The role of OLE2 and POX1-3 in prostaglandin E2 production and virulence is conserved in Candidozyma (Candida) auris
Armand Bolsenbroek, Eduvan Bisschoff, Gabre Kemp, Olihile Sebolai, Carolina Pohl, Jacobus Albertyn

TL;DR
This study shows that Candida auris can produce PGE2, a lipid linked to virulence, and identifies key genes involved, offering potential new drug targets.
Contribution
First report of PGE2 production in C. auris and identification of OLE2 and POX1-3 as key genes in its virulence.
Findings
C. auris produces PGE2, a lipid linked to virulence in other yeasts.
OLE2 and POX1-3 are essential for PGE2 production and virulence in C. auris.
OLE2 deletion leads to monounsaturated fatty acid accumulation, suggesting it is not a Δ9 desaturase.
Abstract
Lipids have recently gained attention for their involvement in the virulence of pathogenic yeast. One group of lipids, prostaglandins, which are arachidonic acid-derived eicosanoids, has received particular attention. Prostaglandin E2 (PGE2) is the most studied prostaglandin in mammals. However, pathogenic yeast, including Candida albicans, Candida tropicalis, Candida glabrata, Candida parapsilosis, and Candida dubliniensis, also produces PGE2. Notably, PGE2 production has not yet been reported in Candidozyma auris (Candida auris). In C. albicans, PGE2 plays a crucial role in colonization and virulence by promoting yeast-to-hyphal transition, enhancing biofilm formation and colonization of the gastrointestinal tract. Although the biochemical pathway responsible for PGE2 synthesis in yeasts is unknown, several genes have been implicated, including OLE2 and POX1-3, which were investigated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
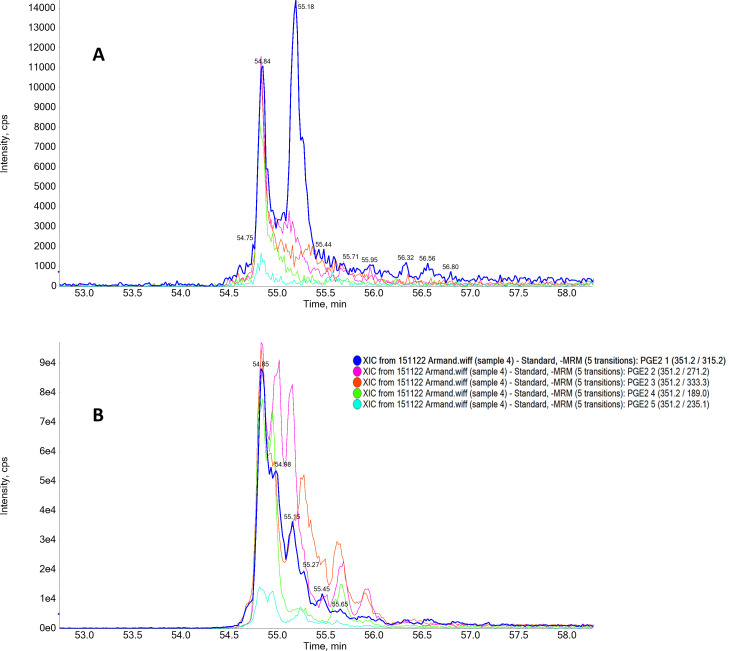 Fig 1
Fig 1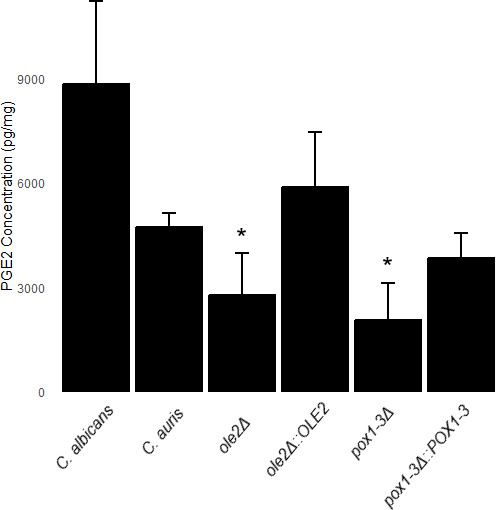 Fig 2
Fig 2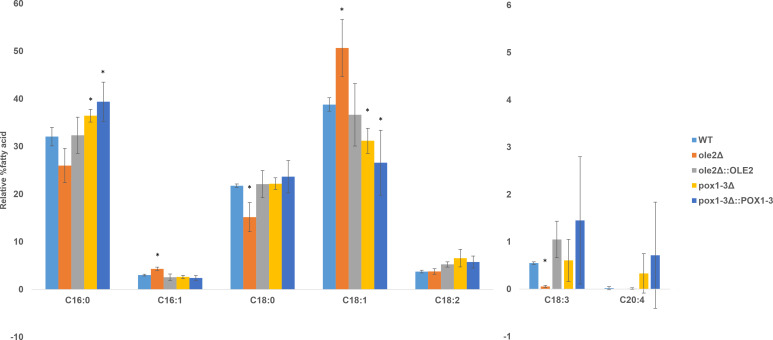 Fig 3
Fig 3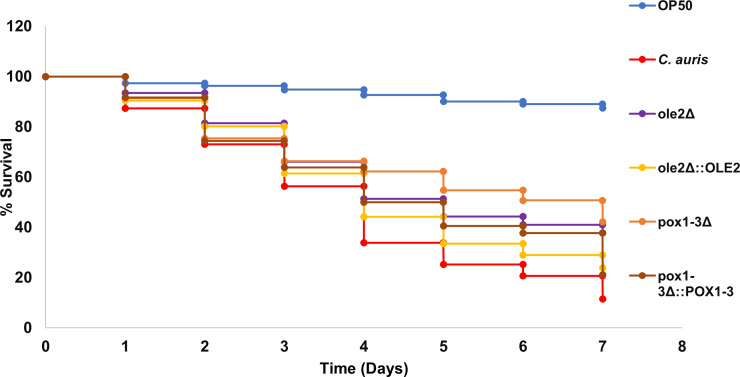 Fig 4
