qacA is a key factor in heteroresistance to vancomycin in sequence type 5 methicillin-resistant Staphylococcus aureus isolates from pneumonia patients
Kaiting Zhang, Lin Xi, Qiyu Bian, Yu Yin, Mengting Chen, Ping Yang, Zhouzhou Chen, Hailan Wu, Yongqiang Zhu, Huajun Zheng, Daijie Chen, Yaxin Fan, Jing Zhang

TL;DR
The study finds that the gene qacA plays a key role in making some MRSA strains less responsive to vancomycin in pneumonia patients.
Contribution
The study identifies qacA as a novel independent predictor of hVISA in ST5-MRSA through genetic and experimental validation.
Findings
qacA is a significant predictor of hVISA in ST5-MRSA isolates from pneumonia patients.
ST5-MRSA strains with qacA exhibit a high hVISA detection rate compared to ST764-MRSA.
Acquisition of the qacA plasmid transforms VSSA strains into hVISA phenotypes.
Abstract
The presence of heterogeneous vancomycin-intermediate Staphylococcus aureus (hVISA) among methicillin-resistant Staphylococcus aureus (MRSA) strains has been associated with vancomycin treatment failure, especially in cases of MRSA pneumonia. This study aimed to analyze the molecular characteristics and risk factors for hVISA among MRSA strains isolated from pneumonia patients and to validate the most important risk factors. All the MRSA clinical isolates were collected in a multicenter, prospective, observational clinical trial from July 2012 to June 2020. The hVISA strains were verified via a modified population analysis profile–area under the curve method, and the prevalence of hVISA in hospitalized pneumonia patients was 53.5% (61/114). Sequence type 5 (ST5)-MRSA, the dominant hVISA strain, and ST764-MRSA, the dominant vancomycin-susceptible S. aureus (VSSA) strain, exhibited…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
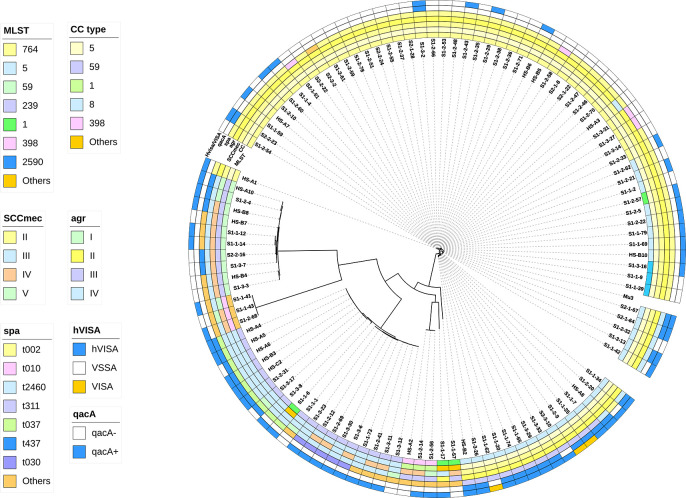 Fig 1
Fig 1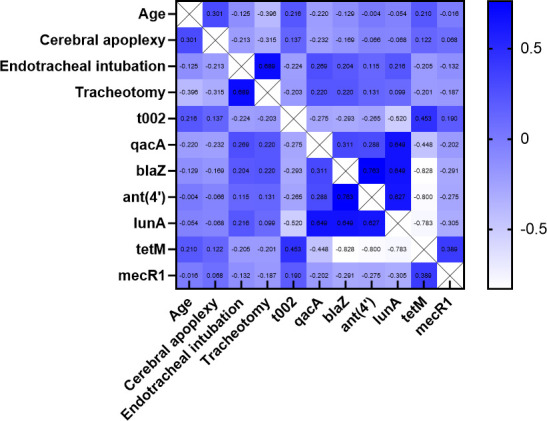 Fig 2
Fig 2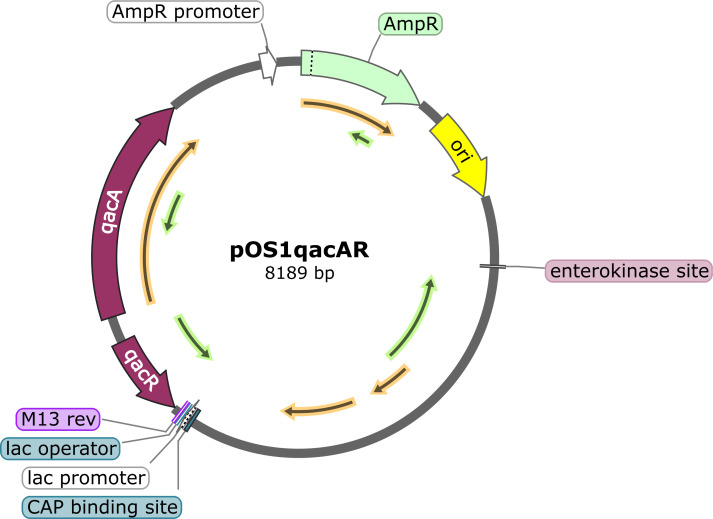 Fig 3
Fig 3| Gene | Annotation | Total ( | ST5-MRSA ( | ST764-MRSA ( |
|---|---|---|---|---|
|
| Signal transduction gene, penam antibiotic target replacement | 63 (86.3%) | 23 (74.2%) | 40 (95.2%) |
|
| Tetracycline-resistant ribosomal protection target | 58 (79.5%) | 16 (51.6%) | 42 (100%) |
| Aminoglycoside antibiotic inactivation | 21 (28.8%) | 17 (54.8%) | 4 (9.5%) | |
|
| β-Lactamase inactivation | 20 (27.4%) | 20 (64.5%) | 0 (0%) |
|
| Major facilitator superfamily antibiotic efflux pump | 20 (27.4%) | 18 (58.1%) | 2 (4.8%) |
|
| Lincosamide-nucleotidyl transferase antibiotic inactivation | 10 (13.7%) | 10 (32.3%) | 0 (0%) |
| Characteristic | Total ( | ST5-MRSA ( | ST764-MRSA ( | |
|---|---|---|---|---|
| Demographic | ||||
| Age (years) | 72 (57–81) | 63 (50–75) | 75 (60–83) |
|
| Sex, male | 46 (63.0%) | 17 (54.8%) | 29 (69.0%) | 0.214 |
| Weight (kg) | 60 (55–70) | 60 (54–70) | 60 (55–70) | 0.842 |
| BMI (kg/m2) | 21.7 (19.5–23.6) | 22.2 (19.8–23.0) | 21.5 (19.0–24.0) | 0.843 |
