Antibacterial and anti-virulence effects of Saxifraga stolonifera Meeb extracts against Pseudomonas aeruginosa
Weidong Chen, Zijie Zhang, Yuanchun Huang, Lin Chen, Yijing Zhuang, Yue Li, Yuxiang Hong, Lei Liu, Qin He, Qing Peng, Fen Yao

TL;DR
This study shows that an extract from Saxifraga stolonifera can reduce the harmful effects of Pseudomonas aeruginosa by inhibiting its virulence and biofilm formation.
Contribution
The n-butyl alcohol extract of Saxifraga stolonifera is shown to have anti-virulence effects against Pseudomonas aeruginosa.
Findings
The n-butyl alcohol extract significantly downregulated virulence-related genes in P. aeruginosa.
The extract inhibited biofilm formation and pyocyanin production at sub-inhibitory concentrations.
HPLC-Q-TOF-MS/MS analysis identified over 11 compounds in the n-butyl alcohol extract.
Abstract
Saxifraga stolonifera Meeb is widely used as a traditional Chinese medicine for the treatment of infections. This study aims to evaluate the antibacterial properties and suppression of virulence by Saxifraga stolonifera Meeb extracts on Pseudomonas aeruginosa. Following extraction of Saxifraga stolonifera Meeb with petroleum ether, ethyl acetate, n-butyl alcohol, and water, the n-butyl alcohol extract had the strongest activity against P. aeruginosa PAO1 and P. aeruginosa ATCC27853, with minimum inhibitory concentration (MIC) values of 10 and 5 mg/mL, respectively. In the presence of the n-butyl alcohol (n-BuOH) extract at 1/4MIC, genes lasI, lasR, rhlI, phzA1, phzA2, and pilG were decreased to levels ranging from 13% (lasI) to 43% (phzA2). Both biofilm formation and pyocyanin production of PAO1 were inhibited by the n-BuOH extract at sub-inhibitory concentrations. N-butyl alcohol…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
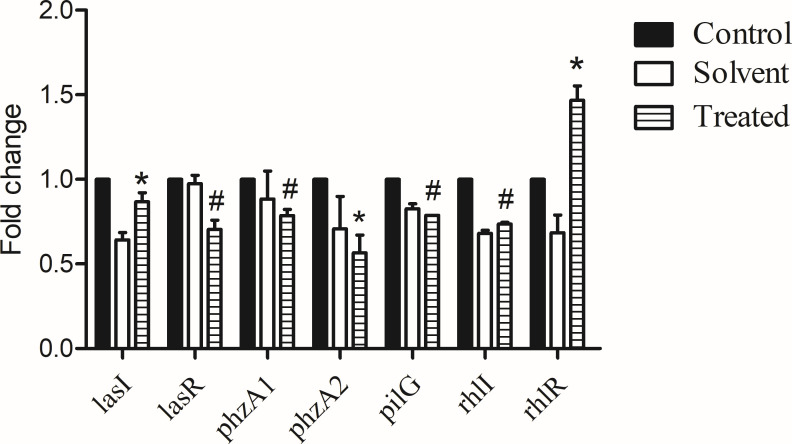 Fig 1