Fig 4| Species | Genotype | Parental strain | Description | Origin |
|---|---|---|---|---|
| Wild type | – | Reference strain | N/A | |
| Wild type | – | Clinical strain | N/A | |
|
| MRU293 | Deletion of | This study | |
|
| MRU293 | Add-back of | This study | |
|
| MRU293 | Deletion of | This study | |
|
| MRU293 | Add-back of | This study |
| Strain | Median lifespan (days) | Standard error | Days to reach 50% survival | Corrected Bonferroni |
|---|---|---|---|---|
| 4.12 | 0.26 | 4 | – | |
| 4.97 | 0.27 | 5 | 0.0168 | |
| 4.53 | 0.27 | 4 | 0.9638 | |
| 5.15 | 0.3 | 7 | 0.0054 | |
| 4.68 | 0.29 | 5 | 0.8736 |
| Organism | Entry | Gene name | Length | Identity |
|---|---|---|---|---|
| A0A1D8PHT9 | 526 aa | 56.6% | ||
| G8BJN8 | Cp | 550 aa | 55.1% | |
| 709 aa | 62.4% | |||
| G8BAZ7 | Cp | 711 aa | 60.5% |
- —National Research Foundation of South Africa
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Inflammatory mediators and NSAID effects · Fatty Acid Research and Health
INTRODUCTION
Eicosanoids derived from polyunsaturated fatty acids (PUFAs) such as arachidonic acid (AA), eicosapentaenoic acid, and docosahexaenoic acid include prostaglandins, thromboxanes, prostacyclins, and leukotrienes (1, 2) and represent a diverse and essential class of signaling lipids involved in a wide range of physiological and pathophysiological processes. The biosynthesis of prostaglandins in mammalian cells is a well-established process in which AA, a PUFA present in the cell membrane, is liberated by phospholipases and subsequently undergoes conversion by cyclooxygenase (COX) enzymes to generate prostaglandin H_2_ (PGH_2_). PGH_2_ serves as a common precursor for synthesizing diverse prostaglandins belonging to different classes, such as prostaglandin D, F, I, and E (1, 2). The expression and activity of distinct downstream enzymes, including prostaglandin synthases and isomerases, determine the ultimate production of specific prostaglandins.
