The chitinolytic enzymes from the entomopathogenic fungus Metarhizium anisopliae: GH family distribution, domain architecture, and protein regulation
Qusai Al Abdallah, Kholoud M. Alananbeh, Nehaya Al-Karablieh, Salah-Eddin Araj

TL;DR
This paper reviews the chitin-degrading enzymes in the fungus Metarhizium anisopliae, focusing on their classification, structure, and potential use in pest control.
Contribution
The paper consolidates current knowledge on M. anisopliae chitinolytic enzymes and their regulatory mechanisms.
Findings
M. anisopliae has 21 chitinases classified into four GH18 subgroups and produces two GH20 and two GH3 NAGases.
The domain architectures of M. anisopliae enzymes are similar to Trichoderma spp. but with evolutionary distinctions.
Chitinases may be used as biopesticides against fungal pathogens and pests.
Abstract
Metarhizium anisopliae is an entomopathogenic fungus that is widely used in the biological control of agricultural pests. During host infection, M. anisopliae secretes an arsenal of hydrolytic enzymes such as proteases and chitinases that allow for cuticle penetration and host colonization. The degradation of chitin into N-acetylglucosamine (GlcNAc) monomers is carried out by chitinases and β-N-acetylglucosaminidases (NAGases). Chitin-degrading enzymes of M. anisopliae have been the subject of extensive research; however, these studies have not been consolidated into a comprehensive review. This review highlights our current knowledge of the chitinolytic enzymes from M. anisopliae, emphasizing the classification of these enzymes based on their mode of action and domain architecture. M. anisopliae possesses a set of 21 chitinases that are classified into four glycoside hydrolase family…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
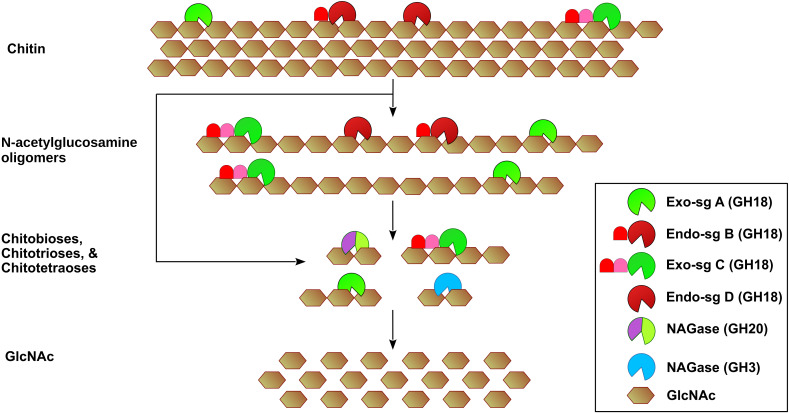 Figure 1
Figure 1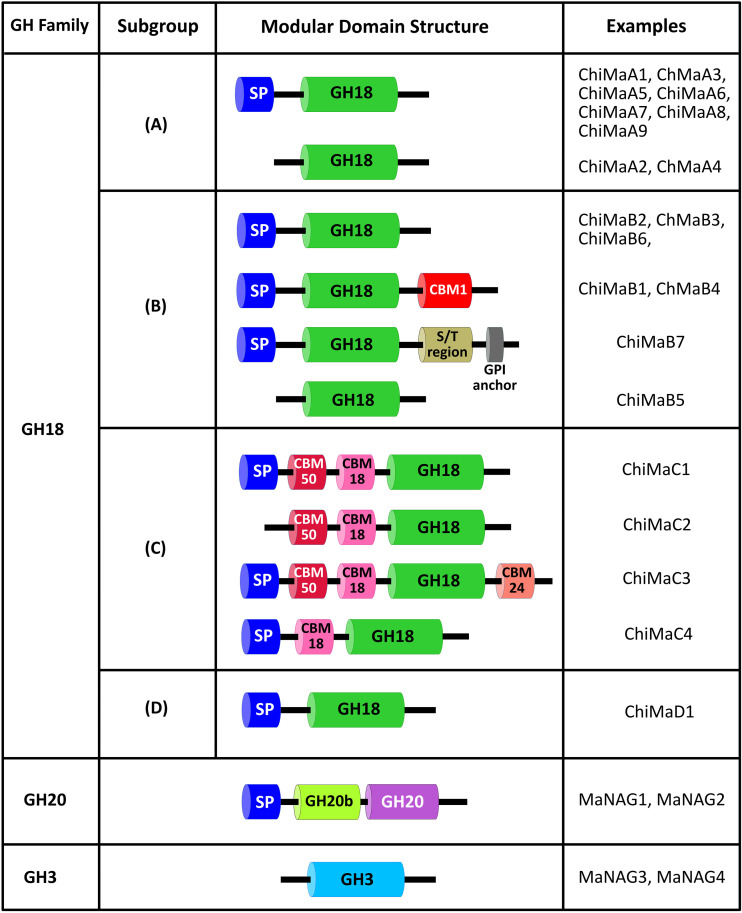 Figure 2
Figure 2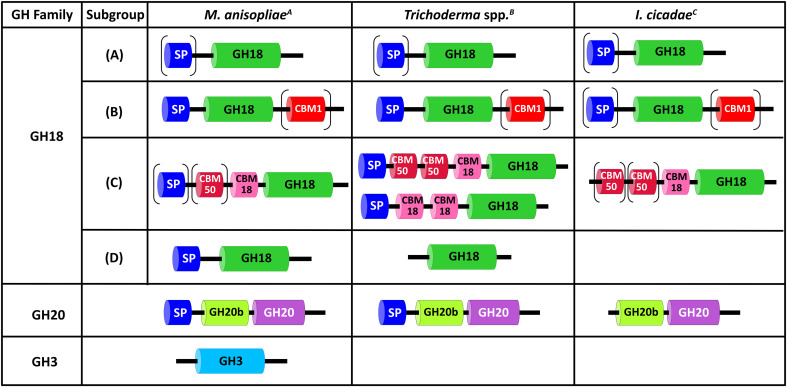 Figure 3
Figure 3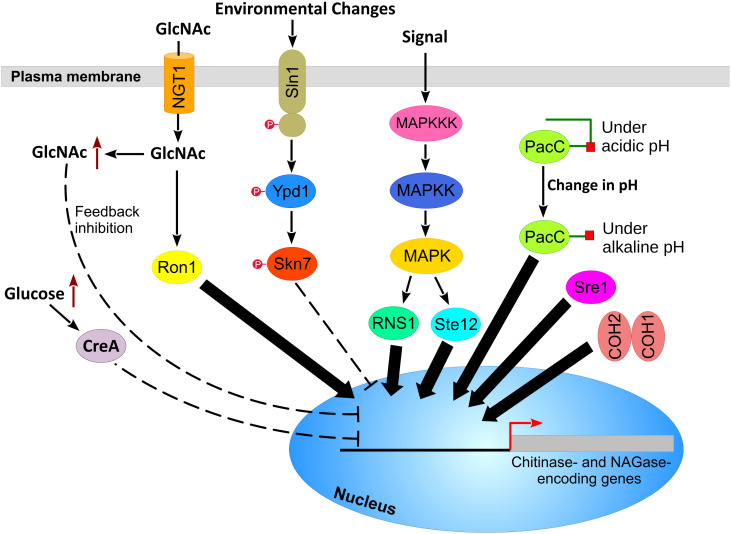 Figure 4
Figure 4| Enzyme (gene name) | GH18 family subgroup | Molecular weight (kDa) | Domain organization | Activity | Virulence impact |
|---|---|---|---|---|---|
