C. elegans as a tractable infection model for the emerging fungal pathogen Candida auris
Melissa Martinez, Melissa R. Cruz, Danielle A. Garsin, Michael C. Lorenz

TL;DR
Researchers developed a C. elegans model to study the emerging fungal pathogen Candida auris, showing it can detect virulence differences and support high-throughput screening.
Contribution
A novel C. elegans infection model for Candida auris that enables high-throughput screening and strain-specific virulence analysis.
Findings
C. elegans exposed to C. auris shows lethal effects with kinetics similar to Candida albicans.
An adenine auxotrophic mutant (ade2∆) of C. auris is avirulent in C. elegans.
The model can detect strain-to-strain virulence differences and supports high-throughput assays using SYTOX Orange.
Abstract
Candida auris is an emerging multidrug-resistant fungal pathogen. The genetic factors contributing to the virulence, drug resistance, and stress-tolerant nature of C. auris are mostly unknown. Additional animal models of virulence are needed, especially those amenable to high-throughput analysis. The nematode Caenorhabditis elegans has been validated as an effective tool for studying multiple fungal and bacterial pathogens. We describe here a C. elegans infection model in which exposure to C. auris is lethal to worms with kinetics similar to killing by Candida albicans; in contrast to C. albicans, C. auris does not form hyphae, indicating distinct virulence mechanisms. Furthermore, an engineered mutant auxotrophic for adenine biosynthesis (ade2∆) is avirulent, as it is in many models, indicating that the nematode can discriminate between virulent and avirulent mutants. Moreover, the C.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
