Campylobacter jejuni infection impacts host-derived miRNAs targeting bacterial and host genes
Siddhi Chitre, Raad Z. Gharaibeh, Rachel C. Newsome, Jinmai Jiang, Andrew Brock, Thomas D. Schmittgen, Christian Jobin

TL;DR
This study shows that Campylobacter jejuni infection changes the host's miRNA profile, which may influence both host and bacterial genes in the gut.
Contribution
The study reveals how C. jejuni alters host EV-derived miRNA profiles, potentially modulating bacterial and host gene expression.
Findings
C. jejuni exposure significantly altered miRNA expression compared to other groups.
EV-derived miRNAs targeted host genes involved in immune regulation and inflammation.
Some miRNAs may bind to bacterial genes, suggesting a role in host-microbe communication.
Abstract
The intestinal microbiota has been shown to influence host-derived microRNAs (miRNAs), small regulatory RNAs involved in host-pathogen interactions. However, the specific role of individual bacterial species in modulating host extracellular vesicles (EV)-derived miRNA profiles remains unclear. We compared stool miRNA profiles from three groups: (i) Germ-free (GF) mice, (ii) mice colonized with a consortium of 13 bacteria (C13), and (iii) mice colonized with C13 and Campylobacter jejuni 81–176 (C13 + C. jejuni). miRNA profiling revealed unique differential expression of 8 miRNAs in C13 vs GF, 38 in C13 + C. jejuni vs C13, and 159 in C13 + C. jejuni vs GF (Padj < 0.05), indicating a substantial shift in host miRNA landscape following C. jejuni exposure. Notably, C13 + C. jejuni mice showed increased expression of miRNAs predicted to target bacterial virulence genes, suggesting a potential…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
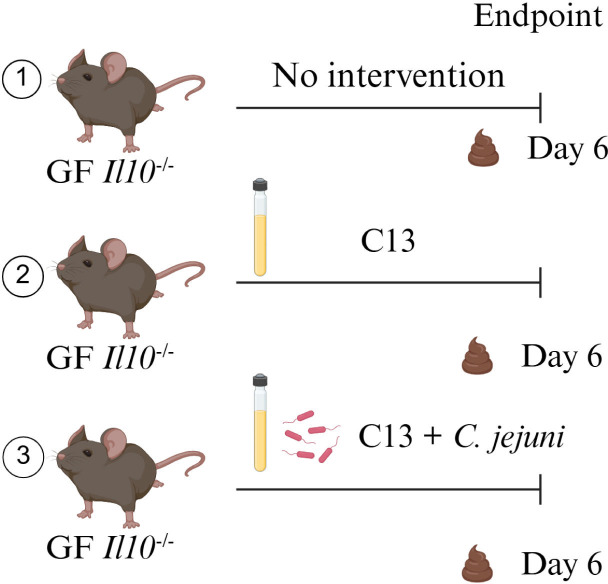 Fig 1
Fig 1 Fig 2
Fig 2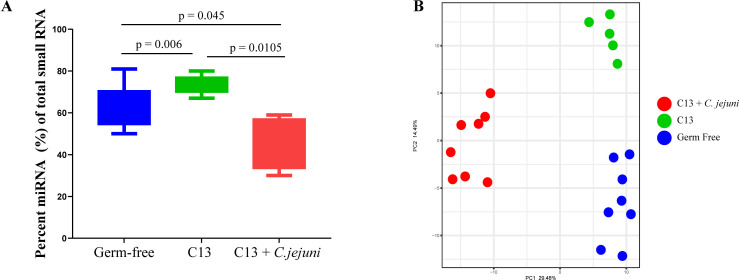 Fig 3
Fig 3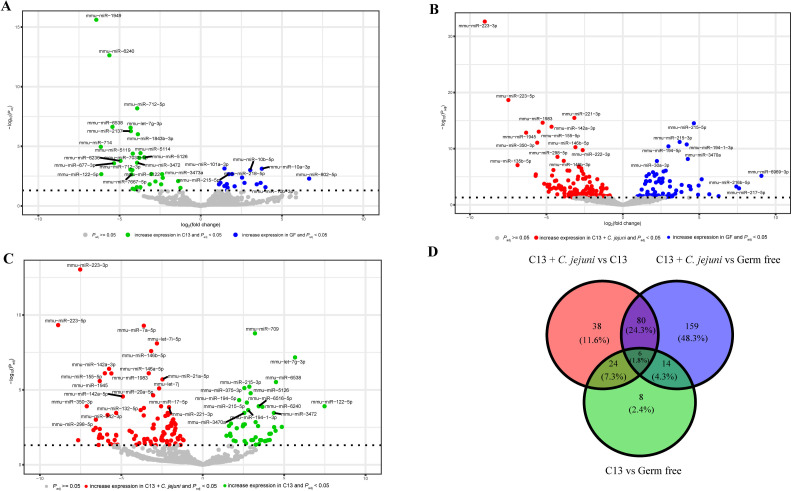 Fig 4
Fig 4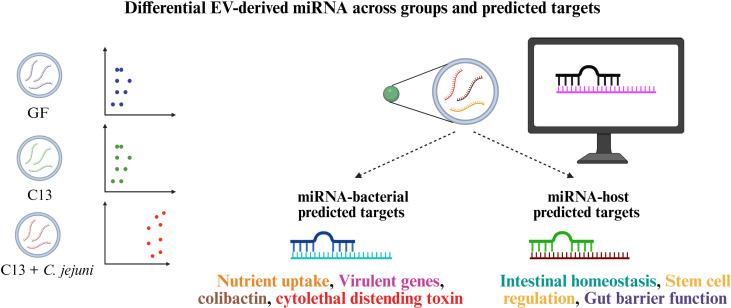 Fig 5
Fig 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Bacterial Infections and Vaccines · MicroRNA in disease regulation
INTRODUCTION
A regulated interaction is essential between the host and the gut microbiota to maintain gut homeostasis and disruption of this interaction leading to microbial dysbiosis can cause various gastrointestinal diseases (1). This homeostatic host-microbiota communication is influenced by numerous extrinsic factors such as diet (2), stress (3), antibiotic exposure (4), and pathogenic infection (1) and host-derived factors such as mucus (5), immunoglobulin A (6), anti-microbial peptides (7), and recently miRNA (8, 9). miRNA-mediated communication has been shown to play a critical role in maintaining healthy gut homeostasis (10), but their dysregulation is also associated with inflammatory bowel disease (IBD) (11). Interestingly, bacterial pathogens can affect the expression of the host miRNAs (12). A previous study showed that host-derived miRNAs (mmu-miR-515-5p and mmu-miR-1226-5p) promoted the growth of F. nucleatum and E. coli, respectively (13). Interestingly, the profile of EV-containing miRNA was altered in the feces of rats exposed to the inflammatory compound dextran sodium sulfate (DSS) compared to healthy rats (14). The authors also demonstrated that treatment with protective miRNAs (e.g., miR-200b-3p) alters the gut microbiome, increasing the abundance of specific beneficial bacteria such as Lactobacillus and reducing the abundance of potentially harmful bacteria like Escherichia (14). We have previously established that mucosal-associated bacterial communities obtained from biofilm-positive or negative colorectal cancer (CRC) patients differentially affect the production of host-derived miRNA and CRC development in a preclinical model (15). However, our understanding of the role of specific bacteria in influencing miRNA production in vivo and subsequent gene targets is limited.
