Inactivating conditions of therapeutic mycobacteriophages
Andrew Wiggins, Umar N. Chaudhry, Fabiana Bisaro, Addison Lueck, Alan A. Schmalstig, Graham F. Hatfull, David B. Hill, Miriam Braunstein

TL;DR
This study explores conditions that inactivate mycobacteriophages, which are being considered as treatments for drug-resistant mycobacterial infections.
Contribution
The study identifies specific conditions affecting phage activity and highlights variability among different phages.
Findings
Some conditions caused rapid inactivation of mycobacteriophages.
Phage-specific differences in sensitivity to inactivating conditions were observed.
The findings emphasize the need for individual evaluation of therapeutic phages.
Abstract
There is a need for new therapies to treat drug-resistant nontuberculous mycobacteria (NTM) disease. Bacteriophages (phages), which are viruses that infect and kill bacteria, are actively being explored as an alternative approach for treating mycobacterial diseases. Several compassionate-use cases of phage therapy for drug-resistant NTM infections exhibit favorable outcomes. To further the development of phage therapy, it is important to recognize and avoid conditions that negatively impact phage activity during phage production, storage, formulation, or treatment. Conversely, there is a need to inactivate free phages in certain preclinical phage therapy experiments. In this study, we investigated three mycobacteriophages BPsΔ33HTH-HRM10, Muddy, and ZoeJΔ45 from compassionate-use NTM treatment cases for their sensitivity to a variety of conditions that included temperature, acid pH,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
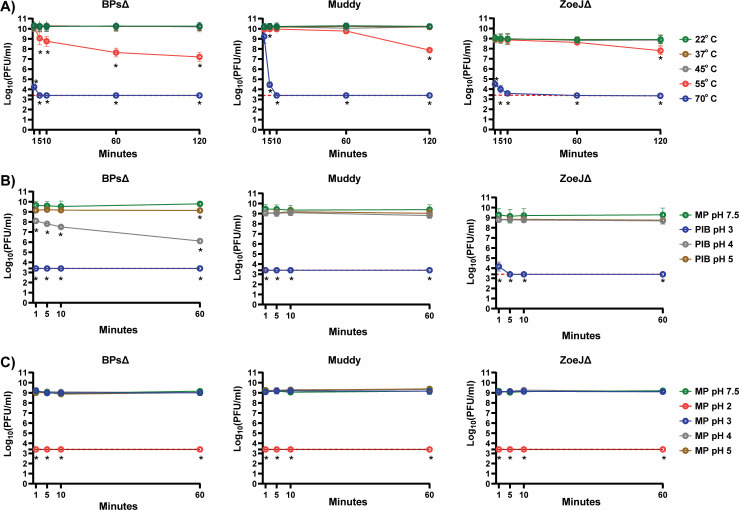 Fig 1
Fig 1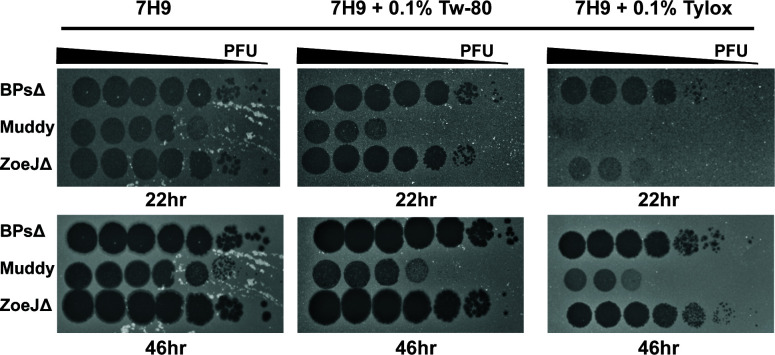 Fig 2
Fig 2| Condition | Time | BPs∆ | Muddy | ZoeJ∆ |
|---|---|---|---|---|
| 37°C | 1, 5, 10, 60, and 120 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 45°C | 1, 5, 10, 60, and 120 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 55°C | 1 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 55°C | 5 min |
| (NS, NS) | (NS, NS) |
| 55°C | 10 min |
| (NS, NS) | (NS, NS) |
| 55°C | 60 min |
| (NS, NS) | (NS, NS) |
| 55°C | 120 min |
|
|
|
| 70°C | 1 min |
|
|
|
| 70°C | 5 min |
|
|
|
| 70°C | 10 min |
|
|
|
| 70°C | 60 min |
|
|
|
| 70°C | 120 min |
|
|
|
| 90% EtOH | 1, 5, 10, and 60 min |
|
|
|
| 63% EtOH | 1 min |
|
|
|
| 63% EtOH | 5 min |
|
|
|
| 63% EtOH | 10 min |
|
|
|
| 63% EtOH | 60 min |
|
|
|
| 1:10 PIB, pH 3 | 1 min |
|
|
|
| 1:10 PIB, pH 3 | 5 min |
|
|
|
| 1:10 PIB, pH 3 | 10 min |
|
|
|
| 1:10 PIB, pH 3 | 60 min |
|
|
|
| 1:10 PIB, pH 4 | 1 min |
| (NS, NS) | (NS, NS) |
| 1:10 PIB, pH 4 | 5 min |
| (NS, NS) | (NS, NS) |
| 1:10 PIB, pH 4 | 10 min |
| (NS, NS) | (NS, NS) |
| 1:10 PIB, pH 4 | 60 min |