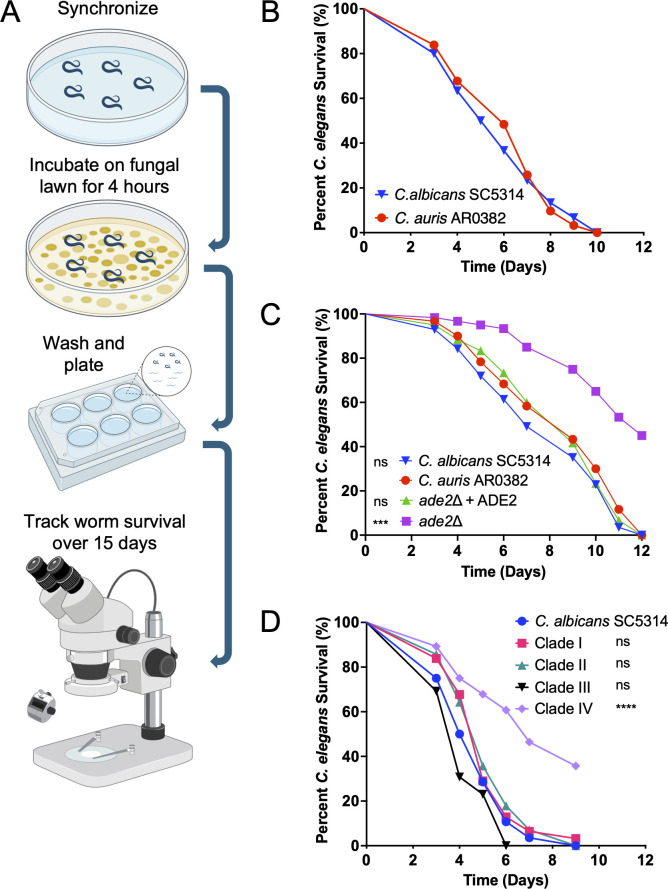 Fig 1
Fig 1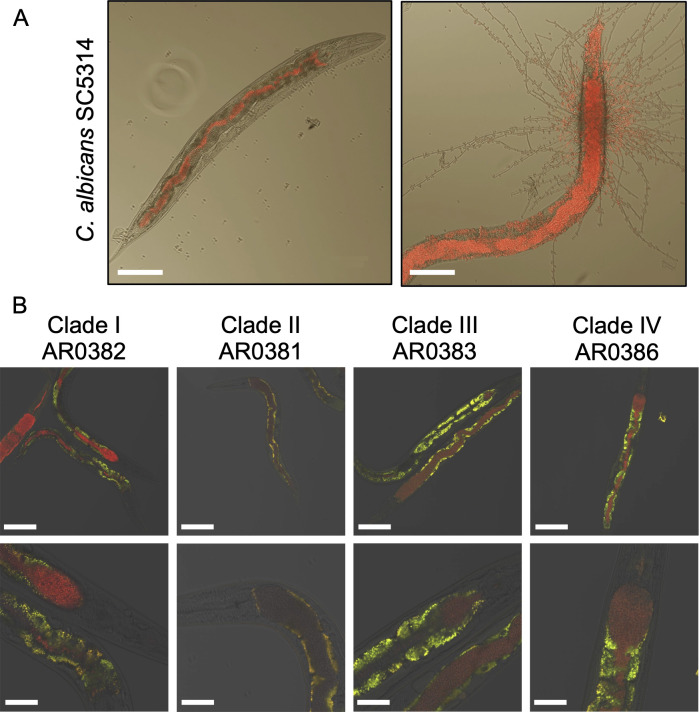 Fig 2
Fig 2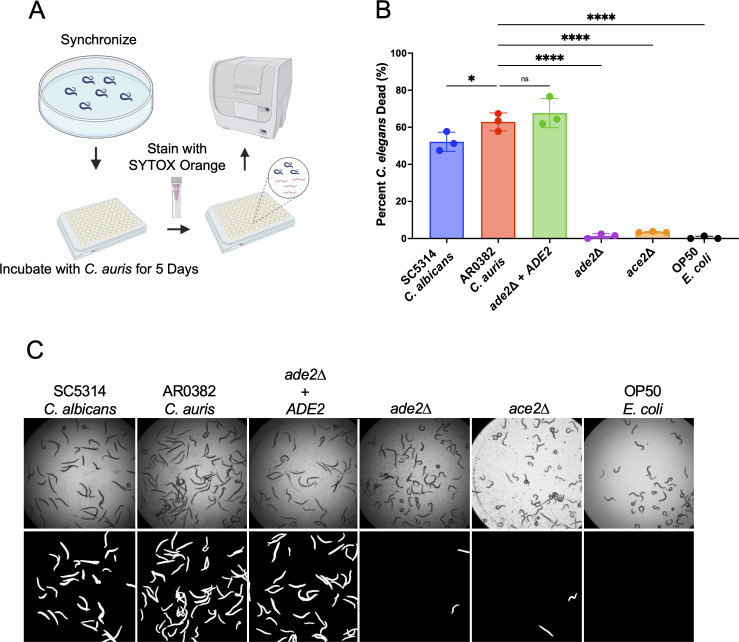 Fig 3
Fig 3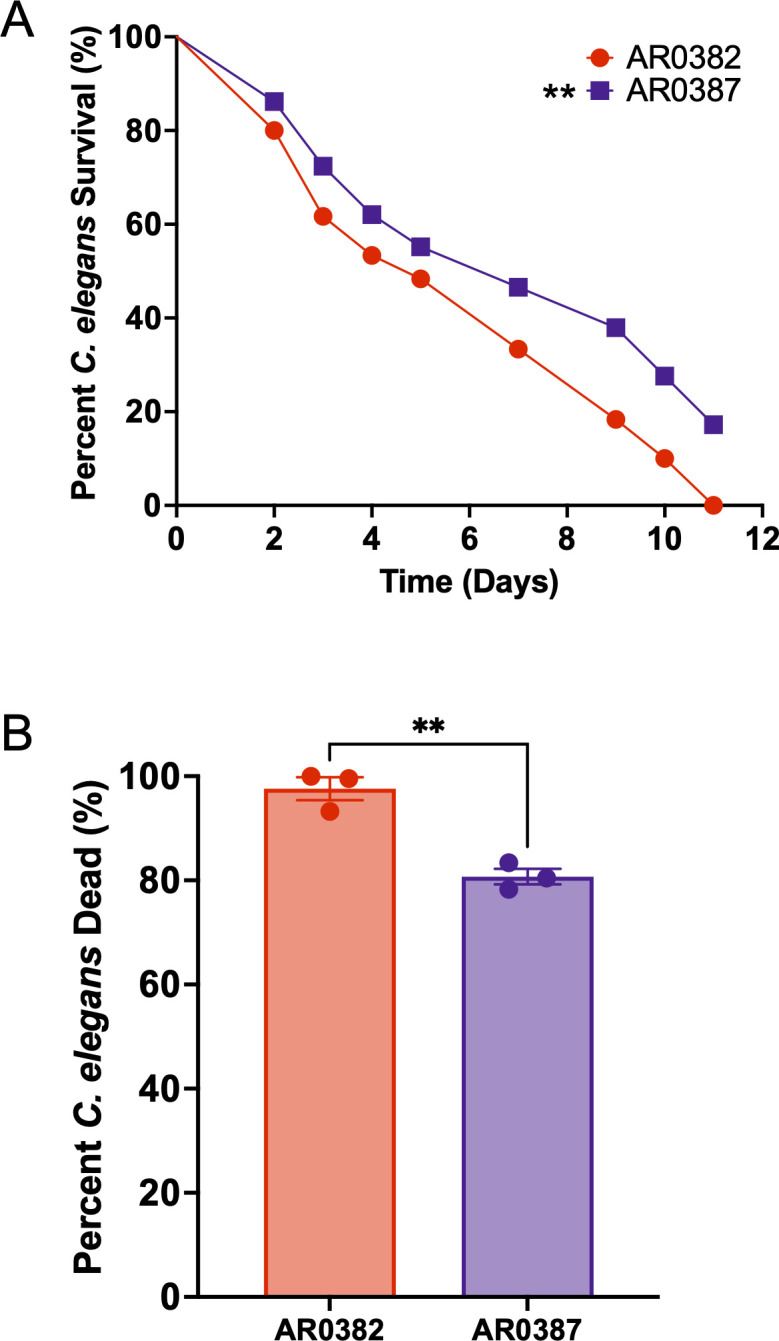 Fig 4
Fig 4- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Genetics, Aging, and Longevity in Model Organisms · Legionella and Acanthamoeba research
INTRODUCTION
Candida (Candidozyma) auris is an emergent fungal pathogen, first described in early 2009, that has rapidly risen in incidence as a highly transmissible hospital-acquired infection that primarily afflicts those who are immunocompromised (1–3). C. auris is particularly concerning due to its multidrug resistance, with about 90% of isolates resistant to some azole antifungals, 30% to amphotericin B, and 5%–10% to echinocandins (CDC). C. auris is particularly difficult to eradicate from healthcare facilities because it withstands standard disinfection protocols and persists on both abiotic surfaces and skin better than other fungal pathogens (4–7). The mortality rate for immunocompromised patients with an invasive C. auris infection can be as high as 60%, while having a chronic illness alongside the infection can increase mortality to rates as high as 80% (8–15). Initially identified in Japan, four distinct genomic clades rapidly arose that were initially geographically concentrated in East Asia, South Asia, Southern Africa, and South America (1, 16). There are now potentially six clades of C. auris (including isolates from Iran and Thailand) with several circulating in the United States and Western Europe, thanks to global travel (17–23). Because of its growing incidence and widespread drug resistance, the World Health Organization has classified C. auris as a critical fungal pathogen threat.