Fig 1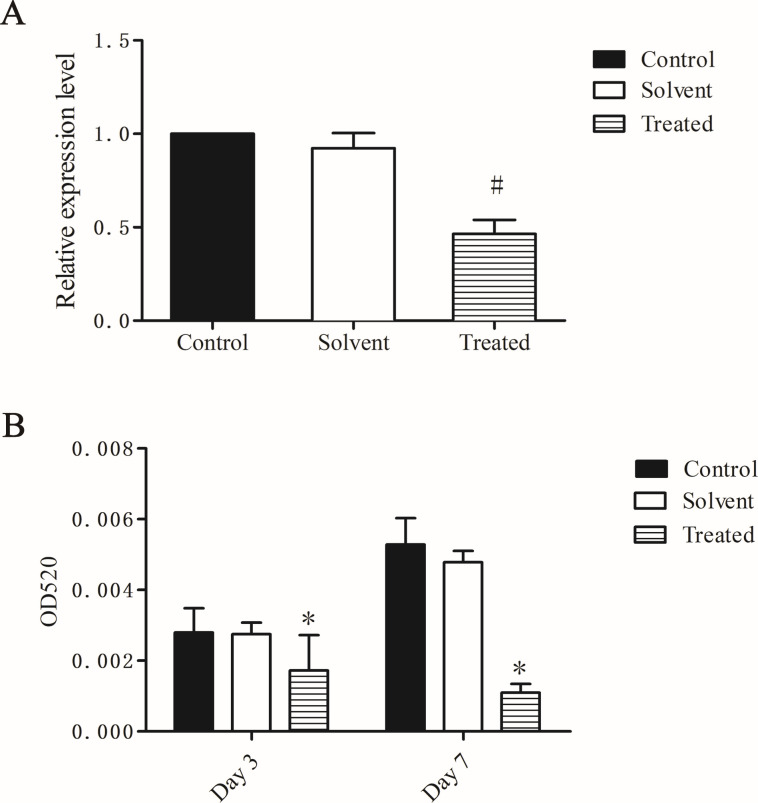 Fig 2
Fig 2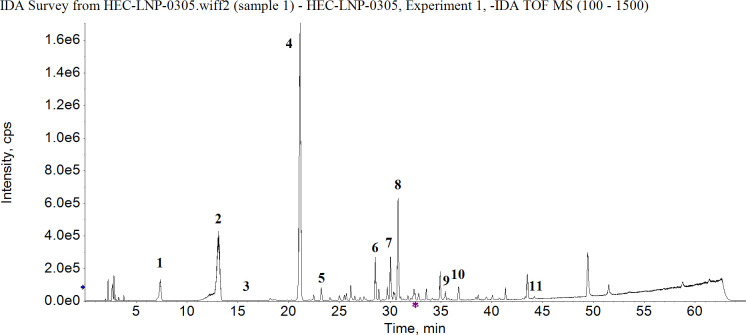 Fig 3
Fig 3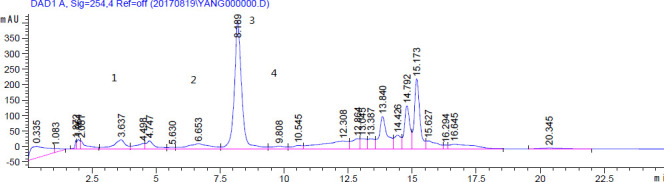 Fig 4
Fig 4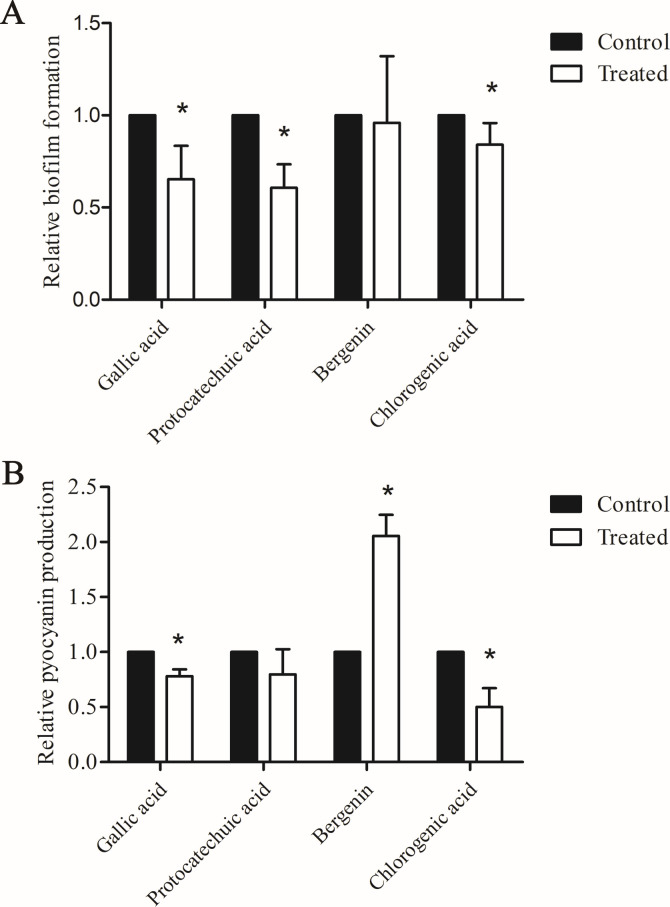 Fig 5
Fig 5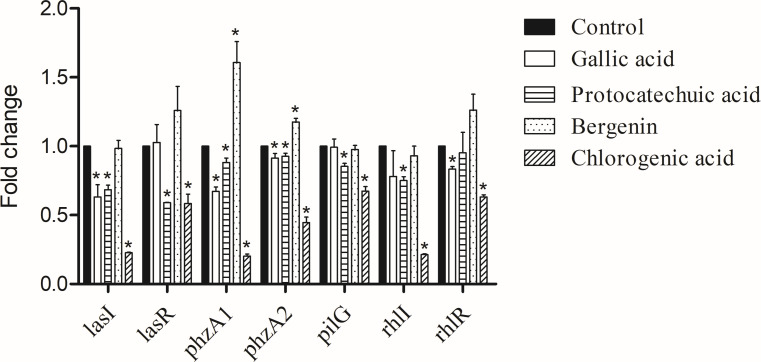 Fig 6
Fig 6| Strain | Description | Source or reference |
|---|---|---|
|
| Wild-type of | This laboratory |
|
| American Type Culture Collection | This laboratory |
|
| ( | |
|
| ( | |
|
| ( | |
|
| ( | |
|
| ( | |
|
| ( | |
|
| ( |
| MIC (mg/mL) | ||||
|---|---|---|---|---|
| Strains | n-BuOH | Ethyl acetate | Water | Petroleum ether |
| PAO1 | 10 | 12.5 | 50 | 50 |
| ATCC 27853 | 5 | 6.25 | 25 | 25 |
| Elemental compositions | [M-H]− | Major fragment ion ( | Tentative identification | ||
|---|---|---|---|---|---|
| 1 | 7.377 | C7H6O5 | 169.0137 | 169, 125, 79, 69 | Gallic acid |
| 2 | 13.089 | C13H14O9 | 313.0556 | 313, 235, 193, 165, 137 | Norbergenin |
| 3 | 15.690 | C7H6O4 | 153.0195 | 153, 109, 91, 81 | Protocatechuic acid |
| 4 | 21.149 | C14H16O9 | 327.0714 | 327, 312, 234, 192 | Bergenin |
| 5 | 23.235 | C16H18O9 | 353.0874 | 191 | Chlorogenic acid |
| 6 | 28.570 | C27H30O17 | 625.1410 | 625, 463, 300 | Quercetin 3-O-gentiobioside |
| 7 | 30.019 | C26H28O16 | 595.1291 | 595, 300 | Quercetin-3- |
| 8 | 30.765 | C21H20O12 | 463.0876 | 463, 301, 151 | Quercetin 3′-O- |
| 9 | 35.471 | C21H20O11 | 447.0938 | 447, 300, 271, 151 | Quercetin 3-O- |
| 10 | 36.736 | C15H12O7 | 303.0503 | 303, 151, 123 | Diquertin |
| 11 | 44.187 | C15H10O7 | 301.0349 | 301, 178, 151, 121, 107 | Quercetin |
| Analyte | Rt (min) | Concentration (mg/g) |
|---|---|---|
| Gallic acid | 3.7 | 6.9 ± 0.2 |
| Protocatechuic acid | 6.4 | 3.5 ± 0.1 |
| Bergenin | 8.3 | 142.3 ± 0.2 |
| Chlorogenic acid | 9.8 | 3.2 ± 0.2 |
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —Natural Science Foundation of Guangdong Provincehttp://dx.doi.org/10.13039/501100003453