| (−0.53, NS) | (NS, NS) |
| 1:10 PIB, pH 5 | 1 min | (NS, NS, NS) | (NS, NS) | (NS, NS) |
| 1:10 PIB, pH 5 | 5 min | (NS, NS, NS) | (NS, NS) | (NS, NS) |
| 1:10 PIB, pH 5 | 10 min | (NS, NS, NS) | (NS, NS) | (NS, NS) |
| 1:10 PIB, pH 5 | 60 min |
| (NS, NS) | (NS, NS) |
| MP, pH 2 | 1, 5, 10, and 60 min |
|
|
|
| MP, pH 2 | 24 h |
|
|
|
| MP, pH 3 | 1, 5, 10, and 60 min | (NS, NS) | (NS, NS, NS) | (NS, NS) |
| MP, pH 4 | 1, 5, 10, and 60 min | (NS, NS) | (NS, NS) | (NS, NS) |
| MP, pH 5 | 1, 5, 10, and 60 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 2.25 mM FAS + 0.002% TA | 1 min |
|
|
|
| 2.25 mM FAS | 1 min |
|
|
|
| 0.002% TA | 3 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 2% HBE mucus | 24 h | (NS, | (NS, NS) | (NS, NS) |
| 4% HBE mucus | 24 h | ( | (NS, NS) | (NS, NS) |
| 6% HBE mucus | 24 h | ( | (NS, | (NS, NS) |
| 10 mM TCEP, pH 1.9 | 24 h |
|
|
|
| 10 mM TCEP, pH 7 | 24 h | (NS, NS) | (NS, NS) | (NS, NS) |
| 1 mM DTT, pH 5.3 | 24 h |
|
|
|
| 1 mM DTT, pH 7.0 | 24 h | (NS, NS) | (NS, NS) |
|
| 0.1% Tween 80 | 1 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 0.1% Tween 80 | 5 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 0.1% Tween 80 | 10 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 0.1% Tween 80 | 60 min | (NS, NS) | (NS, NS) | (NS, |
| 0.1% tyloxapol | 1, 5, 10, and 60 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 0.1% Triton X-100 | 1 min | (NS, NS, NS, NS) | (NS, NS, NS, NS) | (NS, NS, NS, NS) |
| 0.1% Triton X-100 | 5 min | (NS, NS, NS, NS) | (NS, NS, NS, NS) | (NS, NS, NS, NS) |
| 0.1% Triton X-100 | 10 min | (NS, NS, NS, NS) | (NS, NS, NS, NS) | (NS, NS, NS, NS) |
| 0.1% Triton X-100 | 60 min | (NS, | (NS, NS, NS, NS) | (NS, NS, NS, NS) |
| 1% Triton X-100 | 1 min | (NS, NS) | (NS, NS) | ( |
| 1% Triton X-100 | 5 min | ( | (NS, NS) | (NS, NS) |
| 1% Triton X-100 | 10 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% Triton X-100 | 60 min | (NS, NS) | (NS, NS) | (NS, |
| 10% Triton X-100 | 1, 5, 10, and 60 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% DMSO | 1 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% DMSO | 5 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% DMSO | 10 min | (NS, NS) | (NS, | (NS, NS) |
| 1% DMSO | 60 min | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% DMSO | 24 h | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% DMSO | 48 h | (NS, NS) | (NS, NS) | (NS, NS) |
| 1% DMSO | 72 h | (NS, NS) | (NS, NS) | (NS, NS) |
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —Howard Hughes Medical Institutehttp://dx.doi.org/10.13039/100000011
- —Cystic Fibrosis Foundationhttp://dx.doi.org/10.13039/100000897
- —Emily's Entourage
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —Cystic Fibrosis Foundationhttp://dx.doi.org/10.13039/100000897