The best studied of these eicosanoids is prostaglandin E_2_ (PGE_2_), which exhibits immunomodulatory effects in mammals in response to various stimuli, including pathogens (3). It can attenuate dendritic cell maturation and modulate the phagocytic activity of monocytes, macrophages, and neutrophils by selectively inhibiting their function. Additionally, PGE_2_ can suppress the Th1 inflammatory response, essential for host defense against Candida albicans infection, while promoting the Th2 and Th17 inflammatory responses (4–8).
In addition to mammals, eicosanoids can be biosynthesized by yeast and have been observed in pathogenic yeasts such as C. albicans. The first eicosanoid identified to be produced by C. albicans from exogenous AA was 3,18-dihydroxy-5,8,11,14-eicosatetraenoic acid (9). The production of PGE_2_ by C. albicans was also confirmed (10–12) and subsequently verified in several other Candida spp., such as Candida dubliniensis, Candida parapsilosis, Candida glabrata, and Candida tropicalis (13–16).
Interestingly, whole-genome sequencing indicated the absence of shared enzymatic pathways for prostaglandin synthesis between mammals and Candida spp. (11, 12, 17). In addition, the selective COX-2 inhibitor, CAY10404 (3-[4-methylsulfonylphenyl]−4-phenyl-5-trifluoromethylisoxazole), did not decrease PGE_2_ production in C. albicans, indicating that these yeasts utilize a distinct pathway for PGE_2_ synthesis (11). Further investigation revealed that the enzymes that play a role in PGE_2_ production in Candida spp. include a putative stearyl coenzyme A desaturase (encoded by OLE2), which is implicated in C. albicans. Homozygous deletion mutants lacking OLE2 show significantly decreased PGE_2_ production (11), as did deletion of genes of the high-affinity reductive iron acquisition pathway, such as FET99 (encoding a multicopper oxidase involved in iron transport) (18). Interestingly, in the case of C. parapsilosis, PGE_2_ production was not affected by deletion of OLE2 but was negatively influenced by deletion of FET3 (encoding a multicopper oxidase involved in iron transport), POX1-3 (encoding an acyl-CoA oxidase), and POT1 (encoding an acyl-CoA thiolase). This suggests that although there are similarities regarding the enzymes that influence PGE_2_ production in different Candida spp., there are also differences between them.
Candida auris, which was first identified in 2009 (19), is a newly emerged pathogenic yeast that poses a severe threat to hospital patients worldwide. It is highly persistent in healthcare facilities, causing multiple nosocomial outbreaks, and the rapid development of antifungal resistance limits the treatment options (20). This yeast was the first fungal pathogen categorized as a public health threat by the Centers for Disease Control and Prevention, and the World Health Organization has recently included C. auris in the highest group (critical) of priority fungal pathogens with C. albicans, Cryptococcus neoformans, and Aspergillus fumigatus (21). Thus, it is important to gain a better understanding of the biology of this yeast, including pathways involved in PGE_2_ production, which may serve as novel antifungal drug targets (22).
Therefore, this study aims to investigate the production of PGE_2_ in C. auris. Specifically, we assessed the involvement of OLE2 and POX1-3, previously linked to PGE_2_ production in certain Candida spp. Furthermore, we explored the potential contribution of these genes to C. auris virulence.