Most of what we understand about C. auris is inferred mainly from its more well-characterized but distant relative, Candida albicans (24–27). This distance is reflected in the recent reclassification of C. auris as Candidozyma auris, though C. albicans and C. auris are both part of the monophyletic CTG clade that includes several fungal pathogens. These fungi share conserved virulence factors, such as adhesins, proteases, and some genetic mutations that result in drug resistance (26, 28–32). However, significant differences distinguish C. auris from other Candida species. Most notably, while C. albicans is a dimorphic fungus, C. auris exists almost exclusively as a yeast, occasionally forming pseudohyphae under specific conditions (33–35). Indeed, the multidrug resistance common to C. auris is rare in other Candida species (26, 36–38). There is substantial phenotypic variation that arises from the genomic diversity both within and between clades (16, 37–41). This includes virulence in animal models, cellular aggregation, persistence on the skin, and drug resistance (16, 23, 42–45). The genetic factors that give rise to this diversity are largely unknown. Patient risk factors for developing C. auris infections overlap with other common nosocomial pathogens, but the persistence on the skin and abiotic surfaces and the resistance to some hospital disinfectant protocols facilitate person-to-person transmission that is rare among fungal pathogens (5, 46). We currently lack even a rudimentary understanding of the molecular basis of these enhanced challenges.