- —National Science Foundationhttp://dx.doi.org/10.13039/501100008982
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —Cystic Fibrosis Foundationhttp://dx.doi.org/10.13039/100000897
- —National Institute of Diabetes and Digestive and Kidney Diseaseshttp://dx.doi.org/10.13039/100000062
- —National Institute of General Medical Scienceshttp://dx.doi.org/10.13039/100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Monoclonal and Polyclonal Antibodies Research · Mycobacterium research and diagnosis
OBSERVATION
As bacteria get harder to treat with antibiotics, the need for alternative therapies becomes more urgent. An alternative to antibiotics is the use of lytic bacteriophages (phages), which infect and kill bacteria. There are a growing number of compassionate-use cases of phages being used therapeutically to treat patients with multidrug-resistant bacterial infections, including nontuberculous mycobacteria (NTM) pulmonary infections (1, 2). For phage therapy to succeed, it is important to avoid conditions that negatively impact phage activity when producing, storing, or formulating phages since low phage titers could jeopardize treatment, as likely occurred in a clinical trial of phages for Pseudomonas aeruginosa infections (3–5). Conversely, a method for inactivating phages is useful in preclinical experiments where the bactericidal activity of phages is measured by bacterial CFU (i.e., to avoid free phages from reducing CFU during outgrowth). Here, we evaluated three siphovirus mycobacteriophages (MPs) from phage therapy cases of NTM infections, each mapping to a different genomic cluster, for their sensitivity to a variety of conditions: BPsΔ33HTH-HRM10 (BPsΔ; cluster G), Muddy (cluster AB), and ZoeJΔ45 (ZoeJΔ; cluster K) (6).
To measure phage sensitivities, phages at titers of 10^9^–10^10^ PFU/mL in MP buffer were exposed to different conditions at room temperature (22°C), unless otherwise indicated. At specific time points, phages were quantified as PFU per milliliter using an agar overlay assay with Mycobacterium smegmatis. Conditions resulting in a significant reduction in titer compared to an untreated phage control at the same time point were determined by calculating the log change (log PFU/mL_Treatment_ – log PFU/mL_Control_; see Supplemental material for Materials and Methods). All data from replicate experiments are presented in Table 1.
TEMPERATURE
Sensitivity to elevated temperature was monitored over a 120 min time course and compared to room temperature (22°C; Fig. 1A). At 37°C and 45°C, there was no significant reduction in titer for any of the phages. However, at 55°C, there was a time-dependent reduction in all the phages, with BPsΔ exhibiting the greatest sensitivity. All three phages were highly sensitive to 70°C.