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Infections and bacterial resistance · Bioactive Natural Diterpenoids Research
INTRODUCTION
Pseudomonas aeruginosa is an important gram-negative opportunistic pathogen that causes soft tissue, urinary tract, and chronic pulmonary infections (1). As an important pathogen, P. aeruginosa infection is ultimately responsible for morbidity and mortality in patients with cystic fibrosis (CF)(2, 3). Treatment has become increasingly challenging due to the dramatic increase in antibiotic resistance of P. aeruginosa (4).
As a potential approach for new antibiotics, targeting virulence factors has gained increasing attention (5). First, by decreasing the expression or activity of virulence factors, the ability of bacteria to colonize and invade the host is reduced. In addition, in contrast to traditional antibiotics, suppression of virulence does not directly kill the bacteria, leaving the bacteria with less pressure to evolve into resistant clones. Furthermore, this inhibition does not interfere with the normal microbial flora and may also allow the host immune system to prevent bacterial colonization or clear any established infection (6, 7). Quorum-sensing (QS), a process of cell-to-cell communication modulating population density-dependent group behavior and controlling the expression of virulence genes, has been extensively studied (8), and could provide new targets for screening new antimicrobial agents. There are four QS systems in P. aeruginosa, namely the las, rhl, pqs, and iqs systems (8, 9). The transcriptional regulator lasR and the cognate autoinducer synthesized by lasI constitute the las system, while rhlI and transcriptional regulator rhlR constitute the rhl system (10). Some natural anti-virulence agents that inhibit QS systems have also been identified (11, 12).
