is abbreviated to 319 for simplicity. Colors and linkages indicate homology between genes. nox encodes a predicted NADH dehydrogenase/nitroreductase (LLHAOGBK_00 337/CDIF29629_RS01610), uvrA an excinuclease (LLHAOGBK_00 310/CDIF29629_RS01560) and guaA a glutamine-hydrolyzing GMP synthase (LLHAOGBK_00 305/CDIF29629_RS01525).
Detailed analysis of the genomic region of the cfr(B) gene in L-NTCD03 showed that it is carried on a putative mobile genetic element similar, but not identical, to the Tn6218-like element reported in strain Ox3196 (GenBank HG002389). This element is distinct from the Tn6218-like element of strain Ox2176 (GenBank HG002396) and the integrative and conjugative element ICECd-cfr(B) (GenBank PV037680.1) which also have been reported to carry cfr(B) in C. difficile (Dingle et al. 2014, Stojkovic et al. 2019, Brenciani et al. 2025).
Finally, we employed two tools to predict i) putative phages (Wishart et al. 2023) and ii) genomic islands (Bertelli et al. 2017). This identified two possible complete (pro)phages in L-NTCD03, with homology to phiCDHM19 (NC_028 996; bp 664 975–719 994) and phiC2 (NC_09 231; bps 2 626 715–2 687 566) both of which are also predicted with high confidence by geNomad (Camargo et al. 2024), and two large (bp 490 582–569 665 and 3 900 143–3 992 891) plus several small regions that are likely horizontally acquired.
Overall, the complete genome sequence suggests a clear relation with other clade 4 C. difficile isolates, and a significant contribution of horizontal gene transfer to its genetic content.
Discussion
Here, we provided a characterization of L-NTCD03, isolated from a healthy elderly subject. We confirmed that this isolate does not express the large clostridial toxins tcdA and tcdB, using ELISA, and showed that supernatants derived from an L-NTCD03 culture do not induce a loss of TEER in a trans-well model of colonic epithelium. Additionally, data from the swine IsoLoop model suggest that there is no induced pathology in loops inoculated with L-NTCD03 compared to controls. The loop experiment provides insight into strain behavior within a translationally relevant, C. difficile–predisposing intestinal environment, using a customized dysbiotic gut segment that enables precise local control of the bacterial inoculum, bile acid milieu, and other key microenvironmental parameters. The data is in line with the genomic analysis, which shows a lack of identifiable toxin genes, including the binary toxin locus. Together, these data strongly suggest that L-NTCD03 is unlikely to induce disease.
The high-quality complete genome generated for L-NTCD03 as part of this work, allowed us to place the PCR ribotype 416 strain in clade 4. The most closely related publicly available genome at the time of analysis is DSM 29629 (Riedel et al. 2017) (CP016104.1). Like L-NTCD03, this strain was non-toxigenic and assigned to clade 4, however, it is listed as ribotype SLO 235. At present it is unknown if the ribotyping patterns for SLO 235 and RT416 (as determined by capillary ribotyping and assigned at the CDRN) are similar or the same. Interestingly, in the manuscript describing DSM 29629 the authors note metabolic versatility between 17 different C. difficile strains that does not appear to necessarily follow the 5 major phylogenetic clades (clade) or PCR ribotypes tested. In the present work, we find that L-NTCD03 demonstrated a carbon utilization profile that is quite similar to that of other C. difficile strains under the conditions tested. We have to note that this was assessed at a single concentration and we cannot rule out that differences may exists at alternative concentrations, similar to what has been noted for other C. difficile strains belonging to other ribotypes (Collins et al. 2018, Midani et al. 2025).
We have determined the phenotypic antimicrobial resistance profile for 17 antimicrobials and found that several could be explained by genetic determinants identified in the complete genome of L-NTCD03. Some of these determinants appear to be associated with mobile genetic elements (Bannam et al. 1995, Derbise et al. 1997, Soge et al. 2008, Hua et al. 2021), supporting the view that NTCD strains may act as a reservoir for resistance determinants (Imwattana et al. 2020). As observed also by others for NTCD strains (Imwattana et al. 2020), we also observed resistance to rifampicin and ciprofloxacin in L-NTCD03. It is interesting to speculate that high levels of resistance in NTCD may relate to prolonged carriage, or reflect antibiotic use in geographic areas where these strains are common. Rifampicin, for instance, is commonly used in the treatment of tuberculosis patients. For clade 4 strains, high prevalence in Asia has been noted (Imwattana et al. 2020) but such strains are also more broadly found (Viprey et al. 2022). Notably, though the nimB-Cd resistance gene was identified (Supplemental Table 2), we observed no resistance to metronidazole. Consistent with this, nimB is commonly found in susceptible isolates of C. difficile and L-NTCD03 lacks the PnimB^G^ mutation associated with increased nimB expression and inducible metronidazole resistance (Olaitan et al. 2023). It also highlights the putative nature of automated identification of antimicrobial resistance determinants.