(A) Sensitivity of phages to various temperatures. BPsΔ, Muddy, and ZoeJΔ were treated with a range of temperatures from 22°C to 70°C. At specific time points, samples were removed and phages quantified. The mean of two independent experiments, each with three technical replicates, is plotted. Asterisks indicate a significant reduction in log (PFU/mL) between a given treatment compared to the 22°C control at the same time point determined by one-way analysis of variance (ANOVA; P < 0.05) with Tukey’s post-test. (B) Effectiveness of phage inactivating buffer (PIB) at various pH. BPsΔ, Muddy, and ZoeJΔ were treated at a 1:10 ratio with PIB at pH 3, 4, and 5 and compared to MP Buffer pH 7.5. At specific time points, phages were quantified. The mean of two to three independent experiments, each with three technical replicates, is plotted. Asterisks indicate a significant reduction in log (PFU/mL) between a given PIB pH time point compared to the MP buffer pH 7.5 control at the same time point determined by one-way ANOVA (P < 0.05) with Tukey’s post-test. (C) Sensitivity of phages to various pH in MP buffer. BPsΔ, Muddy, and ZoeJΔ were incubated in MP buffer at pH 2, 3, 4, and 5 and compared to MP buffer, pH 7.5. At specific time points, samples were removed and phage quantified. The mean of two to three independent experiments, each with three technical replicates, was plotted. Asterisks indicate a significant reduction in log (PFU/mL) between a given pH time point compared to the MP buffer pH 7.5 control at the same time point determined by one-way ANOVA (P < 0.05) with Tukey’s post-test. The red dashed line indicates the limit of detection. Error bars indicate standard deviation. Samples with zero PFU recovered are plotted at the level of detection.
ETHANOL
Sensitivity to 63% and 90% ethanol, a commonly used disinfectant, was tested (Table 1). All three phages were highly sensitive to 90% ethanol with undetectable levels at 1 min. Ethanol at 63% also had a rapid, although incomplete, effect with BPsΔ again exhibiting the greatest sensitivity.
PHAGE INACTIVATION BUFFER AND ACIDIC pH
When measuring phage bactericidal activity by reductions in CFU, it is advantageous to inactivate free phage before CFU plating. We previously reported the use of a 10 min treatment with a phage inactivating buffer (PIB; 940 mM citric acid, 10 mM potassium chloride, 135 mM NaCl, pH 3.0) to inactivate BPsΔ, Muddy, and ZoeJΔ (7–9). Importantly, the host bacteria of these phages Mycobacterium abscessus and M. smegmatis are not inhibited by PIB pH 3, 10 min (8) (Fig. S1). To determine the time-dependence of inactivation, we performed a PIB time course and observed a rapid reduction of the phages to undetectable levels: by 1 min for BPsΔ and Muddy and 5 min for ZoeJΔ (Fig. 1B). The effect of pH may be an important inactivating component of PIB (10, 11). Therefore, we investigated the adjustment of PIB to either pH 4 or pH 5. Compared to PIB pH 3, PIB pH 4 and 5 had no effect on Muddy or ZoeJΔ, and PIB pH 4 and 5 was less effective at reducing BPsΔ (Fig. 1B). BPsΔ was again more sensitive than the other phages. We also studied the impact of acidic pH in the MP buffer. All the phages were highly sensitive to MP buffer pH 2 (i.e., undetectable at 1 min) and resistant to MP pH 3, 4, and 5 (Fig. 1C). Thus, PIB pH 3 fully inactivates, but MP pH 3 does not, indicating that the pH of PIB is important but not the sole factor responsible for phage inhibition.