MATERIALS AND METHODS
Strains used
The strains that were used are listed in Table 1. C. albicans SC5314 was a kind gift of Prof. B. Hube (Leibniz Institute for Natural Product Research and Infection Biology—Hans Knöll Institute, Germany), and C. auris MRU293 (= B11224) was provided by Dr. Nelesh Govender (National Institute for Communicable Diseases, South Africa). C. auris strains and mutants were stored at −80°C in 30% glycerol and revived on yeast peptone dextrose (YPD; 10 g/L yeast extract, 20 g/L peptone, and 20 g/L glucose) at 30°C.
Construction of C. auris mutants
A published CRISPR-Cas9 system (23) for C. albicans was adapted for use in C. auris MRU293 by replacing C. albicans-specific sequences with C. auris sequences. This system introduces a double-strand break at the target gene, and then modification occurs through the donor DNA in the wild type. A detailed explanation of the methods to obtain these mutants can be found in the supplemental material. After deletion mutants (ole2Δ and pox1-3Δ) were generated, add-back strains were generated by reintroduction of the wild-type gene by modified donor DNA.
Biofilm formation and biomass determination
All strains were streaked onto yeast malt (YM) extract medium agar (3 g/L malt extract, 3 g/L yeast extract, 5 g/L peptone, 10 g/L glucose, and 16 g/L agar) and incubated for 24 h at 30°C. Single colonies on the YM plates were inoculated into 5 mL of yeast nitrogen base (YNB) broth (16 g/L YNB and 10 g/L glucose) and incubated at 30°C for 24 h with shaking. The cells were harvested by centrifugation at 1,878 × g for 5 min, and the supernatant was discarded. The cells were washed three times with sterile phosphate-buffered saline (Oxford, UK) and standardized to 1 × 10^6^ cells/mL in 20 mL filter-sterilized (0.20 μm polyethylene syringe filter, GVS Life Sciences, USA) RPMI-1640 medium (Sigma-Aldrich, USA). Biofilms were grown in 90 mm polystyrene Petri dishes in the presence of 500 μM AA (Sigma-Aldrich, USA), for 48 h at 37°C. After incubation, the biofilm was scraped off and filtered through pre-weighed filters (0.22 μm nitrocellulose filter, Sartorius, Germany), and the supernatant was collected in 50 mL conical tubes. The supernatants were stored at −80°C until further use. The nitrocellulose filter was dried at 37°C for 48 h and weighed to calculate the biomass.
Liquid chromatography-tandem mass spectrometry
LC-MS/MS analysis was conducted to confirm the production of PGE_2_ by C. auris. An API3200QTRAP hybrid triple quadrupole linear ion trap mass spectrometer (ABSciex, Canada) was coupled with an Agilent 1,200 SL series high-performance liquid chromatography (HPLC) system. The samples (10 μL) were injected at a flow rate of 0.5 mL/min, and the analysis was performed at 30°C. The samples were separated using a Zorbax Eclipse XDB C18, 50 × 4.6 mm column (Agilent Technologies, Germany). The mobile phases employed were solvent A and solvent B. Solvent A was a 10 mM ammonium formate aqueous solution in 5% methanol, while solvent B was a 10 mM ammonium formate aqueous solution in 95% methanol. The elution gradient consisted of 0–5 min: 50% B, 5–10 min: 50%–95% B, followed by an equilibration step, resulting in a total chromatographic run of 20 min. Atmospheric pressure electrospray ionization was performed in the negative mode. PGE_2_ reference standard (Cayman Chemicals, USA) was included in the compound optimization feature in the Analyst software to develop a multiple reaction monitoring (MRM) method with five transitions (one precursor producing five unique fragments) prior to sample separation. The ion spray voltage was set at 4,500 V, and the source temperature was maintained at 500°C. The declustering potential was 20 V, and the collision energy ranged from 14 to 30 eV for the fragment ions. The five selected transitions were 351.17/315.2, 351.17/271.2, 351.17/333.3, 351.17/189, and 351.17/235.1. Only when all five transitions were detected at the same retention time was the presence of PGE_2_ in the sample confirmed.