Saxifraga stolonifera Meeb is a traditional Chinese herb that can be found in most areas of southern China. It has been used for hundreds of years for the treatment of benign prostatic hyperplasia (BPH), otitis media, respiratory disease, and skin and wound infections in China (13, 14). However, activity against bacteria has yet to be confirmed. In particular, whether the extracts from S. stolonifera Meeb have anti-virulence effects against P. aeruginosa still remains unclear. In preliminary screening with selected Chinese medical herbs, we found that ethanol extracts from S. stolonifera Meeb had good activity against P. aeruginosa, with MICs ranging from 25 to 50 mg/mL. This suggests that S. stolonifera Meeb extracts might be a promising reagent used for P. aeruginosa infection and could also be used as herbal feed additives to address the increasingly severe issue of antibiotic resistance in animal husbandry (15, 16). In this study, we characterize the anti*-P*. aeruginosa effect of different fractions of an ethanol extract from S. stolonifera Meeb. Furthermore, the inhibitory effects of S. stolonifera Meeb extract on virulence gene regulatory systems, such as the QS system and virulence factors, were also investigated.
MATERIALS AND METHODS
Plant material and chemicals
Fresh leaves and stalks of S. stolonifera Meeb were collected from a rural area in Puning city, Guangdong province, China. The stalks and leaves were dried in an oven at 50℃, then ground to fine powder with a blender. Gallic acid, protocatechuic acid, bergenin, and chlorogenic acid were purchased from Shanghai Yuanye Bio-Technology Company Limited, China.
Extraction
The leaves and stems of S. stolonifera (93 g) were extracted with 95% EtOH (3 × 1 L, every 2 h, 40°C) under ultrasonication. The solution was filtered, and the collection was evaporated in vacuum to obtain a residue (9.2 g). The residue was suspended in H_2_O (0.5 L), then successively extracted at room temperature with 4 × 0.5 L of petroleum ether, EtOAc, and n-butyl alcohol (n-BuOH). Each solution was concentrated under reduced pressure (at 50°C) to result in a petroleum ether extract (1.947 g), ethyl acetate extract (1.582 g), n-BuOH (1.056 g), and water extract (4.615 g).
Determination of MICs
To compare the antibacterial activity against P. aeruginosa in vitro, MIC values were determined using broth microdilution method according to the Clinical and Laboratory Standard Institute guidelines (17). Each extract was thoroughly mixed with 20% methanol as the stock solution and diluted with sterile water. Gallic acid, protocatechuic acid, bergenin, and chlorogenic acid were dissolved in sterile water. Briefly, a 50-μL volume of each solution was dispensed into each well of a microtiter plate, followed by 50 μL Mueller-Hinton Broth containing 10^6^ CFU (colony-forming units)/mL of P. aeruginosa. The plates were incubated at 37°C for 24 h. The growth of microorganisms was observed as turbidity at 600 nm, being measured with a SpectraMax M2e (USA). The MIC was determined as the lowest concentration that completely prevented microbial growth. In addition, the MICs of ceftazidime against PAO1 and ATCC 27853 were also determined as a reference. All samples were tested in triplicate.
Bacterial strains and media
The bacterial strains used in this study are shown in Table 1. P. aeruginosa PAO1 and ATCC 27853 strains were grown and maintained in Luria-Bertani (LB) broth, Mueller-Hinton (MH) broth, or on LB or MH agar. Chromosomal fusion lux-based reporter strains were the gifts from Dr. Kangmin Duan (18).
Detection of virulence gene expression by luminescence assay
The sub-inhibitory concentration that did not inhibit the growth of PAO1 was determined by measuring the growth curve, using a SpectraMax M2e (USA). The expression of genes was determined using the lux-based reporters: the promoter regions of the gene were cloned into the upstream of the lux genes on plasmid pMS402, gene expression in liquid cultures was determined as counts per second (cps) of light production, with a Molecular Devices FilterMax F5 (USA) (18). An overnight culture of a PAO1 lux reporter was diluted 1:100 in MH broth and incubated at 37°C for 3 h. Each culture was then adjusted to OD_600_ = 0.3. In each well of a black, clear-bottom 96-well plate (Costar 3614), 10 μL PAO1 lux reporter culture was inoculated into 90 μL MH broth containing a sub-inhibitory concentration of extract/compounds. The same volume of PAO1 culture inoculated in MH broth without drug was set as the control. Luminescence was measured and the plate was shaken every 30 min for 24 h, using the microplate reader (Filter Max F5, USA). Each assay was repeated three times separately.