FERROUS AMMONIUM SULFATE ± TANNIC ACID
Ferrous ammonium sulfate (FAS) ± tannic acid (TA) is another reported phage inactivating treatment (12–14) . After 1 min incubation in 2.25 mM FAS with 0.002% TA, the titer of all three phages was below the level of detection (Table 1). One-minute treatment with 2.25 mM FAS lacking TA also significantly reduced phage titers, with Muddy being undetectable and BPsΔ and ZoeJΔ exhibiting a 4–5 log reduction. Treatment with 0.002% TA for 3 min had no effect (Table 1).
MUCUS
Several patients receiving phage therapy for pulmonary NTM disease have muco-obstructive lung diseases (2). Studies of mucus effects on other phages reveal inhibitory or beneficial effects (15–17). This led us to test the effect of mucus harvested from cultured human bronchial epithelial cells (18, 19). Phages were incubated for 24 h with 2%, 4%, or 6% mucus, which are concentrations that model healthy, moderate, and severe airway disease mucus (20, 21). Across the concentrations, all three phages exhibited either modest (≤0.7 log) or no significant reduction in titer. This finding of little to no effect of mucus supports the therapeutic use of these phages in people with muco-obstructive diseases.
REDUCING AGENTS
Reducing agents that disrupt disulfide bonds degrade the mucin network of mucus. There is interest in developing reducing agents into mucolytics for muco-obstructive diseases (21, 22). At concentrations that reduce the viscosity of mucus, we tested reducing agents tris(2-carboxyethyl)phosphine (TCEP) and dithiothreitol (DTT) on phage activity (23). Because TCEP and DTT solutions are acidic, we tested each agent as an unbuffered solution and at pH 7. Without buffering, both 10 mM TCEP pH 1.9 and 1 mM DTT pH 5.3 for 24 h significantly reduced the level of all three phages. However, both agents at pH 7 had no effect, apart from a modest reduction in ZoeJΔ PFU with 1mM DTT (Table 1). Because mucus has a high buffering capacity, which limits pH effects in vivo (21), these results suggest that concurrent treatment with reducing mucolytics will not inhibit these phages.
DETERGENTS
Sensitivity to detergents encountered in mycobacterial experiments (Tween-80, tyloxapol, Triton X-100) was investigated. Tween-80 and tyloxapol are included in mycobacterial liquid media to promote dispersed growth. Triton X-100 is used to lyse cultured eukaryotic cells infected with mycobacteria (8). All three phages were resistant to 60 min incubation with 0.1% Tween-80, 0.1% tyloxapol, or 0.1%, 1%, or 10% Triton X-100 (Table 1). This resistance was surprising since Tween-80 is reported to prevent MP D29 infection (24). We wondered if the detergents might not inhibit the phage directly but, rather, prevent phage infection through effects on the mycobacterial host. To test this idea, immediately prior to use, 0.1% Tween-80, 0.1% tyloxapol, or no detergent was added to the M. smegmatis culture used in the overlay for phage quantification (Fig. 2). For BPsΔ and ZoeJΔ, Tween-80 or tyloxapol in the culture did not affect PFU number, although tyloxapol delayed the time for ZoeJΔ plaques to appear. In contrast, inclusion of Tween-80 or tyloxapol in the culture inhibited Muddy PFU. These results demonstrate that the activity of some, but not all, phages is impacted by detergent-specific effects on mycobacteria.
Sensitivity of phages to inclusion of detergent in the M. smegmatis culture used in agar overlays. BPsΔ, Muddy, and ZoeJΔ 10-fold serial dilutions spotted onto agar overlays prepared with M. smegmatis resuspended in Middlebrook 7H9 media supplemented with 0.5% glycerol and 0.2% glucose (no detergent) or the same Middlebrook 7H9 media with freshly added 0.1% Tween-80 (Tw-80) or 0.1% tyloxapol (Tylox). Plates were incubated at 37°C for 22 and 46 h. A representative image from two experiments is shown. All images were taken with the same exposure time of 0.09 s using a Bio-RAD GelDOC Go.