Quantification of PGE2 production by C. auris
Supernatants collected from the biofilms were acidified with 1M formic acid to a pH of less than 4. Lipids were extracted using solid-phase extraction (SPE) C18-T cartridges (Phenomenex, USA). The SPE cartridges were washed with 5 mL of methanol (Merck, Germany) and 5 mL of deionized water. After column preparation, 10 mL of acidified supernatant was added and allowed to flow through the column. Subsequently, 5 mL of water was used to remove the impurities. The lipids were eluted using 5 mL of ethyl acetate containing 1% methanol and collected in conical tubes wrapped in foil. The eluent was then dried under a stream of N_2_ gas, and the PGE_2_ concentration was determined using ELISA (Caymen Chemicals, USA) according to the manufacturer’s instructions. A cell-free control containing AA was included, and the background values obtained were subtracted from the experimental values. The experiment was performed in duplicate, with each sample assayed at two different dilutions, which were assayed in triplicate. The data were analyzed according to the manufacturer’s specifications, and the concentrations were normalized against the biofilm biomass (Fig. S1 and S2).
Determination of lipid profile of C. auris strains
Biofilms were prepared as described above for each strain. The cells were harvested, stored at −80°C overnight, and then freeze-dried. The cells were then suspended in a 2:1 mixture of chloroform:methanol, left overnight, filtered, and dried using a rotary evaporator. The dried filtrates (containing extracted lipids) were dissolved in chloroform, and the fatty acid was transesterified with trimethylsulfonium hydroxide (24). Fatty acid methyl esters were subsequently analyzed on a Shimadzu GC-2010 gas chromatograph with a flame ionization detector and autosampler. Injection volume was 0.5 µL, the injection port temperature was 275°C, and the split ratio was 1:10. An SGE-BPX90 column (length: 60 m, inner diameter: 0.25 mm, and film thickness 0.25 µm) was used. Hydrogen was the carrier gas at 60 cm/s linear velocity (3.19 mL/min). Initial oven temperature was 80°C, held for 1 minute, and then ramped at 10°C–280°C and held for 4 minutes. Instrument control, data collection, and analysis were performed with Shimadzu LabSolutions software. The identity of the analytes was confirmed by comparison of retention times with that of authentic standards.
Caenorhabditis elegans survival assay
The Caenorhabditis elegans glp-4; sek-1 strain was used in this study and obtained from the Caenorhabditis Genetic Center, College of Biological Science, University of Minnesota. Stocks were kept at 15°C on nematode growth medium (NGM; 2.5 g/L peptone; 15 g/L agar; and 3 g/L sodium chloride). Synchronized L4 nematodes were washed off NGM plates using M9 buffer (6 g/L Na_2_HPO_4_; 5 g/L NaCl; 3 g/L KH_2_PO_4_; and 0.25 g/L MgSO_4_) into 50 mL conical tubes and washed four times with M9 buffer. Following the wash, the nematodes were infected by spreading them on brain-heart infusion (BHI; Sigma-Aldrich) agar plates (7.8 g/L brain extract; 9.7 g/L heart extract; 2.0 g/L dextrose; 2.5 g/L disodium phosphate; and 15 g/L agar) with C. auris and C. auris mutant lawns prepared as follows: The yeast strains were inoculated into 5 mL YPD (10 g/L yeast extract, 20 g/L peptone, and 20 g/L glucose), incubated overnight with shaking at 37°C, and standardized (OD_600_ of 0.8). The standardized culture was plated onto BHI plates and incubated at 37°C for 24 h to form a lawn. The nematodes were incubated for 4 h at 25°C. After incubation, the nematodes were washed off the BHI plates with M9 buffer in 50 mL conical tubes and washed four times with M9 buffer. Approximately 60 nematodes were counted and transferred to a single well of a six-well plate containing liquid medium (20% BHI, 80% M9 buffer, and 90 μg/mL kanamycin). The nematodes were monitored for 7 days and counted each day as either alive or dead. They were considered dead if no movement was observed after mechanical stimulation (25).