Campylobacteriosis caused by Campylobacter jejuni (C. jejuni), although categorized as a self-limiting disease, is a significant cause of concern due to the presence of various virulent factors (16, 17). Studies have indicated the enrichment of Campylobacter species in IBD (18) and within tumor and feces of CRC patients (19–22). Recent studies have also demonstrated the presence of C. jejuni virulence genes cdtA, B, and C in diarrheal patients (23, 24). We have previously demonstrated that C. jejuni-derived CDT has tumorigenic and metastatic properties in pre-clinical models (25, 26). Since campylobacteriosis is one of the most prevalent forms of bacteria-induced gastroenteritis, it is important to understand the detailed host responses to this pathogen. We previously established a consortium of 13 bacteria (C13) representing the dominant bacterial phyla (27) in the mouse gut as a tractable system to study host response to Campylobacter jejuni 81–176 infection in GF Il10^−/^^−^ mice. We observed that C. jejuni infection of C13-colonized mice triggered host inflammatory gene expression and increased E. coli colonization in the distal colons compared to C13 alone (27). In this study, we demonstrated that the extracellular vesicle (EV)-derived miRNA profile is altered by C. jejuni infection, which is associated with the production of miRNAs predicted to bind bacterial virulence gene targets. We also leveraged EV miRNA profiling and integrated these data with spatial transcriptomics to examine how C. jejuni infection modulates host EV-derived miRNAs and their targeting of gut epithelial genes, providing insights into host-microbiota communication and pathogen-mediated shifts in microbial and epithelial dynamics.
RESULTS
Stool-derived extracellular vesicles retain size and structural integrity and are free from bacterial RNA
We previously found that biofilm-positive or negative bacteria differentially affect the production of host-derived miRNA in a CRC preclinical model (15). The establishment of a bacterial consortium sensitive to the presence of C. jejuni (27) represents a unique opportunity to interrogate the impact of bacteria on host miRNA expression. The inflammation status of the three groups, GF, C13 colonized, and C13 + C. jejuni-infected mice, has been characterized previously (27) and showed minimal inflammation in GF (mean inflammation score 0.8) and C13-colonized mice (mean inflammation score 1.5), and pronounced mucosal damage, goblet cell loss, and immune cell infiltration in C13 + *C. jejuni-*infected mice (mean inflammation score 4). Using this system, we sought to understand the impact of this pathogen on host miRNA. To investigate this question, we isolated and characterized the EVs purified from the feces of GF, C13, and C13 + C. jejuni-colonized mice (Fig. 1). EVs were isolated by ultracentrifugation and subjected to the nanoparticle tracking analysis system. This analysis detected particles ranging from 33 to 200 nm, with an average of 121.6 nm in size, with two detected peaks at 103 nm and 154 nm, compatible with EV phenotype (Fig. 2A). The cryo-TEM microscopy platform revealed a classic cup shape morphology with a lipid bilayer demonstrating the presence of EVs in the feces of these mice (Fig. 2B). To confirm that the stool EV isolations are free of bacteria and bacterial outer membrane vesicles (OMVs), we used RT-qPCR to analyze the stool EV RNA for eukaryotic and prokaryotic transcripts. The expression levels of these markers confirmed that our EV purification was free from any prokaryotic RNA contamination, as demonstrated by Ct values below the limit of detection for bacterial genes and positive for host eukaryotic markers (Fig. 2C).