Biofilm formation and pyocyanin production
Quantitative analysis of biofilm formation was performed, in a 96-well polystyrene plate, according to the method of Kessler et al with some modifications (19). An overnight culture of PAO1 was diluted to OD_600_ =0.3. Then 50 μL bacterial culture was added into a 96-well plate, followed by addition of 50 μL MH culture medium, with or without n-BuOH extract and four identified compounds (gallic acid, protocatechuic acid, bergenin, and chlorogenic acid) at sub-inhibitory concentrations. The plates were incubated at 37°C as static cultures for 24 h. Following incubation, 25 μL 1% crystal violet stain was added to each well at room temperature. Fifteen minutes later, the liquid was removed, and the wells were washed four times with water. The remaining stained biofilm was dissolved in 200 μL 95% ethanol for 10 min with rocking. Then, a 150-μL aliquot was removed and transferred to another 96-well plate, and the optical density of the dissolved stain was measured at OD_600_. Microtiter wells containing medium and extract were used as blanks. Each assay was repeated three times.
The pyocyanin production assay is based on the absorbance of pyocyanin at 520 nm in acidic solution (20). PAO1 was grown in LB broth, with or without n-BuOH extract and the four identified compounds at sub-inhibitory concentrations, at 37°C for 3 days. One-milliliter supernatant of each culture was removed and then extracted with 0.6 mL of chloroform, then re-extracted with 0.2 mL of 0.2 N HCl. The absorbance of this solution was measured at 520 nm by use of the microplate reader (SpectraMax M2e, USA).
HPLC-Q-TOF-MS/MS analysis
Chromatographic separation of n-BuOH extract was performed with a Shimadzu Prominence high-performance liquid chromatography (HPLC) system using a COSMOSIL 5C18-MS-II Column (4.6 × 150 mm, 5 μm). The mobile phase consisted of two solvents: 0.1% aqueous formic acid (A) and 0.1% aqueous formic acid acetonitrile (B). The gradient program was optimized as follows: 0%–5% B at 0–10 min, 5%–50% B at 2–25 min, 50%–95% B at 25–35 min, 95% B at 35–37 min, and 95%–5% B at 37–38 min. The flow rate was set at 1 mL/min, and an aliquot of 5 μL was set as the injection volume.
Mass spectrometric determination was carried out on an X500R QTOF mass spectrometer with the ESI source in negative mode. The conditions were optimized as follows: ion spray voltage: −4,500 V, ion source temperature: 550°C, gas 1 and 2, nitrogen, 55 psi, and curtain gas, nitrogen, 35 psi. The collision energy (CE) was set at −10 V/−35 V and the declustering potential (DP) was −80 V. For the full MS-IDA (information dependent acquisition) analysis, the MS data were produced across the mass range of m/z from 100 to 1500 Da.
HPLC analysis
Samples were dissolved in methanol by ultrasound and were filtered through 0.22-μm membrane filters before analysis. The quantitative analysis of gallic acid, protocatechuic acid, bergenin, and chlorogenic acid was measured by an Agilent 1200 liquid chromatography system (Agilent Technologies, USA) equipped with a degasser, quaternary pump, an auto-sampler, and a DAD detector. An Eclipse XDB -C18 Column (5 µm, 4.6 mm × 150 mm, Agilent Technologies, USA), with a mobile phase consisting of solvent A (water containing 0.2% phosphoric acid) and solvent B (methanol), was used for separation. N-BuOH (10 µL) was injected at a 0.8 mL/min flow rate and detected at 254 nm.
Statistical analysis
All experiments were performed independently in triplicate with pooled samples of biological replicates. Data were analyzed by one-way analysis of variance with a P-value of 0.05 indicating significance.
RESULTS
MIC determination
All plant extracts showed antibacterial activity against P. aeruginosa to varying degrees (MICs from 5 to 50 mg/mL) (Table 2). The n-BuOH extract showed the strongest antibacterial effect, with MICs of 10 and 5 mg/mL against PAO1 and ATCC 27853, respectively. The water and petroleum ether fractions showed the least activity against PAO1 and ATCC 27853. In this study, the 20% methanol used to dissolve the extracts had no antibacterial effect on the MIC assay. We selected the n-BuOH extract for further experiments due to its highest activity. The MICs for gallic acid, protocatechuic acid, bergenin, and chlorogenic acid were 2, 2, 6, and 4 mg/mL, respectively, against PAO1.
N-BuOH extract differentially affects gene expression of virulence factors
We determined the maximum sub-inhibitory concentration of methanol and n-BuOH extract that did not affect the 24-h growth curve of PAO1 (data not shown). Using the extracts with the maximum sub-inhibitory concentrations, a luminescence assay was performed to determine the effect of the plant extracts on the virulence-related gene expression of PAO1. In order to exclude the influence of solvent, the effect of methanol (2.5%) was also detected. We found that methanol (2.5%) can inhibit the expression of lasI, lasR, rhlI, rhlR, phzA1, phzA2, and pilG by 36%, 3%, 32%, 32%, 12%, 30%, and 18%, respectively (Fig. 1). N-BuOH extract (containing 2.5% methanol) can inhibit the expression of lasI, lasR, rhlI, phzA1, phzA2, and pilG by 49%, 33%, 58%, 33%, 73%, and 39%, respectively. Deducting the effect of solvent, the lasI, lasR, rhlI, phzA1, phzA2, and pilG were reduced by 0.87-, 0.70-, 0.74-, 0.79-, 0.57-, and 0.79-fold in the presence of the n-BuOH extract alone (P < 0.05).