Statistical analysis
C. elegans survival was assessed using the Kaplan-Meier method, and differences were determined with the log-rank test using OASIS 2 with statistical analyses performed using two-way analysis of variance with Bonferroni correction (26). All the other experiments were performed in triplicate, and the average and standard deviation were calculated unless stated otherwise. The student t-test with Bonferroni correction was carried out to determine statistically significant differences between data sets. A *P-*value of ≤0.05 was considered significant.
RESULTS
C. auris produces PGE2, and deletion of either OLE2 or POX1-3 decreases its production
The LC-MS/MS analyses confirmed the production of PGE_2_ by C. auris MRU293 from exogenous AA (Fig. 1) as all five required transitions found in the standard are also present in the extract of C. auris at similar retention times. The role of OLE2 and POX1-3 in the production of PGE_2_ by C. auris was determined by quantifying the PGE_2_ concentrations in the supernatants of deletion mutants of these genes. C. albicans SC5314 was used as a reference strain for PGE_2_ production (15). From Fig. 2, C. albicans SC5314 produced more PGE_2_ than C. auris MRU293. Interestingly, OLE2 and POX1-3 are required for production of wild-type levels of PGE_2_ by C. auris, since their mutants demonstrated decreased PGE_2_ production, which was restored to wild-type levels in the add-back strains (ole2Δ::OLE2 and pox1-3Δ::POX1-3).
Chromatogram obtained with mass spectrometry displaying five transitions MRM spectra for two samples: (A) PGE2 standard and (B) PGE2 purified from C. auris biofilm cultivated in the presence of 500 μM AA. The relevant transitions are 351.2/315.2, 351.1/271.2, 351.2/333.3, 351.2/189.0, and 351.2/235.1.
*PGE2 production by C. albicans SC5314, C. auris MRU 293, and the mutant strains, following biofilm formation in the presence of 500 μM AA, as measured by ELISA. The analysis was conducted in triplicate, with each sample assayed at two different dilutions, which were assayed in triplicate, and the reported values represent the mean with the standard deviation indicated by the error bars. Significantly different from C. auris MRU293 (ole2Δ P = 0.029; pox1-3Δ P = 0.002).
Deletion of OLE2 causes an increase in the relative percentage of monounsaturated fatty acids
From Fig. 3, it is evident that deletion of OLE2 or POX1-3 impacts fatty acid profiles of C. auris. This is especially true for OLE2, where deletion caused a shift toward an increased relative percentage of palmitoleic acid (16:1) and oleic acid (18:1) at the expense of their precursors, palmitic acid (16:0) and stearic acid (18:0). This increase was reversed in the add-back strain. In addition, no change in the relative percentage of linolenic acid (18:2) and a decrease in the relative percentage of linolenic acid (18:3) were observed (Fig. 3). The effect of deletion of POX1-3 was less clear as changes caused by the deletion mutant were not restored to wild-type levels in the add-back strains.
*Fatty acid profile of C. auris MRU 293 and the mutant strains, analyzed by gas chromatography. Values are the average of three biological repeats, and the standard deviation is indicated by the error bars. Significantly different from the wild type (P < 0.05).
OLE2 and POX1-3 are required for full virulence in C. elegans
To evaluate the role of genes implicated in PGE_2_ production in virulence, C. elegans was infected with the different C. auris strains. The results were plotted in a Kaplan-Meier graph (Fig. 4), and data were analyzed (Table 2). The results showed a significant increase in the survival rates of the nematodes infected with the deletion mutants compared with those infected with the wild-type C. auris. Additionally, the add-back strains showed similar results to the wild type, thereby indicating the restoration of the function of the deleted mutants. These findings suggest that these genes have roles in the virulence of C. auris in this infection model.