Experimental workflow for investigating host-bacterial communication in GF, C13, and C13 + C. jejuni infection.
EV-derived miRNA demonstrates a differential miRNA profile. (A) The particle size distribution and concentration of EVs were assessed using NTA. (B) Cryo-TEM images of mouse stool EVs isolated scale bar size = 100 nm. (C) Gene expression of mammalian and prokaryotic gene transcripts in stool-derived EVs. The dashed line indicates the limit of detection for the RT-qPCR.
Host-derived miRNA profile is sensitive to bacterial composition
To study the impact of C. jejuni infection on host EV-derived miRNA profile, we extracted and sequenced miRNA from purified EVs from the feces of GF, C13, and C13 + C. jejuni-colonized mice. We observed that C. jejuni modulates the miRNA profile of mouse stool EVs. Of the total small RNA from EVs, the C13 + C. jejuni feces contained 46% miRNA, whereas EV from the feces of GF and C13 mice had an average of 64% and 74% miRNA, respectively (Fig. 3A; Table S1: https://figshare.com/s/824c92cec2bd7b18bd4e). Principal Component Analysis (PCA) of remaining miRNAs showed three distinct clusters corresponding to the three groups (GF, C13, and C13 + C. jejuni) (Fig. 3B), suggesting that the presence of specific bacteria influences miRNA expression profile.
The presence of C. jejuni in C13 altered miRNA profile in mice. (A) Quantification of miRNA isolated from fecal EVs across all groups GF (n = 9), C13 (n = 6), C13 + C. jejuni (n = 8). Padj-values from R’s two-sample Wilcoxon test after multiple testing correction. (B) PCA plot showing sample clustering based on their miRNA profiles in GF (n = 8), C13 (n = 5), and C13 + C. jejuni (n = 8) groups. Differences in sample clustering were tested using R’s gls: C13 vs GF Padj value = 2.48 × 10−6, C13 + C. jejuni vs GF Padj value = 1.71 × 10−12, and C13 + C. jejuni vs C13 Padj value = 1.36 × 10−8. Statistical significance was defined as P < 0.05.
C. jejuni infection induces unique host EV miRNA expression patterns
To further define differential miRNA responses in this cohort, we analyzed expression patterns across GF, C13, and C13 + C. jejuni groups. We observed that C13 colonization significantly induces the expression of different miRNAs as compared to GF (Fig. 4A). Similarly, C13 + C. jejuni induces the expression of different miRNAs as compared to GF (Fig. 4B). Furthermore, the addition of C. jejuni to the C13 strongly modifies the miRNA expression (Fig. 4C). To further characterize the significantly differentially expressed miRNAs between the three groups, we compared the overlapping miRNA between the three comparisons and identified 8 miRNAs that are unique to the C13 vs GF comparison, 38 unique to C13 + C. jejuni vs C13 comparison, and 159 unique to C13 + C. jejuni vs GF comparison (Fig. 4D). This suggests that the presence of C. jejuni altered both the number and the identity of the significant and differentially expressed miRNAs compared to GF and C13 alone.
Distinct miRNA signatures observed in GF (n = 8), C13 (n = 5), and C13 + C. jejuni (n = 8) infected mice. (A) Volcano plot of differentially expressed miRNAs in C13 vs GF. Horizontal dotted line showing −log10(0.05). (B) Volcano plot of differentially expressed miRNAs in C13 + C. jejuni vs GF. Horizontal dotted line showing −log10(0.05). (C) Volcano plot of differentially expressed miRNAs in C13 + C. jejuni vs C13. Horizontal dotted line showing −log10(0.05). (D) Venn diagram representing shared and unique miRNAs that are significantly and differentially expressed in the three experimental groups. Statistical significance was defined as P < 0.05.
Relationship between EV-derived miRNA with mammalian and bacterial gene target
miRNA sequencing revealed differentially expressed host miRNAs across all three conditions (Table S2: https://figshare.com/s/824c92cec2bd7b18bd4e). Using Probability of Interaction by Target Accessibility (PITA), we predicted several host and bacterial binding partners for these miRNAs. To evaluate possible regulatory effects, we compared the predicted targets with gene expression profiles from a previously generated fecal RNA-seq data set from the same experimental study (27). Although multiple miRNAs were differentially expressed, we highlight four miRNAs that were significantly upregulated and downregulated in the C13 + C. jejuni colonized mice and analyzed their predicted mouse gene transcript targets (Table 1, Table S3: https://figshare.com/s/824c92cec2bd7b18bd4e).