| CHIT42 ( | A | 42 | SP – GH18 | Endo | • Overexpression did not alter virulence against |
| CHI2 ( | B | 42 | SP – GH18 – CBM1 | Endo | • Deletion mutant exhibited reduced virulence against cotton stainer bug ( |
| CHI30 ( | B | 30 | SP – GH18 | Endo & Exo | • Deletion mutant exhibited reduced fungal virulence against cotton stainer bug. |
| ChiMaD1 | D | 36.6 | SP – GH18 | Endo | • Deletion mutant exhibited higher virulence rates against the cattle tick ( |
| GH family | GH18 subgroup | Protein | Protein accession number | Published locus tag | Notes |
|---|---|---|---|---|---|
| GH18 | A | ChiMaA1 | MANI_007345 | Also designated CHIT42 (gene: | |
| ChiMaA2 | MANI_003521 | ||||
| ChiMaA3 | MANI_022967 | ||||
| ChiMaA4 | MANI_012760 | ||||
| ChiMaA5 | MANI_005739 | ||||
| ChiMaA6 | MANI_029841 | ||||
| ChiMaA7 | MANI_000730 | ||||
| ChiMaA8 | MANI_021851 | ||||
| ChiMaA9 | MANI_006991 | ||||
| B | ChiMaB1 | MANI_002801 | Also designated CHI2 (gene: | ||
| ChiMaB2 | MANI_115994 A | Also designated CHI30 (gene: | |||
| ChiMaB3 | MANI_026679 | ||||
| ChiMaB4 | MANI_004417 | ||||
| ChiMaB5 | MANI_021602 | ||||
| ChiMaB6 | MANI_018482 | ||||
| ChiMaB7 | MANI_012994 | ||||
| C | ChiMaC1 | MANI_023684 | |||
| ChiMaC2 | MANI_120160 A | ||||
| ChiMaC3 | MANI_115484 A | ||||
| ChiMaC4 | MANI_116045 A | ||||
| D | ChiMaD1 | MANI_018860 | |||
| GH20 | N/A B | MaNAG1 | MANI_010908 | ||
| N/A B | MaNAG2 | MANI_029504 | |||
| GH3 | N/A B | MaNAG3 | MANI_122030 | ||
| N/A B | MaNAG4 | Not found | MANI_128875 |
| Enzyme | Entomopathogenic | Mycoparasitic | Saprophytic | |||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
| |
| Chi-sg A | 9 | 8 | 7 | 8 | 7 | 7 | 8 | 7 |
| Chi-sg B | 7 | 7 | 5 | 4 | 7 | 11 | 11 | 7 |
| Chi-sg C | 4 | 6 | 2 | 8 | 3 | 9 | 15 | 4 |
| Chi-sg D | 1 | ND | ND | ND | ND | 1 | 1 | 1 |
| Total Chitinases | 21 | 21 | 14 | 20 | 17 | 28 | 35 | 19 |
| GH20 NAGase | 2 | 2 | 2 | 5 | 1 | 2 | 2 | 2 |
| GH3 NAGase | 2 | ND | ND | ND | ND | ND | ND | ND |
| Total | 25 | 23 | 16 | 25 | 18 | 29 | 36 | 21 |
| Regulator/Transporter | Fungus | Protein accession number | Accession number of | Similarity (%) |
|---|---|---|---|---|
| Sln1 |
| 71.1 | ||
| Ypd1 |
| 98.7 | ||
| Skn7 |
| 95.8 | ||
| PacC |
| 99.7 | ||
| Ron1 (syn. PhoG or Vib1) |