*Gene expression changes induced by n-BuOH extract of S. stolonifera Meeb, determined by luminescence assay. Relative fold change of maximum expression levels (cps) during the entire growth course of the PAO1 lux reporter strain in LB supplemented with methanol (solvent) alone (Control), or with sub-inhibitory concentrations of n-butyl alcohol extract (Treated). Control was set as 1, and the results are presented as the means ± SD of three independent experiments. P < 0.05 versus the control group. #P < 0.01 versus the control group.
N-BuOH extract shows an inhibitory effect on biofilm formation and pyocyanin production
Methanol (2.5%) had no effect on biofilm formation and pyocyanin production (Fig. 2). N-BuOH extract at sub-inhibitory growth concentrations can inhibit biofilm formation by 46% compared with the control group. On days 3 and 7, the production of pyocyanin was reduced by 39% and 79%, respectively, with sub-inhibitory concentrations of n-BuOH extract (Fig. 3), which suggests that the n-BuOH extract can suppress the pathogenesis related to pyocyanin production.
*Biofilm formation (A) and pyocyanin production (B) affected by n-BuOH extract of S. stolonifera Meeb. The results are presented as the means ± SD of three independent experiments. P < 0.05 versus the control group. #P < 0.01 versus the control group.
The total ion chromatogram (TIC) for the n-BuOH extract of S. stolonifera (L.) Meeb. gallic acid (peak 1), norbergenin (peak 2), protocatechuic acid (peak 3), bengenin (peak 4), chlorogenic acid (peak 5), quercetin 3-O-gentiobioside (peak 6), quercetin-3-O-β-D-xylopyranosyl-(1→2)-β-D-galactopyranoside (peak 7), quercetin 3′-O- β-D-galactopyranoside (peak 8), quercetin 3-O- β-D-rhamnoside (peak 9), diquertin (peak 10), and quercetin (peak 11).
Flavonoids and phenolic acids are major components in n-BuOH extract of S. stolonifera Meeb
Eleven compounds were detected in n-BuOH extract. Six of them belonged to flavonoids, including quercetin and derivatives of quercetin (Table 3). Phenolic acids, such as norbergenin, bergenin, gallic acid, and protocatechuic acid, were also found.
It has been known that the main components of ethanol extracts from S. stolonifera Meeb contain bergenin, quercetin, protocatechuic acid, gallic acid, and β-sitosterol (21). Based on the HPLC-Q-TOF-MS/MS data, we therefore quantified the contents of gallic acid, protocatechuic acid, bergenin, chlorogenic acid, and quercetin (other chemicals listed in Table 3 were not determined by HPLC because standard samples were not commercially available). The total ion chromatogram (TIC) for the n-BuOH extract is illustrated in Fig. 3, with four compounds, i.e., gallic acid, protocatechuic acid, bergenin, and chlorogenic acid, being clearly identified (Fig. 4). Bergenin was the major chemical constituent in the dry extract, with a chemical content of 142.3 ± 0.2 mg/g, and the chemical contents of gallic acid, protocatechuic acid, and chlorogenic acid being 6.9 ± 0.2, 3.5 ± 0.1, and 3.2 ± 0.2 mg/g, respectively (Table 4). Quercetin was not detectable by HPLC, which suggests the low content of quercetin in the n-BuOH extract of S. stolonifera Meeb.
Chromatograms of the identified compounds in mixed standard solutions, gallic acid (peak 1), protocatechuic acid (peak 2), bergenin (peak 3), and chlorogenic acid (peak 4)
Effects of gallic acid, protocatechuic acid, bergenin, and chlorogenic acid on biofilm formation, pyocyanin production, and virulence factor gene expression
The effects of the four identified compounds at maximum SIC on biofilm formation and pyocyanin production are shown in Fig. 5. Protocatechuic acid, gallic acid, and chlorogenic acid reduced biofilm formation by 40%, 35% and 16% (P < 0.05). Gallic acid, protocatechuic acid, and chlorogenic acid reduced the production of pyocyanin by 22%, 20%, and 50% (P < 0.05). Bergenin had no effect on biofilm formation and promoted the production of pyocyanin.