TABLE 1: miRNAs differentially altered in C13 + C. jejuni with their predicted gene targetsa,b
We then investigated the presence of any EV-derived miRNA that can regulate bacterial genes. Due to the large data sets produced by PITA analysis, we focused on two miRNAs with the highest expression in each group that were either upregulated or downregulated in C13 + C. jejuni or C13 (Table S4: https://figshare.com/s/824c92cec2bd7b18bd4e). The miRNA mmu-miR-223-3p was significantly overexpressed in C13 + C. jejuni-infected mice. PITA analysis predicted that this miRNA could bind several bacterial genes. Examination of these bacterial genes in our previously published RNA-seq data set revealed that their expression of bacterial genes corresponding to nutrient transport, iron uptake, metabolism, and cell wall biosynthesis was decreased in C13 + C. jejuni compared to C13, suggesting a potential repressive effect of mmu-miR-223-3p on these targets. Interestingly, we also observed an increase in the bacterial virulence associated with gene transcripts including iron uptake and chemotaxis in E. coli (Table 2).
TABLE 2: mmu-miR-223-3p significant and overexpressed in C13 + C. jejuni
Conversely, mmu-miR-709, which was downregulated in C13 + C. jejuni-infected mice, was predicted by PITA to target several bacterial gene transcripts. Examination of these transcripts in our previously published RNA-seq data set showed increased expression of genes involved in nutrient uptake, amino acid metabolism, DNA binding and repair, and energy metabolism across the C13 consortium. In C. jejuni, transcripts encoding flagellar proteins were also elevated, suggesting enhanced motility in response to infection (Table 3).
TABLE 3: mmu-miR-709 significant and downregulated in C13 + C. jejuni
We observed that in C13, mmu-miR-6240 is associated with an increased expression of bacterial gene transcripts involved in nutrient uptake, signal transduction, and transport, which maintain bacterial homeostasis. Following C13 + C. jejuni infection, mmu-miR-6240 was downregulated, which may have resulted in increased expression of gene transcripts responsible for metabolism, fatty acid biosynthesis, and amino acid metabolism in C13 + C. jejuni (Table 4).
TABLE 4: mmu-miR-6240 significant and overexpressed in C13
Next, we focused on mmu-miR-10a-3p, which is downregulated in C13, leading to increased expression of bacterial gene transcripts involved in protein synthesis and ATP hydrolysis for substrate transporters across the C13 consortium. With this downregulation, we also observed a significant increase in the bacterial gene expression related to nutrient uptake, increased bacterial adhesion and colonization in the hosts, bacterial motility, adhesion, and DNA replication (Table 5), to name a few biological functions. Collectively, these analyses suggest that C. jejuni infection alters the host EV miRNA profile, which, in turn, may influence bacterial gene expression and contribute to shifts in gut microbiota composition.
TABLE 5: mmu-miR-10a-3p significant and downregulated in C13
Interestingly, our analysis showed 25 unique miRNAs significantly upregulated in C13 and predicted to target E. coli pks genes such as clbB, clbD, clbF, clbG, clbH, clbJ, clbM, and clbQ. Correspondingly, these genes were all significantly upregulated in the feces of C13 + C. jejuni colonized mice (Table S5: https://figshare.com/s/824c92cec2bd7b18bd4e). Interestingly, analysis of C. jejuni cdt gene transcripts revealed significant upregulation in the C13 + C. jejuni group. In this group, we observed that mmu-miR-28a-3p (predicted to bind to C. jejuni cdtA) and mmu-miR-1981-5p (predicted to bind to C. jejuni cdtA and cdtC) were also significantly upregulated. Additionally, mmu-miR-680, which was significantly upregulated in C13, was predicted to target cdtC. These observations suggest that host miRNAs may influence the expression of C. jejuni toxin genes, potentially contributing to host protection or, alternatively, modulating bacterial gene expression. Further experimental studies are needed to clarify the functional impact of these miRNAs.
These findings showed that the addition of C. jejuni to a defined consortium drastically modified host and bacterial gene expression, which is accompanied by altered expression of miRNAs that are predicted to interact with both mammalian and bacterial genes.
Spatial transcriptomics reveal stool-derived EV miRNAs targeting key gut epithelial genes
Using spatial transcriptomics data from our previous work (27), we observed that C. jejuni colonization induced region-specific transcriptional changes in the colonic tissue depending on the bacterial abundance. The regions analyzed were defined based on the presence or absence of C. jejuni in the colon*,* as determined by fluorescence in situ hybridization (FISH) staining assay (27). Notably, some regions were relatively depleted of C. jejuni, which may reflect local differences in microbial colonization, mucus layer composition, or epithelial susceptibility. To determine whether host EV-derived miRNAs contributed to these changes, we integrated spatial transcriptomics results with stool EV miRNA profiles and focused on experimentally validated miRNA–mRNA interactions (Table S6: https://figshare.com/s/824c92cec2bd7b18bd4e). This analysis revealed several miRNAs with distinct expression patterns, each predicted to target key genes involved in immune activation, epithelial repair, or inflammation. Among them, mmu-miR-155-5p was the most prominent, showing marked upregulation in C13 + C. jejuni mice. It was predicted to regulate transcripts including Saa3, Il1a, Ido1, Lyz2, and S100a8, all genes associated with pro-inflammatory or antimicrobial responses. Consistent with this, regions enriched in C. jejuni exhibited strong upregulation of Saa3, Il1a, Lyz2, and S100a8, whereas Ido1, an immune tolerance gene, was significantly downregulated in *C. jejuni-*depleted tissue regions.