| 44.0 | ||
| Sre1 |
| * | 44.6 | |
| NGT1 |
| 42.0 | ||
| CreA |
| 87.8 | ||
| Fus3 MAPK |
| 100 | ||
| Hog1 MAPK |
| 100 | ||
| Slt2 MAPK |
| 99.5 | ||
| Ime2 MAPK |
| 99.2 | ||
| Ste7 MAPK |
| 100 | ||
| Pbs2 MAPK |
| 97.5 | ||
| Mkk MAPK |
| 98.9 | ||
| Ste11 MAPK |
| 99.6 | ||
| Bck1 MAPK |
| 99.4 | ||
| Ssk2 MAPK |
| 98.9 | ||
| RNS1 |
| 97.6 | ||
| Ste12 |
| 85.9 | ||
| COH1 |
| 97.7 | ||
| COH2 |
| 92.0 |
| Enzyme locus in | Accession number of | Similarity % | Regulators of this enzyme A |
|---|---|---|---|
| Subgroup A chitinases | |||
| MAA_02685 ( | 99.0 | PacC ↑ | |
| MAA_04700 ( | 99.0 | PacC ↑ | |
| MAA_03460 ( | 95.8 | MAPK ↓, | |
| MAA_08154 ( | 93.0 | Skn7 ↓, | |
| MAA_10256 ( | 97.1 | PacC ↑ | |
| Subgroup B chitinases | |||
| MAA_09018 ( | 98.3 | PacC ↑ | |
| MAA_01212 ( | 82.1 | RNS1 ↑ | |
| MAA_02744 ( | 94.7 | MAPK ↓ | |
| MAA_00983 ( | 85.6 | PacC ↑ | |
| MAA_00415 ( | 74.0 | PacC ↑ | |
| MAA_09091 ( | 87.7 | PacC ↑ | |
| Subgroup C chitinases | |||
| MAA_00157 ( | 48.5 | MAPK ↑, | |
| MAA_10509 ( | 69.3 | MAPK ↑ | |
| Subgroup D chitinases | |||
| MAA_08315 ( | 92.2 | MAPK ↓ | |
| GH20 NAGase | |||
| MAA_10080 ( | 97.6 | PacC↑, | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStudies on Chitinases and Chitosanases · Entomopathogenic Microorganisms in Pest Control · Fungal and yeast genetics research
Introduction
1
Entomopathogenic fungi play an important role in the biological control of agricultural pests. They can cause diseases in insect populations and thereby improve crop yields. In addition, these fungi inhabit a wide range of habitats, enabling effective control of various pest species under field conditions (Moazami, 2011; Kumar et al., 2019).
Entomopathogenic fungi infect their insect hosts through a process that is broadly comparable to that used by plant-pathogenic fungi (reviewed by Ma et al., 2024). Infection begins when conidia attach to the insect cuticle and germinate to produce a germ tube. The germ tube tip then differentiates to form a specialized infection structure, designated appressorium, which secretes cuticle-degrading enzymes, including chitinases, proteases, and lipases, allowing for fungal penetration into the hemocoel (Khachatourians and Qazi, 2008; Santi et al., 2010a; Chethana et al., 2021; Gebremariam et al., 2022). Following successful invasion, the fungus proliferates within the host, producing mycotoxins and hydrolytic enzymes while assimilating host-derived nutrients, ultimately leading to insect death (Butt et al., 2016). Although several thousand entomopathogenic fungal species are thought to exist, only a small proportion has been studied in detail (Blackwell, 2011; Santos et al., 2022). Among these, Beauveria bassiana and Metarhizium anisopliae remain the most extensively investigated species (Santos et al., 2022).
M. anisopliae is a saprophytic fungus that thrives on decaying organic matter and inhabits the soil of a wide range of habitats (Moonjely and Bidochka, 2019; Tang et al., 2025). The taxonomy of M. anisopliae has been recently addressed by few articles (Bischoff et al., 2009; Kobmoo et al., 2024; Nishi, 2024). In addition, it is a generalist entomopathogenic fungus that infects a broad spectrum of host insects (Bidochka et al., 1998; Schrank and Vainstein, 2010; St. Leger and Wang, 2020; de Miranda et al., 2024). During different stages of the disease cycle, M. anisopliae produces specialized cells such as appressorium, blastospores, and hyphae, which are used for host infection. The disease cycle of M. anisopliae is illustrated in Figure 2 in Schrank and Vainstein (2010) and Figure 1 in Li and Xia (2022).
Schematic diagram for the hydrolysis of chitin by M. anisopliae enzymes. The endo-acting chitinases of subgroups B and D hydrolyze glycosidic bonds randomly, producing soluble short chains of GlcNAc oligomers (5–10 GlcNAc units). The subgroups A and C exo-chitinases act on chitin terminal ends, releasing chitobioses (containing 2 GlcNAc molecules), chitotrioses (containing 3 GlcNAc molecules), and chitotetraoses (containing 4 GlcNAc molecules). The hydrolysis products are further hydrolyzed to GlcNAc by exo-chitinases and β-N-acetylglucosaminidases (NAGases). Although subgroup A chitinases are predicted to exhibit exo-acting activity, CHIT42 displays an endo-acting activity. CHI30, which belongs to the endo-acting subgroup B, exhibits a dual endo- and exo-chitinase activity.