*Biofilm formation (A) and pyocyanin production (B) affected by gallic acid, protocatechuic acid, bergenin, and chlorogenic acid. The results are presented as the means ± SD of three independent experiments. P < 0.05 versus the control group.
The effects of the four identified compounds at maximum SIC on gene expression of virulence factors are shown in Fig. 6. Gallic acid inhibited the expression of lasI, phzA1, phzA2, and rhlR by 37%, 33%, 9%, and 22%, respectively (P < 0.05). Protocatechuic acid inhibited the expression of lasI, lasR, phzA1, phzA2, pilG, rhlI by 32%, 41%, 12%, 8%, 15%, and 25%, respectively (P < 0.05). Chlorogenic acid inhibited the expression of lasI, lasR, phzA1, phzA2, pilG, rhlI, and rhlR by 78%, 42%, 80%, 55%, 33%, 79%, and 27%, respectively (P < 0.05). To our surprise, bergenin increased the expression of phzA1 and phzA2 by 60% and 18%, respectively (P < 0.05).
*Effects of gallic acid, protocatechuic acid, bergenin, and chlorogenic acid on the indicated virulence factor gene expression. P < 0.05 versus the control group.
DISCUSSION
The final products of the genes characterized in this study are important virulence factors that are involved in quorum sensing, motility, and bacteria-host interactions. The QS system regulates the formation of biofilm and the expression of many virulence factors in P. aeruginosa (22).
In order to determine whether the decreased expression of the tested genes results in the phenotypic changes of the PAO1 strain, we characterized the effects of the n-BuOH extract on production of pyocyanin and biofilm formation. Most of the antibiotics used to treat P. aeruginosa infections must cross a biofilm, which represents a protective mode of growth, since biofilm as a barrier blocks the ability of a variety of antibiotics to reach the bacteria, causing multi-drug resistance (23, 24). Biofilm formation is also closely correlated with infection and is involved in establishing chronic infections that are partly regulated by both the las and rhl systems (25, 26). Therefore, inhibiting biofilm formation or the ability to remove a biofilm would be helpful for the treatment of P. aeruginosa infections. In the present study, the n-BuOH extract of Saxifraga stolonifera Meeb suppressed the expression of the lasI, lasR, and rhlI genes, whereas rhlR was unchanged. The las and rhl systems are important components of the QS network and co-regulate the biofilm formation of P. aeruginosa (9). The las system plays an important role in biofilm formation because it is considered to be the top of the QS network (27). LasR, as the transcriptional regulator of the las system, not only regulates the las system, but also influences the expression of the rhl and pqs systems (28). The rhlI gene favors biofilm formation in P. aeruginosa by enhancing the production of matrix exopolysaccharide through the pel operon, and biofilm formation in lasI and rhlI mutants is severely reduced (29). In our luminescence assay experiments, we also showed that the expression of pilG is suppressed by the n-BuOH extract. PilG is one of the factors that influences the twitching motility of P. aeruginosa (30). Twitching motility is required for biofilm development (31). Thus, the downregulation of pilG, lasR, lasI, and rhlI caused by our n-BuOH extract may be the reason for the reduced biofilm formation.
Pyocyanin is one of the major virulence factors in P. aeruginosa and contributes to both acute and chronic infection (32, 33). It is also one of the major factors that suppresses lymphocyte proliferation. Pyocyanin can also exacerbate P. aeruginosa infections by promoting the formation of biofilms and delaying recovery of burn wounds (34). Pyocyanin production is regulated by the QS system. RhlR upregulates the synthesis of pyocyanin and rhamnolipid (35). The two homologous operons, phzA1 and phzA2, are involved in the synthesis of phenazine, a pyocyanin precursor (34). We show that when P. aeruginosa grows in the presence of 2.5 mg/mL n-BuOH extract, the production of pyocyanin is reduced by 0.40- and 0.89-fold on the 3rd and 7th days in Fig. 3, which suggests that the n-BuOH extract can suppress the pathogenesis related to pyocyanin production, and the expression of phzA1 and phzA2 is both inhibited. Therefore, it is possible that the reduced production of pyocyanin might be due to inhibition of rhlR, phzA1, and phzA2 expression.