Studies using vertebrate and invertebrate animal models, including mice, Galleria mellonella, Drosophila melanogaster, and zebrafish, have shown that virulence and infectivity vary both between and within clades (41, 42, 47–51). Relative to well-established murine models for C. albicans, establishing a C. auris infection in mice requires either significant immunosuppressive regimes, a higher inoculum (100 times or more than that of C. albicans), or both (41, 52–55). Additional animal models, especially those amenable to high-throughput studies, are needed to bridge the knowledge gaps in C. auris virulence.
The nematode C. elegans has been established as a model for studying many human bacterial and fungal pathogens like Pseudomonas aeruginosa, Salmonella enterica, Enterococcus faecalis, Cryptococcus neoformans, and Candida albicans. Our group has used the nematode extensively to characterize aspects of E. faecalis pathogenesis and the antifungal properties of peptides derived from an E. faecalis bacteriocin, including initial findings with C. auris (56–59).
In this work, we sought to further characterize and expand the utility of C. elegans as an infection model for C. auris. We found that C. auris can establish a tractable infection in the worm with similar kinetics to that of C. albicans. This model can distinguish between a wild-type strain and engineered mutants predicted to be avirulent. It is also amenable to a medium-throughput 96-well plate-based assay using a live/dead stain and automated image analysis. The ease of inoculation via feeding, low maintenance cost, and high-throughput capability make the nematode an appealing model for studying C. auris, a genetically diverse and largely uncharacterized organism. Our findings suggest many opportunities for applying this model more widely as a screening tool to understand the unique virulence traits of this concerning pathogen.
RESULTS
C. auris infects and kills the nematode with kinetics similar to C. albicans
We know from previous work that C. albicans can colonize the nematode intestine, subsequently undergoing hyphal morphogenesis and tissue invasion that ultimately results in the animal’s death (56, 59–61). In this study, we sought to determine whether C. auris could similarly establish an infection. We used C. auris clinical isolate AR0382 (clade I) from the Antimicrobial Resistance Bank at the Centers for Disease Control and Prevention. We chose this strain because it was previously used in several studies, is virulent in several animal models, and is amenable to genetic modification (62–64). For comparison, we used C. albicans SC5314, a well-characterized laboratory strain for which a C. elegans infection model was previously developed (60, 61). We used a similar infection protocol that involved feeding on lawns of C. auris for 4 hours (Fig. 1A), followed by washing and transfer into liquid medium. C. elegans survival was then tracked daily, with dead worms being removed and recorded. The kinetics of nematode survival were similar to C. albicans; worms began to die by 4 days post-infection, and the median lethal time (LT_50_) of both strains was between 5 and 6 days (Fig. 1B).
C. auris can establish an infection in the nematode with kinetics similar to C. albicans. (A) Schematic of the canonical nematode survival assay. Worms are synchronized to the L4 stage, then incubated on a fungal lawn for 4 hours at 25°C. After incubation, the worms are washed and transferred to liquid medium in a polystyrene plate, where survival is monitored daily. (B) C. auris AR0382 and C. albicans SC5314 kill C. elegans with similar kinetics, with all animals succumbing to infection within 10–12 days. (C) In the nematode model, the C. auris ade2∆ mutant demonstrates significantly reduced virulence compared to both the parent and complemented strains. The statistical comparisons indicated are to AR0382. ∗∗∗∗P < 0.0001; ns, not significant. (D) The nematode model can distinguish between C. auris isolates of varying virulence. The isolate from clade IV is significantly attenuated compared to the other strains tested.
A key feature of any infection model is its ability to distinguish between strains with different virulence profiles. To test whether C. elegans can differentiate between strains of C. auris with varying levels of virulence, we created an ade2∆ mutant in the AR0832 background using CRISPR/Cas9. The ADE2 gene encodes the enzyme phosphoribosylaminoimadazole carboxylase, a key enzyme of adenine biosynthesis; purine metabolism is required for virulence in essentially all pathogens in which it has been tested, including both ade2∆ and ura3∆ mutants in C. albicans and C. neoformans (65–68). As expected, the ade2Δ strain does not grow in media lacking adenosine or adenine (not shown). It shows a modest growth defect compared to the parent strain (Fig. S1A) when grown in nutrient-poor worm medium (80% M9/20% brain-heart infusion [BHI]; see Materials and Methods). There is no growth defect in nutrient-replete yeast extract-peptone-dextrose (YPD) medium (Fig. S1B). We found that the LT_50_ of the mutant was 10 days, compared to approximately 6 days for the parent strain (Fig. 1C). This is a significantly greater attenuation in the worm model than seen for the highly avirulent C. albicans cph1∆ efg1∆ mutant (60, 61). The avirulence of the ade2∆ mutant in C. elegans confirms that this model can elucidate virulence differences between genetic variants in otherwise isogenic strains.