| ICU | ||||
| ICU admission | 50 (68.5%) | 20 (64.5%) | 30 (71.4%) | 0.530 |
| ICU stay (days) | 24 (7–38) | 27 (13–40) | 21 (2–35) | 0.386 |
| Underlying disease/condition | ||||
| Cardiovascular disease | 25 (34.2%) | 7 (22.6%) | 18 (42.9%) | 0.071 |
| Diabetes | 11 (15.1%) | 3 (9.7%) | 8 (19.0%) | 0.438 |
| Cerebral apoplexy | 28 (38.4%) | 6 (19.4%) | 22 (52.4%) |
|
| Dialysis | 2 (2.7%) | 1 (3.2%) | 1 (2.4%) | >0.999 |
| COPD | 4 (5.5%) | 1 (3.2%) | 3 (7.1%) | 0.836 |
| Autoimmune disease | 6 (8.2%) | 2 (6.5%) | 4 (9.5%) | 0.967 |
| Trauma | 12 (16.4%) | 6 (19.4%) | 6 (14.3%) | 0.564 |
| Solid tumor | 1 (1.4%) | 1 (3.2%) | 0 (0%) | 0.425 |
| Operation | 30 (41.1%) | 16 (51.6%) | 14 (33.3%) | 0.117 |
| Implant | ||||
| Venous catheter | 52 (71.2%) | 20 (64.5%) | 32 (76.2%) | 0.276 |
| Endotracheal intubation | 26 (35.6%) | 16 (51.6%) | 10 (23.8%) |
|
| Tracheotomy | 21 (28.8%) | 16 (51.6%) | 5 (11.9%) |
|
| PD | ||||
| MIC50 (mg/L) | 1 | 1 | 0.5 | NA |
| MIC90 (mg/L) | 1 | 1 | 1 | NA |
| Molecular typing | ||||
| SCC | 72 (98.6%) | 30 (96.8%) | 42 (100%) | 0.425 |
| | 72 (98.6%) | 31 (100%) | 41 (97.6%) | >0.999 |
| | 46 (63.0%) | 9 (29.0%) | 37 (88.1%) |
|
| Risk factor | Univariate analysis (OR [95% CI]) | Multivariate analysis | |
|---|---|---|---|
| aOR (95% CI) | |||
| Age (years) | 0.834 (0.924–0.988) | ||
| Cerebral apoplexy | 0.423 (0.109–1.646) | ||
| Endotracheal intubation | 3.010 (0.731–12.398) | ||
| 0.152 (0.051–0.454) | |||
|
| 36.944 (4.571–289.601) | 48.323 (6.004–366.565) | <0.001 |
| 3.409 (1.140–10.190) | |||
|
| 0.089 (0.011–0.744) | ||
| ST type | Strain serial number | Vancomycin MIC (mg/L) | AUCtest | AUCtest/AUCMu3 | hVISA/VSSA |
|---|---|---|---|---|---|
| ST5 | S1-2-32 | 0.5 | 22.51 | 0.8 | VSSA |
| S1-2-32 | 0.5 | 26.69 | 1.0 | hVISA | |
| ST764 | S1-2-46 | 0.5 | 22.06 | 0.8 | VSSA |
| S1-2-46 | 0.5 | 28.18 | 1.1 | hVISA | |
| Mu3 (positive control) | 0.5 | 26.93 | hVISA | ||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Nosocomial Infections in ICU · Antibiotic Resistance in Bacteria
INTRODUCTION
Methicillin-resistant Staphylococcus aureus (MRSA) is a prevalent nosocomial pathogen that causes severe conditions such as hospital-acquired pneumonia (HAP), bloodstream infections (BSIs), complicated skin and soft tissue infections, and infective endocarditis, resulting in significant morbidity and mortality (1). MRSA infection imposes a high and increasing burden on the healthcare system, ranking second in the assessment of public health burden and accounting for 19.2% of the total disability-adjusted life-years (2). The observed contradiction between the gradually declining prevalence of MRSA isolates in some regions, including China, and the increasing complexity of MRSA infection is partially a consequence of treatment advances and the ability of the pathogen to adapt to changing environments via alternative pathways (3, 4).