mmu-miR-122-5p, which was upregulated in the C13 group but reduced in C13 + C. jejuni mice, was predicted to bind Cxcl9, Krt19, Ccl5, and Kctd12. Notably, Cxcl9, Ccl5, and Kctd12 expression was elevated in C13 + C. jejuni mice, suggesting that diminished miR-122-5p expression may relieve repression on Cxcl9, Ccl5, and Kctd12, enhancing immune chemokine signaling. Similarly, Krt19 genes involved in epithelial integrity were downregulated in regions lacking C. jejuni, indicating spatially distinct regulatory mechanisms.
In addition, mmu-miR-21a-5p was significantly increased in C13 + C. jejuni mice, with its target gene transcript S100a8 also showing elevated expression in C. jejuni-enriched regions, implicating this miRNA–mRNA pair in pro-inflammatory signaling. Similarly, mmu-let-7i-5p and its target Itgb2 were both upregulated in C. jejuni-enriched areas, indicating coordinated regulation of immune cell adhesion pathways during bacterial colonization. mmu-miR-142a-3p was upregulated in C13 + C. jejuni, whereas its predicted target Cystm1 was reduced in C. jejuni-depleted tissue regions, suggesting a potential role in modulating epithelial defense mechanisms.
Taken together, these findings demonstrate that C. jejuni colonization reshapes the host EV-miRNA landscape in a spatially resolved manner. The enrichment of miRNAs such as miR-155-5p and the loss of regulatory miRNAs like miR-122-5p suggest a coordinated mechanism by which host-derived miRNAs modulate immune and epithelial gene expression in response to C. jejuni colonization.
DISCUSSION
The relationship between host response to enteric pathogens and induction of luminal miRNAs is poorly understood. We previously established a novel tractable system for studying C. jejuni host response in the presence of a curated consortium of 13 bacteria (C13) representing four dominant phyla in the murine gut (27). This established consortium provided a foundation for the current investigation, to understand previously unidentified host responses.
In this study, we observed that the C13 + C. jejuni group had significantly less miRNA encapsulated in fecal EVs, suggesting that the pathogen may interfere with the biogenesis of EVs and/or miRNA. This finding suggests a potential role for C. jejuni in altering the miRNA content within the EVs, as seen with infection with other pathogenic bacteria (12). We observed an upregulation of miRNAs such as mmu-miR-10b-5p and mmu-miR-215-5p in GF mice, which have a role in maintaining gut homeostasis. For example, a global deficiency of mmu-miR-10b-5p leads to alterations in gut functions such as increased intestinal permeability, dysregulation of tight junctions (28), and disruption of gut immune response (29). mmu-miR-215-5p plays a role in the host by suppressing inflammation and by repressing the inflammatory Interleukin Enhancer Binding Factor 3 (ILF3) and LRR binding FLII interacting protein 1 (LRRFIP1) (30). These data suggest that while GF mice express miRNAs involved in gut homeostatic functions, they can be susceptible to infections due to the abundance of miRNAs that contribute to the suppression of inflammation, which is supported by previous reports that GF mice are prone to infectious diseases (31). These data suggest that a healthy gut microbiota (as established by the C13 consortium) leads to the production of specific miRNAs that maintain cellular homeostasis and protect from disease development. However, this symbiotic balance shifts to dysbiosis after the addition of the pathogenic C. jejuni to the consortium. We also observed an altered miRNA profile associated with inflammation and cancer metastasis. For example, mmu-miR-221-3p, mmu-let-7i-5p, mmu-miR-142a-3p are upregulated in the C13 + C. jejuni group and are associated with inflammation (32–34). Additionally, mmu-miR-223-3p and mmu-miR-7a-5p were increased in this group and are associated with the upregulation of Stat3 and Jak2 expression, respectively. Interestingly, He et al. (25) recently demonstrated that C. jejuni CdtB enhanced cancer metastasis through the JAK2/STAT3 signaling pathway, subsequently increasing the expression of pro-metastatic matrix metalloprotease 9 (MMP9). Thus, our data suggest a possible host-derived miRNA link in the mechanism of C. jejuni pathogenicity.
We identified miRNA and predicted their potential gene targets across the consortium. For example, mmu-miR-6240, which is significantly increased in C13, is predicted to bind to genes implicated in nutrient uptake, signal transduction, and transport across Blautia, Bacteroides, Enterococcus, Ligilactobacillus, and Parabacteroides suggesting a regulatory mechanism for maintaining bacterial homeostasis. Following C. jejuni inclusion, there is an imbalance in the regulation as seen by significant downregulation of another miRNA, mmu-miR-468-3p, which may lead to increased expression of pathogenic genes such as E. coli colibactin ClbH gene.