We further wanted to explore whether C. elegans can distinguish between different isolates of C. auris. We tested one isolate from each of the four major clades. In this assay, the isolate from clade IV was less virulent than isolates from the other three clades, which behaved similarly to the SC5314 strain. The data suggest that C. elegans can distinguish different levels of virulence between C. auris strains (Fig. 1D). This may represent inherent differences between clades in this model but is perhaps more likely to recapitulate strain-to-strain variation in virulence seen in other models, including in mice (39, 40, 49, 51, 52, 63, 64, 69).
C. auris colonizes the intestine of C. elegans but does not form hyphae
In previous studies, C. albicans was observed to establish an infection in C. elegans by colonizing the lumen of the intestine. Later in the infection, typically around 5–6 days post-inoculation, C. albicans begins to form hyphae, which penetrate the worm from the inside out (56, 60, 61, 70) and result in worm death, as the hyphae and/or pseudohyphae damage the intestinal wall and allow the fungus to invade surrounding tissues (Fig. 2A) (56, 60, 61). Like C. albicans, C. auris colonized the lumen of the worm intestine, distending the normal gut architecture (Fig. 2B). Though there is some evidence that C. auris can form pseudohyphae under certain conditions, we did not observe this occurring in the C. elegans intestine. Therefore, C. auris virulence in the worm appears to use mechanisms unrelated to tissue invasion, indicating significant differences relative to C. albicans. What these might be remains unknown, but C. auris has, for instance, a distinct set of novel adhesins like Scf1, which may contribute to colonization in this model (64). There were no qualitative differences in gut colonization across strains from each of the four major clades of C. auris.
C. auris colonizes the worm gut without penetrating intestinal tissue. (A) C. albicans first colonizes the intestine, causing distention, followed by hyphal morphogenesis, which results in tissue invasion. Scale bars are 100 μm. (B) Nematodes infected with clinical isolates from the four major C. auris clades. The bottom row shows magnified views of the distended intestine. Candida cells are red from a fluorescent marker, and the intestinal cells are yellow/green due to autofluorescence. Scale bars: 120 μm (top row) and 40 micrometers (bottom row).
A C. elegans high-throughput assay to study C. auris infection
Recent studies have utilized the nematode model in high-throughput assays, primarily focused on discovering novel antimicrobial agents (71–77). We wanted to determine whether the C. elegans model could be adapted to a high-throughput approach to investigate C. auris virulence. Rather than inoculation by feeding on a fungal lawn, a fungal inoculum is added to C. elegans in liquid medium, followed by a 5-day coincubation (Fig. 3A). The worms are then washed and stained overnight with SYTOX Orange, a nucleic acid-binding dye that stains dead worms for an easily visualized live/dead assay. The dead-to-total worms ratio is calculated using an automatic image analysis pipeline adapted from Conery et al. and used as a proxy for virulence (72). In this context, we sought to assess the virulence of the ade2Δ mutant and an ace2Δ mutant created by the O'Meara lab that had attenuated virulence in the G. mellonella model (64, 78). We found that the high-throughput assay recapitulated the results from the standard C. elegans survival assay, with the ade2Δ and ace2Δ mutants associated with significantly reduced worm death relative to the parent AR0382 strain (Fig. 1C and 3B). Qualitative data could also be derived from the images used in the pipeline. Representative bright-field and fluorescent images, processed via the CellProfiler pipeline (72), are shown in Fig. 3C. The virulence patterns of these mutants were similar to those observed with the OP50 Escherichia coli strain widely used to propagate C. elegans in the lab (Fig. 3C).