DIMETHYL SULFOXIDE
Many antimycobacterial drugs are dissolved in dimethyl sulfoxide (DMSO), such that 1% DMSO is likely to be included when phage-antibiotic combinations are tested on mycobacteria. DMSO may also be used as a protectant in long-term phage storage (25). Up to 72 h, all three phages were resistant to 1% DMSO (Table 1).
In conclusion, in studying MPs from phage therapy cases (2), we discovered several conditions that result in dramatic and rapid reductions in phage activity. We also observed differences in sensitivities between phages, emphasizing the need for such studies. Although currently, MPs are most commonly administered intravenously, there are cases where pulmonary delivery (i.e., nebulization) may be used (2). For such cases, the potential impact of lung surfactant is another variable to consider. Future studies will address interactions between phages and lung surfactant. Taken together, the results presented in this report provide valuable information to support further evaluation of these phages as therapeutics for NTM disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Strathdee SA, Hatfull GF, Mutalik VK, Schooley RT. 2023. Phage therapy: from biological mechanisms to future directions. Cell 186:17–31. doi:10.1016/j.cell.2022.11.01736608652 PMC 9827498 · doi ↗ · pubmed ↗
- 2Dedrick RM, Smith BE, Cristinziano M, Freeman KG, Jacobs-Sera D, Belessis Y, Whitney Brown A, Cohen KA, Davidson RM, van Duin D, et al.. 2023. Phage therapy of Mycobacterium infections: compassionate use of phages in 20 patients with drug-resistant mycobacterial disease. Clin Infect Dis 76:103–112. doi:10.1093/cid/ciac 45335676823 PMC 9825826 · doi ↗ · pubmed ↗
- 3Blazanin M, Lam WT, Vasen E, Chan BK, Turner PE. 2022. Decay and damage of therapeutic phage OMKO 1 by environmental stressors. P Lo S One 17:e 0263887. doi:10.1371/journal.pone.026388735196336 PMC 8865689 · doi ↗ · pubmed ↗
- 4Jończyk-Matysiak E, Łodej N, Kula D, Owczarek B, Orwat F, Międzybrodzki R, Neuberg J, Bagińska N, Weber-Dąbrowska B, Górski A. 2019. Factors determining phage stability/activity: challenges in practical phage application. Expert Rev Anti Infect Ther 17:583–606. doi:10.1080/14787210.2019.164612631322022 · doi ↗ · pubmed ↗
- 5Jault P, Leclerc T, Jennes S, Pirnay JP, Que YA, Resch G, Rousseau AF, Ravat F, Carsin H, Le Floch R, Schaal JV, Soler C, Fevre C, Arnaud I, Bretaudeau L, Gabard J. 2019. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (Phago Burn): a randomised, controlled, double-blind phase 1/2 trial. Lancet Infect Dis 19:35–45. doi:10.1016/S 1473-3099(18)30482-130292481 · doi ↗ · pubmed ↗
- 6Dedrick RM, Guerrero-Bustamante CA, Garlena RA, Russell DA, Ford K, Harris K, Gilmour KC, Soothill J, Jacobs-Sera D, Schooley RT, Hatfull GF, Spencer H. 2019. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat Med 25:730–733. doi:10.1038/s 41591-019-0437-z 31068712 PMC 6557439 · doi ↗ · pubmed ↗
- 7Brindley MA, Maury W. 2008. Equine infectious anemia virus entry occurs through clathrin-mediated endocytosis. J Virol 82:1628–1637. doi:10.1128/JVI.01754-0718057237 PMC 2258727 · doi ↗ · pubmed ↗
- 8Schmalstig AA, Wiggins A, Badillo D, Wetzel KS, Hatfull GF, Braunstein M. 2024. Bacteriophage infection and killing of intracellular Mycobacterium abscessus m Bio 15:e 02924-23. doi:10.1128/mbio.02924-2338059609 PMC 10790704 · doi ↗ · pubmed ↗