Domain architecture of chitinases and NAGase from M. anisopliae. SP, signal peptide; GH18, GH18 catalytic domain; GH20, GH20 catalytic domain; GH3, GH3 catalytic domain; GH20b, non-catalytic GH20b domain; CBM, carbohydrate-binding module; S/T region, Ser/Thr linker region; GPI anchor, GPI for anchoring protein to the plasma membrane.
The first genome sequence of M. anisopliae was published using the isolate ARSEF23 as a reference strain; however, this isolate was reclassified as Metarhizium robertsii (Gao et al., 2011). The complete genome sequence of M. anisopliae was repeated using the isolates E6 and Ma69 (Pattemore et al., 2014; Staats et al., 2014; Iwanicki et al., 2022). The genome size of M. anisopliae is about 38.5 Mb with 127 proteins that have no homologs in M. robertsii or M. acridum genome. The fungus produces 1620 secreted proteins, including an arsenal of proteases and chitinases that allow for cuticle penetration and host colonization (Pattemore et al., 2014; Staats et al., 2014). M. anisopliae chitinases are key players in fungal virulence against different stages of insect pests.
To understand their activity and specificity, chitinases from the entomopathogenic fungus M. anisopliae have been the subject of extensive studies that span biochemical characterization, structural analysis, and regulation of gene expression. However, comprehensive reviews that highlight these findings are lacking. The main objectives of this review are to (1) consolidate our current knowledge on chitin-hydrolyzing enzymes from M. anisopliae, including their glycoside hydrolase (GH) classification and domain architecture, and (2) identify conserved molecular mechanisms underlying chitinase gene expression using studies from other entomopathogenic fungi.
Chitin as a structural barrier in insect hosts
2
Insects have an external complex structure called the exoskeleton. The insect exoskeleton, which is also called the cuticle, is composed of two non-living layers: the epicuticle and the procuticle (Chen et al., 2008; Andersen, 2009; Doucet and Retnakaran, 2016; Vittori, 2024). The cuticle serves essential functions in insect support and protection. For example, it serves as an external framework that supports the body of insects and determines their shape. In addition, muscles are attached to the cuticle and thus aid in insects’ mobility. Furthermore, the exoskeleton operates as a barrier protecting insects from external environmental factors and physical damage (Gorb, 2001; Vincent, 2001; Hamid et al., 2013). The exoskeleton of insects generally contains 30% to 45% protein and 25% to 40% lipids. The chitin content in the exoskeleton varies among the species, ranging from 5 to 15% (Izadi et al., 2025).
Chitin is a linear polysaccharide of N-acetylglucosamine (abbr.: GlcNAc) repeating units that are linked by β-1,4-glycosidic bonds. Structurally, GlcNAc is similar to glucose but has an acetamide group at the C2 position instead of the hydroxyl group (Jones et al., 2020). Chitin exists in three allomorphic forms: α-chitin, β-chitin, and γ-chitin, which differ in chain orientation and hydrogen bonding patterns, resulting in distinct mechanical properties. α-chitin consists of antiparallel chains stabilized by intra- and intermolecular hydrogen bonds, resulting in greater stiffness. In contrast, the structure of β-chitin consists of parallel chains connected via weak intermolecular hydrogen bonds. β-chitin exhibits a reduced stiffness that provides more flexibility compared to α-chitin. Both α-chitin and β-chitin are commonly found in arthropods. The structure of γ-chitin exhibits a mixture of α- and β-chitin, and it is mainly found in fungi, yeasts, and insect cocoons. For a schematic representation of the three allomorphic forms of chitin, refer to Figure 1 in Mei et al. (2024) and Figure 4 in Kumirska et al. (2011). Partial deacetylation of chitin results in the production of chitosan, which is a linear polymer of D-glucosamine and N-acetyl-D-glucosamine (Al-Nasrawi et al., 2024; Abdulhameed et al., 2025).