We also observed a significant increase of mmu-miR-223-3p in the C13 + C. jejuni group. This miRNA is predicted to bind to distinct gene transcripts involved in nutrient transport, iron uptake, metabolism, and cell wall biosynthesis, including Blautia, Bacteroides, Ligilactobacillus, and Parabacteroides suggesting a potential inhibitory mechanism that could cause a shift in gut microbiota composition. Additionally, mmu-miR-223-3p is predicted to bind to bacterial gene transcripts responsible for iron acquisition and chemotaxis across Akkermansia, Campylobacter, Escherichia, and Parabacteroides, suggesting a potential activation of virulent genes that would confer survival advantages over other bacteria. Interestingly, we observed that the presence of C. jejuni in the C13 led to an increased expression of miRNA mmu-miR-1981-5p, predicted to bind cdtA and cdtC transcripts. Host-derived miRNA targeting C. jejuni CDT subunits may represent a mechanism to counteract the pathogenic effects of CDT.
Integrating previously generated spatial transcriptomics data with host EV-derived miRNA profiles highlights a novel layer of host-microbiota communication. Spatial transcriptomics allowed us to identify key epithelial genes whose expression patterns potentially may be modulated by EV miRNAs, including mmu-miR-155-5p, mmu-miR-122-5p, mmu-miR-21a-5p, mmu-let-7i-5p, and mmu-miR-142a-3p. These miRNAs target genes involved in pro-inflammatory signaling, antimicrobial defense, chemokine-mediated immune recruitment, epithelial repair and barrier maintenance, immune cell adhesion, and immune tolerance, suggesting that host EVs can fine-tune epithelial responses to microbial colonization. Together, integrating spatial and host EV-derived miRNA data offers a powerful strategy to dissect host-microbiota crosstalk. We previously demonstrated that C. jejuni infection in GF Il10^−^/^−^ mice leads to rapid disruption of mucosal architecture, characterized by the loss of epithelial and goblet cells and marked immune cell infiltration by day 14 post-infection (35). In a separate study (36), acute ulcerative colitis in Il10^−^/^−^ mice was characterized by colonic ulceration, bleeding, diffuse mucosal and submucosal infiltrates, and loss of goblet cells and crypts by day seven post-infection. Together, these findings indicate that infection triggers both direct epithelial damage and secondary effects associated with inflammation and tissue remodeling. In this context, the EV changes observed in our study are likely to arise from multiple cellular sources. As our GF model does not allow precise cell-type attribution, it remains unclear whether epithelial or immune cells contribute most significantly to the EV pool. Future studies employing cell-type-specific EV isolation or labeling strategies to better define the origins and functional roles of EVs during C. jejuni infection. Nevertheless, our study provides the first characterization of EV-derived miRNAs in the context of C. jejuni infection. Future studies investigating the profile of EV-derived miRNA in C. jejuni mono-associated GF mice may provide additional information on the contribution of this pathogenic bacterium to host-microbe interaction.
Further investigation is also warranted to understand the functional implications of miRNA binding in homeostasis and pathogenic states. The differential profile of miRNAs generated by the host upon colonization status (GF, C13, and C13 + C. jejuni) highlights their potential significance in regulating host-microbe interaction. The main limitation of this study is the correlative nature of the complex miRNA-bacterial interactions presented in the paper. The data are based on in silico analysis, and functional experiments will be needed to demonstrate cause-effect relationships with candidate miRNAs. The synthesis of candidate miRNA and target delivery in bacterial cultures of individual C13 members may allow the assessment of bacterial gene expression in a controlled manner.
Conclusion
In conclusion, we demonstrate that C. jejuni infection alters fecal EV-derived miRNA profile (Fig. 5). The miRNAs abundantly expressed upon C. jejuni infection are predicted to bind to the bacterial virulence genes thereby modulating their function.
Graphical representation of significant findings. Fecal EV-derived miRNA sequencing from GF, C13, and C13 + C. jejuni group revealed distinct miRNA clustering. Predicted interactions between miRNAs and host or bacterial gene transcripts were identified using PITA analysis. These results highlight potential targets for further functional validation.
MATERIALS AND METHODS
Mice experiment
All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) at the University of Florida (UF) and performed at UF Animal Care Facilities (IACUC Protocol #202200000637). Mice fecal samples used in this study are a part of an experimental cohort previously described in a separate manuscript (27) from which stool samples were collected and analyzed here. Briefly, 6–10 week old, mixed gender GF Il10^−/−^ (129/SvEv) mice were divided into three groups: (i) GF, (ii) C13 consortium containing the four most prominent phyla present in the healthy mouse gut (C13), and (iii) C13 containing C. jejuni 81–176 (C13 + C. jejuni). Mice were gavaged with the different bacterial consortium (10^6^ CFU). C13 + C. jejuni colonized mice were monitored daily for the clinical signs of campylobacterosis, allowing us to capture EV-miRNA and transcriptional changes associated with active infection while minimizing severe pathology. Stool samples were collected on day 6 post-gavage for all the groups.