Kaplan-Meier plot indicating the survival probability to assess the virulence of C. auris strains. Escherichia coli (OP50) was used as the control.
DISCUSSION
The synthesis of the immunomodulatory eicosanoid, PGE_2_, has been established in various pathogenic Candida species, including C. albicans, C. dubliniensis, C. parapsilosis, C. glabrata, and C. tropicalis (13–16). The current work expands the conserved nature of the ability of yeasts to produce this immunomodulatory lipid molecule to C. auris. Although the metabolic pathway responsible for PGE_2_ synthesis in these yeasts has not yet been elucidated, multiple genes have been implicated in PGE_2_ synthesis (22). In this study, two genes, OLE2 and POX1-3, were investigated and found to be involved in PGE_2_ synthesis in C. auris.
OLE2 encodes a putative fatty acid desaturase that may introduce double bonds into fatty acids (27). However, the deletion of OLE2 in C. albicans did not influence fatty acid profiles of the cells, although overexpression of OLE2 did cause a slight increase in 18:1 but did not influence the levels of other PUFAs. In C. parapsilosis, the deletion of OLE2 caused an accumulation of 16:1 and 18:1, indicating that it does not encode a Δ9 desaturase (16). Similarly, we found that the deletion of OLE2 in C. auris causes an accumulation of 16:1 and 18:1 at the expense of 16:0 and 18:0, providing further indication that this gene does not encode a Δ9 desaturase. It may be speculated that Ole2 could be involved in the downstream modification or turnover of monounsaturated fatty acids. However, although no change in the relative percentage of 18:2 was observed, there was a decrease in the relative percentage of 18:3, which may also indicate that Ole2 plays a role in the activity of the Δ15 desaturase, Fad3 (Fig. 3). Future work may focus on absolute quantification of the various fatty acids to obtain clearer information regarding the role of Ole2.
Despite a similar effect on fatty acid profile between C. parapsilosis and C. auris, the effect of deletion of OLE2 on PGE_2_ synthesis in C. auris is different from C. parapsilosis and similar to that of C. albicans. The involvement of OLE2 in the production of PGE_2_ by C. albicans has been previously demonstrated as the disruption of the OLE2 gene resulted in a marked reduction in PGE_2_ production (11), but for C. parapsilosis, the deletion of the OLE2 gene showed no decrease in PGE_2_ production (16). In the case of C. auris, the deletion strain (ole2Δ) displayed a significantly reduced PGE_2_ production compared to the wild-type and add-back mutant (ole2Δ::OLE2), with the latter restoring PGE_2_ production to wild-type levels. This difference may be due to the lower percentage protein identity between Ole2 from C. parapsilosis and C. auris than between the proteins from C. albicans and C. auris (Table 3) or specific amino acid differences between the proteins.
POX1-3 encodes an acyl-CoA oxidase, which is also involved in the beta-oxidation of fatty acids by catalyzing the conversion of acyl-CoA to 2-trans-enoyl-CoA, hydrogen peroxide, and acetyl-CoA (28). Disruption of POX1-3 significantly decreased PGE_2_ production in C. parapsilosis (16, 28). This corresponds to our results since deletion of POX1-3 in C. auris also caused reduced PGE_2_ production compared to the wild type. The production was restored to wild-type levels in the add-back mutant (pox1-3Δ::POX1-3). In Table 3, it can also be seen that the percentage identity of Pox1-3 between C. auris, C. albicans, and C. parapsilosis is more than 60%, indicating that they are likely to perform similar functions in these three yeasts. Unfortunately, the increase in 16:0 and decrease in 18:1 observed for the POX1-3 deletion mutant were not restored to wild-type levels in the add-back strain, indicating potential off-target effects of the deletion.