Comparison of the domain architecture of the chitinolytic enzymes from M. anisopliae, Trichoderma spp., and Isaria cicadae. SP: signal peptide, GH18: GH18 catalytic domain, GH20: GH20 catalytic domain, GH3: GH3 catalytic domain, GH20b: non-catalytic GH20b domain, CBM: carbohydrate-binding module. Data were collected from A (Junges et al., 2014; Oliveira et al., 2018), B (Seidl et al., 2005; López-Mondéjar et al., 2009; Häkkinen et al., 2012; Seidl-Seiboth et al., 2014), and C (Peng et al., 2020).
Hydrolysis of chitin by chitinases and β-N-acetylglucosaminidases from M. anisopliae
3
The degradation of chitin by fungal chitinolytic enzymes is a consecutive and coordinated multistep process that involves exo-chitinases, endo-chitinases, lytic polysaccharide monooxygenases (LPMOs), and β-N-acetylglucosaminidases (NAGases) (Busk and Lange, 2015). Chitinases also exhibit enzymatic activity against chitosan; however, such activity is strongly dependent on the degree of acetylation (Hartl et al., 2012). It should be mentioned that LPMOs will not be covered in this review because of a lack of sufficient data. In this review, the terms chitinolytic enzymes, chitin-degrading enzymes, and chitin-hydrolyzing enzymes are used interchangeably to refer to chitinases and NAGases.
The classification of chitinases can be based on either the mode of action or amino acid sequence similarities (Henrissat and Davies, 1997; Poria et al., 2021). The Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (IUBMB) classifies chitinases based on their mode of action into endo-acting chitinases and exo-acting chitinases. Endo-acting chitinases (EC 3.2.1.14) cleave randomly along chitin internal chains, producing soluble N-acetylglucosamine oligomers (Poria et al., 2021). Exo-acting chitinases act on chitin terminal ends at the reducing end (EC 3.2.1.201) and the non-reducing end (EC 3.2.1.200), releasing chitobioses (contain two GlcNAc molecules), chitotrioses (contain three GlcNAc molecules), and chitotetraoses (contain four GlcNAc molecules). Chitin hydrolysis products are further cleaved by exo-chitinases and β-N-acetylglucosaminidases (NAGases) (EC 3.2.1.52) (Hartl et al., 2012; Muschiol et al., 2020; Jiang et al., 2025) (Figure 1).
However, the IUBMB classification does not provide insight into enzymes structural features or predict their 3D structures (Henrissat and Davies, 1997). The Carbohydrate-Active Enzymes database (CAZy, https://www.cazy.org/) has, therefore, developed a classification system based on amino acid sequence similarity, enabling the grouping of enzymes with conserved structures and providing insights into protein structure and mechanism (Henrissat and Davies, 1997).
The class glycoside hydrolases (GHs) contains enzymes with EC 3.2.1.- activity that hydrolyze glycosidic bonds (Henrissat and Davies, 1997; Talamantes et al., 2016). Under this classification system, endo- and exo-chitinases are placed in GH families 18 and 19, whereas β-N-acetylglucosaminidases are distributed in GH families 3, 20, and 84 (Oyeleye and Normi, 2018). Fungal chitinases are exclusively found in GH family 18. The only exception is NbChiA (a chitinase isolated from the fungus Nosema bombycis), which is the only known fungal chitinase that belongs to GH family 19 (Han et al., 2016).