Vancomycin, which was introduced in the clinic more than 60 years ago, remains an effective intervention to treat MRSA infection. No vancomycin-resistant S. aureus strains have been identified in China, but an increasing prevalence of heterogeneous vancomycin-intermediate Staphylococcus aureus (hVISA) and vancomycin-intermediate S. aureus (VISA) strains has emerged under external selection pressure with wider utilization of and longer treatment with vancomycin. Multiple retrospective analyses have revealed that the hVISA phenotype is a potential risk factor for predicting vancomycin treatment failure and is responsible for considerable morbidity (5, 6).
Currently, the predominant hospital-acquired MRSA (HA-MRSA) genetic types globally are sequence type (ST) 239, ST36, and sequence type 5 (ST5), while ST239, ST241, and ST5 are the top three types in Asia (7). We previously identified ST5 as a risk factor for bacterial persistence in patients with MRSA pneumonia, and adults with ST5 MRSA infection showed worse clinical signs and symptom improvement than did those with ST764 infection (8, 9).
Notably, the epidemiologic features of hVISA/VISA strains were closely related to those of the ST5 and ST239 clones. A meta-analysis of hVISA epidemiology revealed that ST239 and ST5 are the most common epidemic genotypes of hVISA, with prevalence rates of 58.62% and 14.45%, respectively. Both are international HA-MRSA lineages prevalent in Asia, South America, and Eastern Europe, with Asia having the highest incidence (10).
It is likely that the features of hVISA are closely related to cell wall genetic mutations in ST5-MRSA, especially in two-component systems such as walKR, vraSR, and graSR, considering that the target of vancomycin is D-Ala-D-Lac, a material involved in bacterial peptidoglycan synthesis (11). However, the decreased susceptibility to antibiotics and the subsequent development of resistance often involve a multitude of diverse and interrelated mechanisms. Among them, efflux pumps may represent one of the most rapid and effective pathways under external stress, which has been well established in gram-negative bacteria; however, few studies have explored the genetic characteristics of hVISA strains in depth (12).
Retrospective studies have shown that the carriage of qacA, an S. aureus-specific efflux pump, is significantly associated with the hVISA phenotype in a single intensive care unit (ICU)-acquired MRSA strain, ST5 (13), but little is known about other MRSA infections in different regions or about this association. This study aimed to determine the molecular characteristics and risk factors for hVISA among MRSA strains isolated from pneumonia patients in China and then reveal and experimentally confirm the correlation between qacA and hVISA formation in ST5-MRSA.
RESULTS
Clinical MRSA isolates and the hVISA phenotype in pneumonia patients
All the MRSA clinical isolates were collected from a multicenter, prospective observational clinical trial conducted from July 2012 to June 2020. A total of 114 clinical MRSA strains were isolated from the sputum of Chinese patients diagnosed with MRSA pneumonia. Routine in vitro vancomycin susceptibility testing was performed using the agar dilution method. The MIC_50_ and MIC_90_ were both 1 mg/L, indicating that all 114 MRSA strains collected were sensitive to vancomycin.
hVISA strains were screened using the modified population analysis profile–area under the curve (PAP-AUC) method (14). Among these strains, 53.5% (61/114) hVISA and 2.6% (3/114) VISA strains were identified (Fig. 1). The significantly higher hVISA rate than that reported in historical data both globally and in China highlights the typical reduced vancomycin susceptibility of our MRSA strains. The MIC_50_ and MIC_90_ for hVISA/VISA were both 1 mg/L, whereas for the vancomycin-susceptible S. aureus (VSSA) strains, the MIC_50_ and MIC_90_ were 0.5 and 1.0 mg/L, respectively.
Phylogenetic tree of 114 MRSA strains with molecular typing, hVISA phenotype, and qacA carriage information.