Interestingly, although the role of OLE2 in lipid metabolism, including PGE_2_ production, seems to differ between different yeast species, the role of this gene in virulence of different pathogenic yeasts across various infection models, including human monocyte-derived macrophages (16) and nematodes, is conserved, indicating that the role of this gene in full virulence may be independent of its role in PGE_2_ production.
Deletion of POX1-3 caused a decrease in PGE_2_ production as well as virulence in both C. parapsilosis and C. auris. This implies that there is a link between the phenotypes, but it cannot be ruled out that it could be due to other metabolic consequences of this gene deletion. The addition of exogenous PGE_2_ to this infection model may untangle these two phenotypes and provide clarity regarding the mechanisms by which these two genes influence virulence. However, since the role of PGE_2_ in C. elegans and mammals is not necessarily conserved, with this eicosanoid playing a role in inflammation in mammals and in longevity in C. elegans (18), care should be taken when correlating such links in C. elegans to mammals. Nevertheless, these findings provide evidence of a conserved role of this gene in eicosanoid production, highlighting the conserved nature of PGE_2_ production in pathogenic yeasts and laying the foundation for studying this unique metabolic pathway, which differs from mammalian hosts, in search of novel antifungal drug targets.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Christie WW, Harwood JL. 2020. Oxidation of polyunsaturated fatty acids to produce lipid mediators. Essays Biochem 64:401–421. doi:10.1042/EBC 2019008232618335 PMC 7517362 · doi ↗ · pubmed ↗
- 2Mendoza SR, Zamith-Miranda D, Takács T, Gacser A, Nosanchuk JD, Guimarães AJ. 2021. Complex and controversial roles of eicosanoids in fungal pathogenesis. J Fungi (Basel) 7:254. doi:10.3390/jof 704025433800694 PMC 8065571 · doi ↗ · pubmed ↗
- 3Mochochoko BM, Pohl CH, O’Neill HG. 2023. Candida albicans-enteric viral interactions—The prostaglandin E 2 connection and host immune responses. i Science 26:105870. doi:10.1016/j.isci.2022.10587036647379 PMC 9839968 · doi ↗ · pubmed ↗
- 4Smeekens SP, van de Veerdonk FL, van der Meer JWM, Kullberg BJ, Joosten LAB, Netea MG. 2010. The Candida Th 17 response is dependent on mannan- and β-glucan-induced prostaglandin E 2. Int Immunol 22:889–895. doi:10.1093/intimm/dxq 44221059767 · doi ↗ · pubmed ↗
- 5Valdez PA, Vithayathil PJ, Janelsins BM, Shaffer AL, Williamson PR, Datta SK. 2012. Prostaglandin E 2 suppresses antifungal immunity by inhibiting interferon regulatory factor 4 function and interleukin-17 expression in T cells. Immunity 36:668–679. doi:10.1016/j.immuni.2012.02.01322464170 PMC 3334441 · doi ↗ · pubmed ↗
- 6Kalinski P. 2012. Regulation of immune responses by prostaglandin E 2. J Immunol 188:21–28. doi:10.4049/jimmunol.110102922187483 PMC 3249979 · doi ↗ · pubmed ↗
- 7Ma H, Wan S, Xia C-Q. 2016. Immunosuppressive CD 11b+Ly 6Chi monocytes in pristane-induced lupus mouse model. J Leukoc Biol 99:1121–1129. doi:10.1189/jlb.3A 0415-158R 26657791 · doi ↗ · pubmed ↗
- 8Martínez-Colón GJ, Moore BB. 2018. Prostaglandin E 2 as a regulator of immunity to pathogens. Pharmacol Ther 185:135–146. doi:10.1016/j.pharmthera.2017.12.00829274705 PMC 5898978 · doi ↗ · pubmed ↗