Results from canonical C. elegans survival experiments are recapitulated using a high-throughput assay. (A) Schematic of the high-throughput SYTOX Orange assay. Worms are synchronized, incubated with C. auris culture for 5 days, washed, stained with SYTOX Orange, and imaged using a plate reader. (B) The C. auris ade2∆ mutant shows reduced virulence compared to both the parent and complemented strains. ∗P < 0.05; ∗∗∗∗P < 0.0001; ns, not significant. (C) Representative images from the high-throughput SYTOX Orange assay. The top row shows pictures taken in bright field, and the bottom row contains fluorescent images processed by the CellProfiler analysis pipeline, as described (72), to normalize for image intensity and background.
Virulence phenotypes differentiating C. auris strains are recapitulated in the C. elegans model
We aimed to assess whether our model could replicate the phenotypes observed in mammalian models, particularly in mice. Previously published studies using mice showed that AR0382 is more virulent than AR0387, another clade I isolate. Santana et al. report that mice infected with AR0382 succumbed within 15 days post-inoculation, while 50% of those infected with AR0387 survived beyond 20 days (64). Additionally, the fungal burden observed in mice differed between the two isolates; Vila et al. reported higher CFU counts in the kidneys of mice inoculated with AR0382 (48). When tested in C. elegans as shown in Fig. 4, worms infected with AR0382 succumbed more rapidly to infection, with an LT_50_ of 5 days, compared to 7 days for those infected with AR0387 (Fig. 4A). This difference in comparative virulence was also reflected in the SYTOX assay, where AR0382 was associated with significantly greater nematode death (Fig. 4B). These findings demonstrate that AR0382 is more infectious than AR0387 in the C. elegans model, which agrees with the mouse model (18, 30, 48, 64). These data further support C. elegans as a relevant model for C. auris infection and one with substantial experimental advantages.
AR0387 is less virulent than AR0382 in the C. elegans model. (A) In the standard assay, C. elegans survival over time following exposure to C. auris AR0387 compared to AR0382. ∗∗P < 0.01. (B) Percent C. elegans dead following 5 days of exposure to AR0387 and AR0382 and assayed using the SYTOX Orange assay. ∗∗P < 0.01.
DISCUSSION
C. auris is a greatly concerning clinical problem that presents many mysteries. Its near-simultaneous emergence on multiple continents in at least four (and perhaps six) genetically distinct clades defies an easy explanation. While hypotheses have been mooted for its environmental reservoirs, drug resistance, and sudden emergence, none of these are backed by compelling evidence (79–82). Nevertheless, clinical cases are increasing exponentially in the United States and across the world (22, 23, 83–85). Containment and treatment of this emerging pathogen requires better experimental models to understand the disease potential of different strains and clades and the underlying genetic features that define them. We show here that C. elegans can be used as a C. auris infection model. One earlier study using C. elegans evaluated virulence differences among clinical isolates exclusively with a 5-day survival assay (47). Here, we expand on this approach by employing both an extended survival assay and a shortened, high-throughput-compatible assay. By tracking survival over time, we demonstrated that C. auris can establish an infection in the nematode with kinetics similar to those of C. albicans. In both species, this begins with gut colonization, but C. auris infection proceeds without the tissue penetration observed with C. albicans. A SYTOX-based live/dead fluorescence assay successfully recapitulated these results, providing the basis of high-throughput screening.
Importantly, a mutant in ADE2, known to strongly affect virulence in multiple microbial pathogens, was attenuated compared to the parent strain in both the survival and SYTOX assays. Deleting the ADE2 gene in other pathogenic fungi like C. albicans and C. neoformans species was associated with dramatically reduced virulence (67, 68). The C. auris ade2∆ null mutant in our model was significantly less virulent than the parent strain, as expected. The C. elegans model also recapitulated strain variation in phenotypes observed in murine studies, with clinical isolate AR0382 being considerably more virulent than AR0387.