Molecular typing of MRSA clinical isolates
Whole-genome sequencing (WGS) was performed on all 114 clinical MRSA isolates included in this study. A phylogenetic tree of 114 MRSA strains with CC, MLST, SCCmec, agr, and spa molecular typing is shown in Fig. 1.
CC5-SCCmecII-agrII-t002 was the dominant clone (48/114, 42.1%) in our study. CC5 was the most common epidemic subtype, accounting for 67.5% (77/114) of all the isolates, among which ST5 (31/77, 40.2%) and ST764 (42/77, 54.5%), differing in only one housekeeping gene, aroE, were the prevalent subtypes in the CC5 family. The SCCmecII type accounted for 66.7% (76/114) of all isolates, suggesting a hospital-related background for the MRSA strains. The agrII genotype, which is distributed mainly in S. aureus isolates that are moderately resistant to glycopeptides, accounted for 67.5% (77/114) of all the MRSA strains. spa typing revealed diversity, as the main class was t002 (50/114, 43.9%), followed by t311 (12/114, 10.5%) and t2460 (10/114, 8.8%).
Notably, ST5 was the most prevalent hVISA subtype (25/61, 41%), and all the VISA strains (3/3, 100%) identified belonged to the ST5 family. The main VSSA clone was ST764 (33/50, 66%). The vancomycin MIC assay results revealed that both the MIC_50_ and MIC_90_ of ST5-MRSA were 1 mg/L, whereas ST764-MRSA had an MIC_50_ of 0.5 mg/L and an MIC_90_ of 1.0 mg/L. The hVISA detection rate in ST5-MRSA was significantly greater than that in ST764-MRSA (90.3% vs 21.4%, P < 0.001).
The two phylogenetically close clonal types exhibited diverse phenotypes in terms of vancomycin susceptibility, indicating that further exploration of ST5-MRSA could reveal the mechanism underlying the high hVISA incidence via comparison against that of ST764-MRSA.
Genes encoding antimicrobial resistance factors were further investigated in the ST5-MRSA (n = 31) and ST764-MRSA (n = 42) clinical isolate groups via BLAST analysis against the Comprehensive Antibiotic Resistance Database (CARD) (15) (Table S1). In total, 44 antimicrobial resistance genes were identified from the ST5-MRSA and ST764-MRSA strains. Twenty-five genes, including mecA, mecR, tet(38), parC, gyrA, and arlR, were carried by all 73 strains, whereas six genes [norA, rpsJ, mecl, ant(9′)-la, uhpt, and fosB3] presented marginal differences between the two groups, with detection rates greater than 90%, indicating the high potential for resistance of the MRSA strains to multiple antimicrobial agents. Three genes (mupA, fusC, and cat7A) presented a low prevalence (<10%) in both groups. Notably, the prevalence of six genes [qacA, blaZ, ant(4′), lunA, tetM, and mecR1] significantly differed between the ST5-MRSA and ST764-MRSA groups (Table 1). qacA, blaZ, ant(4′), and lunA were the beneficial genes in ST5-MRSA, whereas mecR1 and tetM were the beneficial genes in ST764-MRSA.
Clinical characteristics of patients with ST5- and ST764-MRSA pneumonia
The clinical characteristics of the 73 enrolled patients with pneumonia caused by infection with ST5-MRSA or ST764-MRSA are summarized in Table 2. More than half of the patients were male (63%), with a median age of 72 (57–81) years. Most of the patients (68.5%) had a history of intensive care unit (ICU) admission, with a median stay of 24 (7–38) days. Histories of cardiovascular disease and cerebral apoplexy were found in 34.2% and 38.4% of the pneumonia patients, respectively. Invasive procedures such as venous catheter implantation, endotracheal intubation, and tracheotomy were implemented in 71.2%, 35.6% and 28.8% of the patients, respectively. The MIC_90_ of vancomycin for the 73 ST5 and ST764 MRSA isolates was 1 mg/L, while the MIC_50_ was higher for the ST5 strain (1 mg/L) than for the ST764 strain (0.5 mg/L).
Predictive factors for hVISA occurrence in ST5-MRSA compared with that in ST764-MRSA
The ST5-MRSA and ST764-MRSA clinical isolate groups were further compared in terms of hosts, vancomycin pharmacodynamics (PD) and bacterial factors. As shown in Table 2, in terms of clinical features, patients in the ST5-MRSA group were younger (63 vs 75 years) and more likely to have undergone endotracheal intubation (51.6% vs 23.8%) and tracheotomy (51.6% vs 11.9%) than those in the ST764-MRSA group were; moreover, patients infected with ST764-MRSA had a greater incidence of cerebral apoplexy (52.4% vs 19.4%). In addition, the spa-t002 genotype was the dominant variant in ST764-MRSA over ST5-MRSA (88.1% vs 29.0%).