Flavonoids and phenolics in n-BuOH extract are responsible for both antibacterial and anti-virulence effects. Although bergenin accounted for more than 14% of the total content of n-BuOH extract, in this study, bergenin alone had no antibacterial or anti-QS effects against standard P. aeruginosa strains at concentrations up to 3 mg/mL. Flavonoids from multiple plants have been reported to be effective in inhibiting the growth of P. aeruginosa (36, 37). Galvão et al. proposed that the antibacterial mechanism of flavonoids is related to their binding to the P. aeruginosa cell wall (36). As the n-BuOH extract is rich in flavonoids, we therefore consider that the flavonoids contribute to the anti-P. aeruginosa effect of n-BuOH extract from S. stolonifera Meeb. Flavonoids (including quercetin, baicalein, naringenin, and kaempferol) and flavonoid-rich fractions from some herbs have also been reported to have anti-quorum sensing and anti-virulence effects against P. aeruginosa (38–40). Paczkowski et al. showed that flavonoids inhibit P. aeruginosa QS systems through antagonism lasR and rhlR (41). These studies suggest that flavonoids might also play key roles in the anti-quorum sensing effect of n-BuOH extract from S. stolonifera Meeb.
Another major group of compounds from Saxifraga stolonifera Meeb is phenolic acids. Several studies demonstrate that phenolic acids are responsible for the antibacterial activity of some plants against P. aeruginosa (42, 43). The mechanism of the anti-P. aeruginosa effect of phenolic acids is related to their ability to damage the cell wall and cytoplasmic membrane (44). It may play an important role in inhibiting the virulence expression of foodborne bacteria and their prevention and control (45). Phenolic acids can inhibit the QS system of P. aeruginosa isolates, thereby influencing the expression of virulence factors, for example, by reducing pyocyanin production and inhibiting biofilm formation and swarming motility (28, 42). Thus, phenolic acids might partly explain the antibacterial and anti-virulence effects of n-BuOH extract from S. stolonifera Meeb.
Conclusion
Our results demonstrate that n-BuOH extract of S. stolonifera Meeb has both antibacterial and anti-virulence effects on P. aeruginosa. N-BuOH extracts of S. stolonifera Meeb may be utilized in the future as an antibacterial and anti-virulence agent for the treatment of P. aeruginosa.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Reynolds D, Kollef M. 2021. The epidemiology and pathogenesis and treatment of Pseudomonas aeruginosa infections: an update. Drugs (Abingdon Engl) 81:2117–2131. doi:10.1007/s 40265-021-01635-6PMC 857214534743315 · doi ↗ · pubmed ↗
- 2Weimann A, Dinan AM, Ruis C, Bernut A, Pont S, Brown K, Ryan J, Santos L, Ellison L, Ukor E, et al.. 2024. Evolution and host-specific adaptation of Pseudomonas aeruginosa. Science 385:eadi 0908. doi:10.1126/science.adi 090838963857 PMC 7618370 · doi ↗ · pubmed ↗
- 3Rossi E, La Rosa R, Bartell JA, Marvig RL, Haagensen JAJ, Sommer LM, Molin S, Johansen HK. 2021. Pseudomonas aeruginosa adaptation and evolution in patients with cystic fibrosis. Nat Rev Microbiol 19:331–342. doi:10.1038/s 41579-020-00477-533214718 · doi ↗ · pubmed ↗
- 4Botelho J, Grosso F, Peixe L. 2019. Antibiotic resistance in Pseudomonas aeruginosa – mechanisms, epidemiology and evolution. Drug Resist Updat 44:100640. doi:10.1016/j.drup.2019.07.00231492517 · doi ↗ · pubmed ↗
- 5Hibbert TM, Whiteley M, Renshaw SA, Neill DR, Fothergill JL. 2024. Emerging strategies to target virulence in Pseudomonas aeruginosa respiratory infections. Crit Rev Microbiol 50:1037–1052. doi:10.1080/1040841 X.2023.228599537999716 · doi ↗ · pubmed ↗
- 6Scoffone VC, Barbieri G, Irudal S, Trespidi G, Buroni S. 2024. New antimicrobial strategies to treat multi-drug resistant infections caused by gram-negatives in cystic fibrosis. Antibiotics (Basel) 13:71. doi:10.3390/antibiotics 1301007138247630 PMC 10812592 · doi ↗ · pubmed ↗
- 7Liao C, Huang X, Wang Q, Yao D, Lu W. 2022. Virulence factors of Pseudomonas aeruginosa and antivirulence strategies to combat its drug resistance. Front Cell Infect Microbiol 12:926758. doi:10.3389/fcimb.2022.92675835873152 PMC 9299443 · doi ↗ · pubmed ↗
- 8Rodríguez-Urretavizcaya B, Vilaplana L, Marco MP. 2024. Strategies for quorum sensing inhibition as a tool for controlling Pseudomonas aeruginosa infections. Int J Antimicrob Agents 64:107323. doi:10.1016/j.ijantimicag.2024.10732339242051 · doi ↗ · pubmed ↗