We suggest that C. elegans has an important role as a screening model for in vivo study of new and rapidly evolving pathogens like C. auris, complementing more complex and costly vertebrate models. Its advantages include the ability to propagate large numbers of isogenic animals quickly and cheaply in the laboratory environment, thanks to its short generation time (3 days), well-established genetics, and its successful use with many bacterial and fungal pathogens (86). The model can be used in a standard plate-based nematode survival assay, but we also showed that it is amenable to a high-throughput methodology using a SYTOX-based live/dead assay to test C. auris virulence. Furthermore, C. elegans provides a platform for in vivo imaging of microbial infections, offering valuable insights into the host-pathogen interaction, highlighted by the fact that C. auris does not invade the worm tissue, unlike C. albicans. This reinforces that the virulence mechanisms of these two distant relatives are likely to be quite different, and that C. albicans paradigms cannot be extrapolated to C. auris without direct study.
The C. elegans infection model is intended to augment rather than replace mammalian models. The nematode lacks an adaptive immune system and specialized immune cells, such as macrophages and neutrophils, which are essential components of the immune response during Candida infections in mammals. The C. elegans experiments are performed at a temperature below that of the mammalian body. However, we have shown that the nematode can effectively identify significant phenotypes, which can then be further investigated using mammalian models.
C. elegans offers a rapid and effective approach for determining potential virulence factors. It has been used to screen insertion mutant libraries in P. aeruginosa, E. faecalis, C. neoformans, and C. albicans, leading to the identification of both established and novel virulence factors (87–90). Beyond these pathogens, C. elegans has also been employed to study rare pathogens. For example, the novel virulence factor AidA was identified partly through C. elegans experiments in the Burkholderia cenocepacia cystic fibrosis isolate H111 (91). Yersinia pestis kills C. elegans through a biofilm-dependent mechanism, highlighting the versatility of this model in uncovering diverse pathogenic strategies (76). Our ultimate goal is to utilize the nematode to identify novel virulence factors in fungal pathogens. While the SYTOX assay has most commonly been used to identify and optimize novel antimicrobials in C. albicans, here we demonstrated that it can be used to test for virulence in C. auris, and this may have advantages in the genetic dissection of virulence in this haploid species.
MATERIALS AND METHODS
Microbial strains and media
For C. elegans maintenance and propagation, animals were fed E. coli strain OP50 on nematode growth medium agar using standard techniques (92). Fungal strains were routinely grown in YPD medium. C. albicans wild-type strain SC5314 was used as a control for most experiments (93). Mutants were generated in the AR0382 strain (a.k.a. B11109, CDC Antimicrobial Resistance Bank). C. auris was transformed using a modified CRISPR-Cas9 approach (63, 94–96). In short, CAS9 was PCR-amplified from pRB732. We selected specific guide RNAs and PCR-amplified them using pTO136 as a template. A repair construct was designed by flanking the nourseothricin resistance marker with approximately 500 base pairs targeting the gene ADE2 (97). To create the complement strain, the ADE2 gene was reintegrated via homologous recombination at the native locus. C. auris strains AR0381 (B11220, clade II), AR0382 (B11109, clade I), AR0383 (B11221, clade III), and AR0386 (B11245 IV) used for fluorescence microscopy were also obtained from the CDC Antimicrobial Resistance Bank. These strains were tagged with the fluorophore mScarlet as previously described (98). As previously described, C. albicans SC5314 was tagged with fluorophore dTomato (99).
C. elegans survival assay
C. elegans glp-4(bn2);sek-1(km-4) were used for all experiments. The survival assays were performed as in early studies (56, 57, 61, 100). Infection plates were prepared by growing fungal strains in YPD liquid media for 24 hours at 30°C with shaking. Five hundred microliters of the culture was plated onto BHI solid media containing gentamycin at 10 μg/mL for 24 hours at 30°C. Synchronized L4 stage nematodes were decanted on the fungal lawns and allowed to graze for 4 hours. The nematodes were washed with M9 pipetted into six-well plates (approximately 30 worms per well, two wells per condition) containing 2 mL of 20% BHI 80% M9. Plates were incubated at 25°C, and worm death was scored daily.
Fluorescence microscopy of infected animals
To investigate colonization of the intestine during infection, worms were infected with mScarlet and dTomato tagged strains as described above for the survival assay. After 6 days, live worms were washed with M9 and mounted on 2% agarose pads. The animals were imaged using an Olympus FLUOVIEW FV 300 confocal microscope imaging system and Fluoview FC315-SW software.