Correlation analysis was performed on all significant single variables from the host, vancomycin PD, and genotyping comparisons between the ST5-MRSA and ST764-MRSA strains (Fig. 2) to assist in variable selection for subsequent multivariate analysis. Seven variables, including age, cerebral apoplexy, endotracheal intubation, spa-t002, qacA, ant(4′) and mecR1, were included in the multivariate logistic regression analysis of hVISA occurrence.
Correlation analysis of significant variables related to the host, vancomycin treatment, and genotype.
Finally, multivariate analysis (Table 3) revealed that the qacA gene was an independent predictor of hVISA occurrence in ST5-MRSA strains (aOR, 48.323; 95% CI, 6.004–366.565; P < 0.001). The Hosmer–Lemeshow goodness-of-fit test revealed that the model fit well (P > 0.05).
Carriage of the efflux pump gene qacA mediated hVISA occurrence in ST5-MRSA isolates
The qacA carriage rates of our MRSA isolates are presented in Fig. 1. A total of 58.1% (18/31) of the ST5-MRSA strains carried the qacA gene, and the spa subtypes t311 and t2460 accounted for 32.3% (10/31) and 25.8% (8/31) of the strains, respectively. Approximately half (13/31) of the ST5-MRSA strains were qacA(−), and most of the qacA(−)-ST5-hVISA strains belonged to the t002 subtype (29.0%, 9/31). The transmission of qacA was clearly lineage dependent, and compared with ST5-t002, ST5-t311 and ST5-t2460 were more advantageous clones.
The ability of qacA carriage in ST5-MRSA to mediate phenotypic transformation from VSSA to hVISA was verified by observing the phenotypic changes after the electroporation of qacA-borne plasmids into specific recipient strains of the ST5 (S1-2-32) and ST764 (S1-2-46) strains (Table 4). Polymerase chain reaction (PCR) experiments confirmed that all the plasmids were successfully transferred into the recipient strains. An in vitro vancomycin susceptibility test revealed no significant change in the MIC values for each strain that acquired the qacA-harboring plasmid. After electroporation with a plasmid carrying qacA, the ST5-MRSA (S1-2-32qacA) and ST764-MRSA (S1-2-46qacA) strains clearly exhibited an hVISA phenotype, as confirmed by the modified PAP-AUC method.
DISCUSSION
hVISA was first isolated in 1997 from the pathogen Mu3 collected from sputum samples of pneumonia patients (15), after which emerging cases were reported in various countries and regions. A meta-analysis of hVISA/VISA epidemiology demonstrated overall prevalence rates of 6.05% and 3.01% for hVISA and VISA, respectively (10). Our analysis utilized clinical MRSA strains collected from MRSA-infected HAP patients and revealed a significantly greater incidence of hVISA (53.5%, 61/114) via PAP-AUC, which was more than triple the combined prevalence rate for hVISA in mainland China (15.78%) (16–19). Because MRSA infection occurrence, especially healthcare-associated infections, is increased in Asian countries (20), this has led to more frequent use of vancomycin, which poses selective pressure on endemic MRSA strains. An investigation of the mechanisms underlying the emergence of regional hVISA has significant clinical implications for treatment with vancomycin.
The genetic prevalence of hVISA is dominated by ST239 (58.62%) and ST5 (14.45%) globally, and almost half of hVISA strains are SCCmecII-type strains (48.16%) (10). Our MRSA isolates presented the reverse genetic profile of hVISA, with ST5 (41.0%, 25/61) being the most common epidemic variant, followed by ST239 (23.0%, 14/61), whereas SCCmecII (63.9%, 39/61) accounted for more than half of all the hVISA strains. This SCCmecII majority was in line with the healthcare background of our MRSA strains isolated from a nosocomial environment, but the unique and subtle dominance of ST5 over ST239 in our hVISA strains from hospitalized pneumonia patients made further investigation of the underlying factors worthwhile.
Many studies have focused on describing the epidemiology and molecular characteristics of hVISA/VISA strains directly, whereas few have focused on choosing a suitable reference to perform in-depth comparisons; such a reference would ideally be phylogenetically similar but phenotypically different from the typical hVISA clone. As a single-locus variant of ST5-MRSA, the ST764 clone diverged from the ST5 lineage via a single genetic mutation and is a hybrid variant with community-associated features. ST764 has gradually become prevalent among nosocomial MRSA infections in Asian countries in recent decades (21, 22). A recent study reported that ST5/ST764-SCCmecII was the most common type among MRSA-associated BSIs in southern China (23). Interestingly, our study identified ST5/ST764-SCCmecII as the dominant clone in HAP MRSA infections (63.2%, 72/114), and the similar proportions of the ST5 (27.2%, 31/114) and ST764 (36.8%, 42/114) clonotypes indicate possible coevolution, given their genetic relatedness. In contrast to the high frequency of ST5 among hVISA strains, ST764 was the representative lineage among VSSA strains. The different MIC_50_ values of ST5-MRSA (MIC_50_ = 1 mg/L) and ST764-MRSA (MIC_50_ = 0.5 mg/L) also indicated their phenotypic diversity in terms of hVISA, as changing vancomycin MICs directly impact hVISA rates, because the proportion of hVISA isolates increases with increasing vancomycin MICs, even within the susceptible range (24). Thus, the ST5-MRSA and ST764-MRSA strain groups represent a suitable match to further elucidate the mechanism underlying the high hVISA frequency in ST5-MRSA.