Isolation of EVs from stool samples of GF, C13, and C13 + C. jejuni infected mice
Fecal samples from GF, C13, and C13 + C. jejuni, snap-frozen in liquid nitrogen and stored in −80°C were utilized for this analysis. Due to limited stool quantity obtained from one mouse in C13 + C. jejuni group, we pooled this sample with the stool sample from other mouse within the same group to ensure sufficient EV yields, resulting in a final sample size of GF (n = 9), C13 (n = 6), and C13 + C. jejuni (n = 8). EVs were isolated via standard differential centrifugation and ultracentrifugation methodologies following an established mouse stool EV isolation protocol (37) but with some modifications (38). Briefly, flash frozen (stored in −80°C) mouse stool from individual mice was thawed on ice, and individual stool pellets were placed in 50 mL conical tubes with 15 mL protease inhibitor cocktail solution (one tablet per 10 mLs ice cold PBS) and incubated at 4°C for 30 min. After 30 min of soaking at 4°C, the tube was vortexed at maximum speed to fully break apart stool pellet. Stool suspension was centrifuged at 3,000 × g for 10 min at 4°C, and the supernatant was collected. An additional 10 mL of protease inhibitor cocktail tablet solution was added to the stool pellet, and the sample was vortexed for 5 min followed by another 3,000 × g for 10 min at 4°C. The supernatants (~25 mL) were pooled and centrifuged at 3,000 × g for 30 min. Pooled supernatants were filtered through fast-flow filter paper to remove the remaining stool debris. The flow through was centrifuged at 40,000 × g for 1.5 h to pellet the large EVs, and the supernatant was collected and filtered using 0.22 µm filter to remove microbes. To pellet the final small EVs, the 40,000 × g supernatant was ultracentrifuged at 167,000 × g for 2.5 h. This pellet was carefully washed twice with PBS and resuspended in ~50 µL PBS. EVs were freshly used or stored at −80°C until EV characterization and RNA isolation.
RNA isolation and quantitative gene expression
Total RNA was extracted from stool EVs isolated by UC using the miRNeasy Mini Kit (Qiagen) per the manufacturer’s instructions. Purified RNA was eluted in 30 µL RNase-free water and stored at −80°C until use. RNA concentration was determined using a ND-1000 Spectrophotometer (Nanodrop Technologies). Fifty nanograms of total RNA isolated from the stool EVs was treated with DNase (ThermoFisher) and a standard reverse transcription using MultiScribe Reverse Transcriptase with random hexamers and stem loop primers (ThermoFisher) to specific mature miRNAs and U6 were used to generate cDNA. The thermocycling conditions are as follows: 16°C for 30 min, 42°C for 30 min, and 85°C for 5 min. cDNA was diluted 1:33 and qPCR was performed on a QuantStudio 7 Flex Real-Time PCR System (Applied Biosystems). Detection using SYBR Green to in-house designed qPCR primers (18S, 16S, FliC, and FliC K12) and Taqman Assays (mature miRNAs and U6) targets (ThermoFisher). Gene expression analysis is presented as mean CT values in triplicate. qPCR primer sequences are as follows: (i) 18S FP 5′ GTAACCCGTTGAACCCCATT 3′, RP 5′ CCATCCAATCGGTAGTAGCG 3′. (ii) 16S FP 5′ CCAGCAGCCGCGGTAATAC 3′, RP 5′ TCAGATGCAGTTCCCAGGTTG 3′. (iii) FliC FP 5′ GTGACGGTACAGCGTTTGATG 3′, RP 5′ AGCAGCAGAACCTGTTGTTACG 3′. (iv) FliC K12 FP 5′ TGGTGCTACCACCACAAACAA 3′, RP 5′ ATCACCACCGGTGATTTTCG 3′.
Nanoparticle tracking analysis
The high-resolution particle size distribution and concentrations of EVs were determined using NTA by calculating light scattering and Brownian motion properties. Briefly, isolated EVs were diluted 500-fold using ion-free PBS and injected into a Nanosight NS300 instrument (Malvern Instruments, Westborough, MA, USA) equipped with a blue488 laser and a sCMOS camera. The samples were measured for 60 s and five movies were taken for each sample with the single shutter (1,300) and gain (512) mode at 25 FPS. The analysis settings were (i) threshold: 6; (ii) blur size: auto; and (iii) max jump distance: Auto: 13.1–17.1 pix; temperature: 23.6–23.8°C. For determining particle concentration, dPBS was run as a blank for background particle concentration.
Cryo-transmission electron microscopy
Cryo-transmission electron microscopy (Cryo-TEM) from the University of Florida ICBR was used to image EVs. In brief, 3 µL of a suspension of concentrated mouse stool EVs was pipetted onto glow-discharged, C-Flat holey-carbon grids (Protochips Inc.). The grids were immediately vitrified in a slurry of liquid ethane cooled by liquid nitrogen in a Mark IV Vitrobot (FEI Co.) at 4°C and 95% humidity. The grids were examined, and images gathered on a FEI Tecnai G2 F20-TWIN microscope (RRID:SCR_019146) operated at 200 kV and ~20 e−/Å2 dosage (low dose). The scale bars are 100 nm.