M. anisopliae possesses a complex arsenal of chitin-degrading enzymes; however, only four chitinases have been functionally characterized (Table 1). CHIT42 is the first characterized chitinase from M. anisopliae. It is an endo-acting chitinase that is composed of 423 amino acid and has a molecular mass of 42 kDa. The enzyme contains a signal peptide for protein secretion (Bogo et al., 1998; Baratto et al., 2003). Despite its clear biochemical activity, constitutive expression of the chit1 gene under the control of the Aspergillus nidulans gpdA promoter did not elevate virulence toward Manduca sexta caterpillars, suggesting that increased chitinase activity alone is insufficient to enhance pathogenicity (Screen et al., 2001). CHI2 is another chitinase that was characterized in M. anisopliae. The enzyme, which exhibits endochitinase activity, is composed of 419 amino acids and has a molecular mass of 42 kDa (Baratto et al., 2006; Boldo et al., 2009). In contrast to CHIT42, CHI2 appears to play a more direct role in virulence. Over-expression of the chi2 gene in M. anisopliae results in a strain with elevated virulence against the cotton stainer bug (Dysdercus peruvianus). Deletion of the chi2 gene in M. anisopliae reduced fungal virulence by 20% compared to the wild type strain (Boldo et al., 2009). These observations are consistent with broader trends in Metarhizium and Beauveria, where virulence differences often reflect variation in expression and deployment of secreted enzymes rather than their mere presence (Ortiz-Urquiza and Keyhani, 2013). Interestingly, two proteins are produced from the chi2 gene via alternative splicing of the chi2 transcripts (Boldo et al., 2010). CHIT30 is a 30 kDa chitinase from M. anisopliae that is encoded by the chi3 gene and exhibits dual endo- and exochitinase activities (Pinto et al., 1997; da Silva et al., 2005). Deletion of the chi3 gene reduced fungal virulence against cotton stainer bug D. peruvianus, suggesting a major role in cuticle degradation and host penetration. Interestingly, the secretion of CHIT30 is also induced by heat shock at 40°C, suggesting another role of CHIT30 in heat-shock adaptation (Staats et al., 2013). This multifunctional role aligns with observations in other hypocrealean fungi, including Trichoderma spp., where secreted chitinases contribute both to host interaction and environmental adaptation (Kubicek et al., 2011; Mukherjee et al., 2013). ChiMaD1 is the fourth chitinase to be characterized from M. anisopliae. The molecular characterization and virulence impact of ChMaD1 will be discussed below.
GH18 family proteins
3.1
The glycoside hydrolase family 18 (GH18) includes fungal chitinases as well as chitinases from other organisms, such as bacteria, animals, and plants (reviewed in detail by Chen et al., 2020). In general, chitinases of the GH18 family possess a conserved catalytic domain that adopts a (β/α)8 TIM (triosephosphate isomerase) barrel fold. Additionally, the substrate-binding site of GH18 chitinases typically accommodates at least five sugar units, where cleavage occurs between the third and fourth units. Enzymes of the GH18 family are further subgrouped into class III and class V chitinases. These two classes differ in their mode of action and substrate binding groove, which influences substrate specificity and enzymatic function (Seidl, 2008; Hartl et al., 2012).
The domain architecture of GH family 18 chitinases comprises a combination of four functional regions: (1) a signal peptide that directs secretion of the protein outside the cell, (2) a GH18 catalytic domain that cleaves β-1,4 glycosidic bonds between GlcNAc units, (3) a carbohydrate (or chitin)-binding module (CBM) that facilitates binding and hydrolysis of insoluble chitin, and (4) a serine (Ser)/threonine (Thr)-rich linker. The biological function of the linker is not fully understood; however, it has been proposed that the Ser/Thr-rich linker enhances the enzymatic activity against insoluble substrates and protects chitinases from proteolytic degradation, particularly by extracellular proteases (González et al., 2012; Huang et al., 2012). In addition, the Ser/Thr-rich linker may contribute to proper protein folding and has been implicated in glycosylphosphatidylinositol (GPI) anchoring of chitinases to the plasma membrane (Junges et al., 2014).
Based on the comparative analysis of 18 chitinases from the filamentous fungus Trichoderma reesei, fungal GH18 chitinases have been classified into three subgroups designated A, B, and C (Seidl et al., 2005). This classification system is based on a combination of protein domain architecture, enzymatic mode of action (endo- or exo-acting), and the structural and physiochemical properties of the substrate-binding cleft (Hartl et al., 2012). Subgroups A and B show homology to class V and class III, respectively, of the GH18 family. In contrast, subgroup C comprises a novel class of high molecular weight chitinases. Interestingly, one chitinase, designated Chi18-15, did not exhibit sequence similarity to any of the three fungal subgroups (Seidl et al., 2005). Nevertheless, Seidl et al. (2005) did not assign Chi18–15 to a distinct subgroup.