Depending on the antibiotic challenge imposed, efflux could be the fastest-acting and most effective resistance mechanism in the bacterial repertoire of stress responses and is not only commonly observed in gram-negative species but also a significant contributing factor to gram-positive resistance. Efflux pumps can dynamically transport not only clinically relevant antimicrobial agents but also environmental biocides and disinfectants that colonize the healthcare environment; these multiple structurally unrelated compounds result in a multidrug resistance (MDR) phenotype (12, 25). Compared with that of ST764-MRSA, the efflux pump gene qacA was identified in our study as a distinguishable gene carried by ST5-MRSA as a predictor of hVISA formation. The qacA gene encodes the QacA protein, which is an S. aureus-specific MDR efflux pump that uses a proton motive force-dependent antiport mechanism to drive the export of various cationic and lipophilic compounds, such as chlorhexidine, a commonly used decolonization agent for MRSA in hospitals, to mediate S. aureus resistance or reduce susceptibility to many clinical antibiotics (26). QacA genes are plasmid encoded and thus are relatively easily transferred horizontally between species and clonotypes. The expression of plasmid-based genes is usually sufficient for relevant resistance to be observed without the need for additional mutations because of the multicopy nature of these genetic elements (25).
Multiple studies have indicated that clinical strains carrying the qacA gene are predominantly hospital-associated pathogens such as ST5-MRSA (27–29), which is consistent with our finding that the ST5 type accounted for a majority (66.6%, 18/27) of the qacA(+)-MRSA strains. Interestingly, all the qacA(+)-ST5-MRSA strains were hVISA/VISA strains, which have not been previously reported. Although little is known about whether qacA is associated with reduced vancomycin susceptibility or with ST5-MRSA, experiments and analyses have been conducted to investigate this aspect. After successful electroporation of the qacA-borne plasmid extracted from the qacA(+)-ST5-hVISA strain into the qacA(−)-ST5-VSSA and qacA(−)-ST764-VSSA strains, the hVISA phenotype was observed in the electroporated strains. In addition to the selective advantage for ST5-MRSA in hospital environments, qacA carriage also provided it with reduced vancomycin susceptibility as a significant contributor to the transformation of VSSA to hVISA.
Moreover, one qacA(−)-ST764-VSSA strain was also selected for the electroporation experiment, in addition to the qacA(−)-ST5-VSSA strain, as we aimed to explore whether qacA-mediated hVISA formation is lineage dependent. In almost half of the qacA(−)-VSSA strain group (41.2%, 47/114), the abundance of the ST764 lineage was more than 10 times greater (68.1%, 32/47) than that of the ST5 lineage (6.4%, 3/47). The successful phenotypic change in the qacA(−)-ST764-VSSA strain indicated that the generality of qacA carriage for hVISA formation was not restricted to ST5-MRSA but was applicable to other genotypes. The significantly greater carriage of qacA in our ST5-MRSA isolates may have contributed to the higher hVISA detection rate than that in ST-764. With MRSA infection expanding from the nosocomial environment to the community environment, the association of qacA with hVISA formation identified from hospital isolates of ST5-MRSA also has clinical implications for community-characterized MRSA strains, such as ST764-MRSA strains.
Notably, an association of qacA with the hVISA phenotype was previously reported by Cho et al. (13) among MRSA strains in the ICU during decolonization with chlorhexidine. Our study further revealed a similar significant correlation among MRSA isolates from pneumonia patients. However, similar conclusions have been drawn from different ratiocinations, as our research focused on the significantly higher hVISA rate observed in pneumonia patients infected with MRSA, and stepwise analysis revealed that qacA is a risk factor for hVISA formation in ST5-MRSA. However, their work focused directly on the spread of qacA in patients undergoing chlorhexidine-based MRSA decolonization and revealed its potential link to clinical characteristics, including the hVISA phenotype. Moreover, associations may be nonspecifically detected, especially from clinical MRSA isolates with complex backgrounds and many confounding factors. Thus, our work further confirmed this association as transferable and reproducible in terms of phenotype change to hVISA in experimentally electroporated ST5 and ST764 MRSA strains after the acquisition of the qacA-borne plasmid. Our MRSA strains were collected from 10 hospitals nationwide, which is geographically representative of MRSA epidemiology in China and in line with the findings from a single ICU in Korea, with some commonalities in the pattern in which qacA affects hVISA occurrence in Asia.