Finally, we identified the cfr(B) gene in L-NTCD03. The cfr(B) gene encodes a rRNA methyltransferase that confers multidrug resistance by ribosomal target modification and, in addition to C. difficile, has been found in Enterococcus spp and methicillin-resistant Staphylococcus aureus. At present there is no evidence that this particular element is transferable to the species in which cfr(B) carriage is potentially problematic. Indeed, the sequence of the Tn6218-like element from Enterococcus faecium (GenBank KR610408) appears to be quite different from the element in L-NTCD03, as also observed by others (Bender et al. 2016), supporting this notion. Interestingly, even for ICECd-cfr(B), that contains a clear conjugation module, authors failed to show transfer (Brenciani et al. 2025). We showed for the first time that cfr(B) can confer resistance to clindamycin (>256 mg/l), linezolid (12 mg/l), repatamulin (16 mg/l), and streptogramin A (16 mg/l) in the RT012 laboratory strain 630Δerm (van Eijk et al. 2015). Our data confirm that cfr(B) is a legitimate PhLOPSa (phenicol, lincosamide, oxazolidinone, pleuromutilin, streptogramin A) resistance gene. We were unable to establish whether cfr(B) plays a role in phenicol resistance, though, as L-NTCD03 also contains the catP (chloramphenicol resistance gene) and the plasmid we used to introduce the cfr(B) gene in our laboratory strain also carries a catP resistance determinant (Fagan and Fairweather 2011, Oliveira Paiva et al. 2016). We have, however, independently observed that catP-negative clinical isolates that carry cfr(B) demonstrate elevated chloramphenicol MICs compared to isolates that lack cfr(B), suggesting that cfr(B) also affects chloramphenicol MICs. Though it has previously been shown that cfr gene carriage can be associated with linezolid resistance in C. difficile (Marin et al. 2015, Stojkovic et al. 2019) or lead to an increase in MIC for certain antibiotics when expressed in a heterologous host (Hansen and Vester 2015), to our knowledge, the present study is the first direct demonstration of this function for cfr(B) as a PhLOPSa resistance gene in this organism.
Overall, the safety profile of L-NTCD03 suggests that it may be suitable as a live biotherapeutic product if the antimicrobial resistance pattern of the strains is deemed acceptable or even desirable for its specific application. The strain has successfully been used in a controlled human colonization experiment approved by a medical research ethics committee (https://clinicaltrials.gov/study/NCT05693077).
Supplementary Material
xtag010_Supplemental_Files
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aktories K, Schwan C, Jank T. Clostridium difficile toxin biology. Annu Rev Microbiol. 2017;71:281–307. 10.1146/annurev-micro-090816-093458.28657883 · doi ↗ · pubmed ↗
- 2Aspinall ST, Hutchinson DN. New selective medium for isolating clostridium difficile from faeces. J Clin Pathol. 1992;45:812–4. 10.1136/jcp.45.9.812.1401214 PMC 495111 · doi ↗ · pubmed ↗
- 3Baktash A, Corver J, Harmanus C et al. Comparison of whole-genome sequence-based methods and pcr ribotyping for subtyping of Clostridioides difficile. J Clin Microbiol. 2022;60:e 0173721. 10.1128/JCM.01737-21.34911367 PMC 8849210 · doi ↗ · pubmed ↗
- 4Bannam TL, Crellin PK, Rood JI. Molecular genetics of the chloramphenicol-resistance transposon tn 4451 from clostridium perfringens: the tnpx site-specific recombinase excises a circular transposon molecule. Mol Microbiol. 1995;16:535–51. 10.1111/j.1365-2958.1995.tb 02417.x.7565113 · doi ↗ · pubmed ↗
- 5Bayne J, Charavaryamath C, Hu Y et al. The swine isoloop model of the gut host-microbiota interface enables intra-animal treatment comparisons to advance 3r principles. Gut Microbes. 2025;17:2568706. 10.1080/19490976.2025.2568706.41137517 PMC 12562777 · doi ↗ · pubmed ↗
- 6Bender JK, Fleige C, Klare I et al. Detection of a cfr(b) variant in German enterococcus faecium clinical isolates and the impact on linezolid resistance in enterococcus spp. P Lo S One. 2016;11:e 0167042. 10.1371/journal.pone.0167042.27893790 PMC 5125667 · doi ↗ · pubmed ↗
- 7Bertelli C, Laird MR, Williams KP et al. Islandviewer 4: expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017;45:W 30–5. 10.1093/nar/gkx 343.28472413 PMC 5570257 · doi ↗ · pubmed ↗
- 8Boekhoud IM, Hornung BVH, Sevilla E et al. Plasmid-mediated metronidazole resistance in Clostridioides difficile. Nat Commun. 2020;11:598. 10.1038/s 41467-020-14382-1.32001686 PMC 6992631 · doi ↗ · pubmed ↗