High-throughput SYTOX Orange assay
The SYTOX Orange assay was performed as in earlier studies (58, 71–73). L4 stage nematodes were pipetted into a 96-well plate (about 30–50 worms per well, 3+ wells per condition in assays with controls only, one well per condition in isolate library screen) in 20% BHI 80% M9 plus gentamycin at 10 μg/mL concentration. In most assays, the wells were inoculated with fungal culture grown for 16 hours at 30°C with shaking and standardized to a final optical density at 600 nm of 0.03. For screening the isolate library, the strains were grown in 200 μL of YPD liquid media for 16 hours at 30°C with shaking in a 96-well plate. One microliter per condition was used to inoculate the worm plate. After 5 days, the plates were washed with M9 and stained with Invitrogen SYTOX Orange Nucleic Acid Stain (diluted 2 μL to 10 mL of M9). Plates were washed and imaged using a BioTek Cytation5 Imaging Reader. Fluorescence and bright-field images were taken of each well. The TRITC filter cube was used for fluorescent imaging. The automated Cell Profiler pipeline was used to determine the percentage of dead worms. This pipeline counts the black pixels in the bright-field image and red pixels in the fluorescent image. The ratio of red to black pixels determines the percentage of dead worms.
Statistics
Kaplan-Meier survival curves were generated and compared using Mantel-Cox log-rank analysis. For SYTOX assays, unpaired t-tests or one-way ANOVA with multiple comparisons and Dunnett’s test were used to compare conditions. GraphPad Prism was used for statistical analysis. For all statistical tests, P values of <0.05 were considered statistically significant, and asterisks in the figure panels indicate the levels of significance as follows: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Satoh K, Makimura K, Hasumi Y, Nishiyama Y, Uchida K, Yamaguchi H. 2009. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol Immunol 53:41–44. doi:10.1111/j.1348-0421.2008.00083.x 19161556 · doi ↗ · pubmed ↗
- 2Cristina ML, Spagnolo AM, Sartini M, Carbone A, Oliva M, Schinca E, Boni S, Pontali E. 2023. An overview on Candida auris in healthcare settings. J Fungi 9:913. doi:10.3390/jof 9090913 PMC 1053297837755021 · doi ↗ · pubmed ↗
- 3Lionakis MS, Chowdhary A. 2024. Candida auris infections. N Engl J Med 391:1924–1935. doi:10.1056/NEJ Mra 240263539565991 · doi ↗ · pubmed ↗
- 4Haq MF, Pearlmutter BS, Cadnum JL, Donskey CJ. 2024. Efficacy of 23 commonly used liquid disinfectants against Candida auris isolates from the 4 major clades. Infect Control Hosp Epidemiol 45:127–131. doi:10.1017/ice.2023.15737528766 · doi ↗ · pubmed ↗
- 5Omardien S, Teska P. 2024. Skin and hard surface disinfection against Candida auris – what we know today. Front Med 11. doi:10.3389/fmed.2024.1312929 PMC 1087957138384416 · doi ↗ · pubmed ↗
- 6Horton MV, Johnson CJ, Kernien JF, Patel TD, Lam BC, Cheong JZA, Meudt JJ, Shanmuganayagam D, Kalan LR, Nett JE. 2020. Candida auris forms high-burden biofilms in skin niche conditions and on porcine skin. m Sphere 5:e 00910-19. doi:10.1128/m Sphere.00910-1931969479 PMC 6977180 · doi ↗ · pubmed ↗
- 7Eix EF, Nett JE. 2022. Modeling Candida auris skin colonization: mice, swine, and humans. P Lo S Pathog 18:e 1010730. doi:10.1371/journal.ppat.101073036074786 PMC 9455849 · doi ↗ · pubmed ↗
- 8Armstrong PA, Rivera SM, Escandon P, Caceres DH, Chow N, Stuckey MJ, Díaz J, Gomez A, Vélez N, Espinosa-Bode A, Salcedo S, Marin A, Berrio I, Varón C, Guzman A, Pérez-Franco JE, Escobar JD, Villalobos N, Correa JM, Litvintseva AP, Lockhart SR, Fagan R, Chiller TM, Jackson B, Pacheco O. 2019. Hospital-associated multicenter outbreak of emerging fungus Candida auris, Colombia, 2016. Emerg Infect Dis 25:1339–1346. doi:10.3201/eid 2507.18049131211679 PMC 6590770 · doi ↗ · pubmed ↗