Admittedly, our analysis has several limitations. The molecular mechanisms underlying the development of hVISA strains are interactive and confounding, and a comprehensive assessment could still be applied to further understand the correlation. However, we believe that the findings do not undercut the evident association of qacA with hVISA formation in the ST5-MRSA strains identified in our study. Further work is needed to explore the possible mechanism underlying this phenomenon in terms of qacA expression, qacA-mediated vancomycin recognition, and extrusion to confirm the identified correlation.
In summary, a higher frequency of hVISA than typically recorded was observed in clinical MRSA pneumonia isolates, with ST5 being the dominant hVISA strain and ST764 being the dominant VSSA strain. The efflux pump gene qacA was identified as a significant predictor of hVISA formation in the ST5-MRSA strains. qacA carriage is lineage-specific in that the ST5-t311 and ST5-t2460 MRSA strains are advantageous clonal variants over ST5-t002 in terms of qacA carriage. The qacA-borne plasmid promoted the transformation of ST5-MRSA from VSSA to hVISA as an adaptive response that contributed to reduced vancomycin susceptibility during infection in healthcare settings.
Further research on qacA in ST5-MRSA is warranted in an effort to improve our understanding of how these systems interact with vancomycin. This knowledge may inform the development of means to postpone the occurrence of vancomycin resistance during MRSA infection in pneumonia patients and provide innovative perspectives on the effectiveness of vancomycin.
MATERIALS AND METHODS
Bacterial isolates
This study was conducted at 10 teaching hospitals from July 2012 to June 2020 and used a database containing data collected from two multicenter, prospective observational clinical studies.
All clinical isolates of S. aureus were collected, and identical strains from the same patient were excluded. The MICs of oxacillin and vancomycin were determined by the agar dilution method at a CHINET microbiology laboratory. Strains that grew at an oxacillin concentration of ≥4 mg/L were identified as MRSA.
hVISA screening
A modified PAP-AUC method with vancomycin concentrations of 0.5, 1.0, 2.0, 2.5, 4.0, and 8.0 mg/L was adopted to screen for hVISA strains (14). Tested isolates with an AUC ratio against the control strain (Mu3, ATCC 700698) greater than 1.3 were defined as VISA strains; those with ratios between 0.9 and 1.3 were defined as hVISA strains; and those with ratios less than 0.9 were defined as VSSA strains.
WGS and genotyping
DNA from MRSA strains was extracted with a Takara MiniBEST Bacteria Genomic DNA Extraction Kit v.3.0 (Takara Biomedical Technology Co., Ltd., Beijing, China). A 300 bp double-ended library was generated using the NEXTflex DNA Sequencing Kit (Bio Scientific, USA) and sequenced on an Illumina X10 platform (Illumina, San Diego, CA, USA) via 2 × 150 bp paired-end sequencing.
The sequence data of seven housekeeping genes (arcC, aroE, glpF, gmk, pta, tpi, and yqiL) were analyzed for multifocal sequence typing according to the PubMLST database (https://pubmlst.org/). Six of seven identical housekeeping genes belonged to the same clonal complex (CC). SCCmec types were identified using specific gene fragments of SCCmecI-V described by Oliveira et al. (30) and Hanssen and Sollid (31). The agr gene of each strain was compared with four genes in the NCBI GenBank database: agrI (GenBank accession no. AF210055), agrII (GenBank accession no. AF001782), agrIII (GenBank accession no. AF001783), and agrIV (GenBank accession no. AF288215). The strains were assigned to the spa genotypes according to the Ridom SpaServer database (http://spaserver.ridom.de/). BLAST software was used to determine the similarity and respective length between sequences. The genomes of the MRSA strains were screened for antimicrobial resistance-related genes using the CARD (https://card.mcmaster.ca/). The phylogenetic tree was integrated with molecular typing data, the hVISA phenotype, and qacA carriage using iTOL v.5 (https://itol.embl.de/).
Plasmid analysis and electroporation experiments
The plasmid electroporation procedure consisted of five steps: extraction of the vancomycin-resistant S. aureus plasmid (VRSAp) (a typical plasmid from our clinical strains carrying qacA), construction of the pOS1qacAR plasmid, transfer of the pOS1qacAR plasmid into DH5α cells, electroporation of the pOS1qacAR plasmid into JMC1 cells, and electroporation of the JMC1 demethylated pOS1qacAR plasmid into recipient strains.