Library construction and miRNA sequencing of stool EV RNA
miRNA sequencing was performed on the RNA isolated from individual stool pellets of GF group, C13 group, and C13 + C. jejuni group 6 days after bacteria colonization. In brief, total RNA from EVs was isolated following the manufacturer’s protocol of miRNeasy Mini kit (Qiagen). Isolated RNA was run on Agilent 2100 BioAnalyzer to determine the amount of miRNA from the total small RNA concentration. One sample each from the GF and the C13 groups was excluded from miRNA library preparation and sequencing due to low miRNA concentration (less than 10 ng) as determined by the Bioanalyzer, resulting in a final sample size of GF (n = 8), C13 (n = 5), and C13 + C. jejuni (n = 8). The amount of RNA used in library preparation was standardized to 10 ng of miRNA for each sample. miRNA sequencing libraries were synthesized and gel size-selected using the NEBNext Multiplex Small RNA Library Prep Set for Illumina (NEB). Samples were sequenced by the University of Florida NextGen DNA Sequencing (NS) core (RRID:SCR_019152) core on the Illumina Novoseq 6000 instrument (2 × 150 run, S4 Flow cell).
miRNA analysis
Data were analyzed as described previously (15). Briefly, CAP-miRSeq (39) was used to process the miRNA sequences as follows: reads were filtered and trimmed using cutadapt (40). Identification and quantification of miRNA were done using miRDeep2 (41), and differential miRNAs analysis was detected using edgeR. We considered a miRNA differentially expressed if its edgeR Padj -value < 0.05. PCA was created using R’s prcomp function from the normalized and log transform miRNA counts and differences in samples clustering were tested using gls (Generalized Least Squares model) function in R package nlme.
Bacterial target prediction for mouse miRNA was done using PITA (42); an miRNA target prediction tool that evaluates the thermodynamic free energy between the miRNA-target complex. We considered a bacterial gene a potential target for a particular mouse miRNA if its ΔΔG score < −10 kcal/mol. To analyze miRNA and mouse gene targets, multiMiR (http://multimir.org/) was used.
Statistical tests
We used R’s two-sample Wilcoxon test (also known as Mann-Whitney test) and for comparisons involving more than two, we corrected the P-values for multiple testing using R’s p.adjust function with method set to BH (Benjamini and Hochberg) (43).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hou K, Wu Z-X, Chen X-Y, Wang J-Q, Zhang D, Xiao C, Zhu D, Koya JB, Wei L, Li J, Chen Z-S. 2022. Microbiota in health and diseases. Signal Transduct Target Ther 7:135. doi:10.1038/s 41392-022-00974-435461318 PMC 9034083 · doi ↗ · pubmed ↗
- 2Ross FC, Patangia D, Grimaud G, Lavelle A, Dempsey EM, Ross RP, Stanton C. 2024. The interplay between diet and the gut microbiome: implications for health and disease. Nat Rev Microbiol 22:671–686. doi:10.1038/s 41579-024-01068-439009882 · doi ↗ · pubmed ↗
- 3Tofani GSS, Leigh S-J, Gheorghe CE, Bastiaanssen TFS, Wilmes L, Sen P, Clarke G, Cryan JF. 2025. Gut microbiota regulates stress responsivity via the circadian system. Cell Metab 37:138–153. doi:10.1016/j.cmet.2024.10.00339504963 · doi ↗ · pubmed ↗
- 4Strati F, Pujolassos M, Burrello C, Giuffrè MR, Lattanzi G, Caprioli F, Troisi J, Facciotti F. 2021. Antibiotic-associated dysbiosis affects the ability of the gut microbiota to control intestinal inflammation upon fecal microbiota transplantation in experimental colitis models. Microbiome 9:39. doi:10.1186/s 40168-020-00991-x 33549144 PMC 7868014 · doi ↗ · pubmed ↗
- 5Konrad A, Cong Y, Duck W, Borlaza R, Elson CO. 2006. Tight mucosal compartmentation of the murine immune response to antigens of the enteric microbiota. Gastroenterology 130:2050–2059. doi:10.1053/j.gastro.2006.02.05516762628 · doi ↗ · pubmed ↗
- 6Peterson DA, Mc Nulty NP, Guruge JL, Gordon JI. 2007. Ig A response to symbiotic bacteria as a mediator of gut homeostasis. Cell Host Microbe 2:328–339. doi:10.1016/j.chom.2007.09.01318005754 · doi ↗ · pubmed ↗
- 7Bevins CL, Salzman NH. 2011. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol 9:356–368. doi:10.1038/nrmicro 254621423246 · doi ↗ · pubmed ↗
- 8Goga A, Yagabasan B, Herrmanns K, Godbersen S, Silva PN, Denzler R, Zünd M, Furter M, Schwank G, Sunagawa S, Hardt W-D, Stoffel M. 2021. mi R-802 regulates Paneth cell function and enterocyte differentiation in the mouse small intestine. Nat Commun 12:3339. doi:10.1038/s 41467-021-23298-334099655 PMC 8184787 · doi ↗ · pubmed ↗