A representative ST5-MRSA strain (serial number S1-1-76, spa-t2460) for which the qacA(+)-plasmid VRSAp was extracted using a plasmid extraction kit with the plasmid concentration determined by a NanoDrop was selected as the plasmid provider. The pOS1qacAR plasmid (Fig. 3) was prepared by joining the qacAR fragment from the VRSAp template with qacAR-F/qacAR-R as upstream and downstream PCR primers with the pOS1 plasmid fragment separated from Escherichia coli DH5α. We simultaneously introduced the upstream repressor protein-encoding gene qacR to eliminate its influence on qacA expression. The pOS1qacAR plasmid was subsequently transformed into DH5α cells, and pOS1VF/pOS1VR were used as primers for PCR and sequencing verification. Afterward, the complete pOS1qacAR plasmid was electroporated into the JMCI strain to obtain a demethylated pOS1qacAR plasmid, which is more easily accepted by recipient strains. Once the recipient strain competent cells were obtained, the demethylated pOS1qacAR plasmid was electroporated and subjected to PCR with the pOS1VF/pOS1VR primers, using the pOS1qacAR plasmid as a positive control, to obtain the corresponding qacA expression strains. The strains, plasmids, and primers used are summarized in Tables S2 to S4, respectively.
pOS1qacAR plasmid map.
Statistical analysis
All the statistical analyses were performed using SPSS 19 (SAS Institute, Cary, NC, USA). Categorical variables are presented as descriptive statistics, such as the number of cases (percent), and continuous variables are presented as medians (interquartile ranges). Univariate comparisons were performed using Fisher’s exact test for categorical variables and the Mann‒Whitney U test for continuous variables. P values less than 0.05 were considered to indicate statistical significance. All significant variables were included in the correlation analysis using GraphPad Prism 8.0 software. Variables with correlation coefficients less than 0.5 and those deemed potentially relevant to the treatment outcomes were included in the final multivariate analysis. The adequacy of the model fit was assessed using the Hosmer–Lemeshow goodness-of-fit test, with a P value exceeding 0.05 indicating satisfactory model fit.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boucher HW, Corey GR. 2008. Epidemiology of methicillin-resistant Staphylococcus aureus. Clin Infect Dis 46 Suppl 5:S 344–9. doi:10.1086/53359018462089 · doi ↗ · pubmed ↗
- 2Cassini A, Högberg LD, Plachouras D, Quattrocchi A, Hoxha A, Simonsen GS, Colomb-Cotinat M, Kretzschmar ME, Devleesschauwer B, Cecchini M, et al.. 2019. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: a population-level modelling analysis. Lancet Infect Dis 19:56–66. doi:10.1016/S 1473-3099(18)30605-430409683 PMC 6300481 · doi ↗ · pubmed ↗
- 3Hu F, Zhu D, Wang F, Wang M. 2018. Current status and trends of antibacterial resistance in China. Clin Infect Dis 67:S 128–S 134. doi:10.1093/cid/ciy 65730423045 · doi ↗ · pubmed ↗
- 4Turner NA, Sharma-Kuinkel BK, Maskarinec SA, Eichenberger EM, Shah PP, Carugati M, Holland TL, Fowler VG. 2019. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nat Rev Microbiol 17:203–218. doi:10.1038/s 41579-018-0147-430737488 PMC 6939889 · doi ↗ · pubmed ↗
- 5Casapao AM, Leonard SN, Davis SL, Lodise TP, Patel N, Goff DA, La Plante KL, Potoski BA, Rybak MJ. 2013. Clinical outcomes in patients with heterogeneous vancomycin-intermediate Staphylococcus aureus bloodstream infection. Antimicrob Agents Chemother 57:4252–4259. doi:10.1128/AAC.00380-1323796929 PMC 3754327 · doi ↗ · pubmed ↗
- 6Yang C-C, Sy C-L, Huang Y-C, Shie S-S, Shu J-C, Hsieh P-H, Hsiao C-H, Chen C-J. 2018. Risk factors of treatment failure and 30-day mortality in patients with bacteremia due to MRSA with reduced vancomycin susceptibility. Sci Rep 8:7868. doi:10.1038/s 41598-018-26277-929777150 PMC 5959888 · doi ↗ · pubmed ↗
- 7Lakhundi S, Zhang K. 2018. Methicillin-resistant Staphylococcus aureus: molecular characterization, evolution, and epidemiology. Clin Microbiol Rev 31:e 00020-18. doi:10.1128/CMR.00020-1830209034 PMC 6148192 · doi ↗ · pubmed ↗
- 8Fan Y-X, Chen M-T, Li N-Y, Liu X-F, Yang M-J, Chen Y-C, Liang X-Y, Wu J-F, Guo B-N, Song S-C, Zhu Y-Q, Zhang F-Y, Hang J-Q, Wu S-B, Shen B, Li H-Y, Wang Q, Luo X-M, Chen Q-G, Zhang H-F, Wang R-L, Shen L-H, Fu F-M, Song X-L, Zhang J. 2022. Sequence Type 5 (ST 5) as a possible predictor of bacterial persistence in adult patients with methicillin-resistant Staphylococcus aureus pneumonia treated with vancomycin. Microbiol Spectr 10:e 01348-22. doi:10.1128/spectrum.01348-2236094217 PMC 9603198 · doi ↗ · pubmed ↗