In silico analysis of the M. anisopliae genome revealed the presence of 21 putative chitinases, including the three characterized chitinases (CHIT42, CHI2, and CHI30). Based on sequence homology, these enzymes were categorized into the three fungal subgroups: A, B, and C (Junges et al., 2014). The distribution of enzymes across the three subgroups was as follows: nine enzymes were grouped in subgroup A and were designated ChiMaA1 to ChiMaA9, seven enzymes were grouped in subgroup B and were designated ChiMaB1 to ChiMaB7, and four enzymes were grouped in subgroup C and were designated ChiMaC1 to ChiMaC4. In addition, Junges et al. (2014) assigned a chitinase that is homologous to the T. reesei Chi18–15 to a novel subgroup D and was designated ChiMaD1 (Table 2). As anticipated, the analysis of M. anisopliae genome did not identify any GH19 chitinases.
Fungal subgroup A chitinases
3.1.1
Fungal subgroup A chitinases constitute the predominant subgroup of fungal GH18 chitinases and are conserved across all fungal genomes. These enzymes typically exhibit a low molecular weight ranging from 40–50 kDa. The substrate-binding cleft of subgroup A enzymes is deep and narrow. The majority of subgroup A chitinases are composed of an N-terminal signal peptide followed by a GH18 catalytic domain. Despite this conserved domain architecture, some subgroup A chitinases lack the signal peptide and are therefore predicted to remain intracellularly or to be secreted via unconventional protein secretion pathways (Seidl, 2008; Ding et al., 2012). In M. anisopliae, seven subgroup A chitinases exhibit an N-terminal signal peptide for secretion outside the cell, whereas the remaining two enzymes (ChiMaA2 and ChiMaA4) lack a signal peptide (Junges et al., 2014). The well-characterized CHIT42 (ChiMaA1) of M. anisopliae belongs to subgroup A and contains both an N-terminal signal peptide and a GH18 catalytic domain consistent with the subgroup A architecture (Figure 2).
Fungal subgroup B chitinases
3.1.2
The majority of subgroup B chitinases are low molecular weight enzymes, typically ranging from 30–50 kDa. In contrast to subgroup A, the substrate-binding cleft is shallow and open. The domain architecture of subgroup B chitinases consists of an N-terminal signal peptide, followed by a GH18 catalytic domain. In addition, subgroup B chitinases frequently contain a C-terminal CBM module (Hartl et al., 2012). Some subgroup B chitinases harbor additional domains such as a Ser/Thr-rich domain and a C-terminal glycosylphosphatidylinositol (GPI)-anchoring motif for attaching the protein to the plasma membrane (Takaya et al., 1998; Yamazaki et al., 2008).
In M. anisopliae, six subgroup B chitinases exhibit a GH18 catalytic domain and an N-terminal signal peptide. In contrast, ChiMaB5 is the only subgroup B chitinase that lacks the signal peptide. Both ChiMaB1 and ChiMaB4 have an additional C-terminal CBM1 for strong attachment to insoluble chitin. Interestingly, ChiMaB7 displays a domain-rich architecture comprising the signal peptide, GH18 catalytic domain, and a C-terminus Ser/Thr-rich region coupled with a GPI anchoring motif (Junges et al., 2014). Both CHI2 and CHI30 of M. anisopliae belong to subgroup B of GH18 protein. CHI2 comprises an N-terminal signal peptide, a GH18 catalytic domain, and a C-terminal CBM (Boldo et al., 2009). CHIT30 exhibits a similar domain architecture but lacks the C-terminal CBM module (Figure 2) (Junges et al., 2014).
Fungal subgroup C chitinases
3.1.3
Fungal subgroup C chitinases share several structural and functional features with subgroup A enzymes. For example, subgroup A and C enzymes are both exo-acting enzymes that belong to class V chitinases and have a deep and narrow substrate-binding groove. Nevertheless, subgroup C chitinases have several features that distinguish them from subgroups A and B. First, subgroup C enzymes are high molecular weight proteins (140–170 kDa). In addition, the domain organization of subgroup C chitinases contains a CBM 18 domain and a CBM 50 domain (also known as LysMs) for binding insoluble chitin. Both CBM 18 and CBM 50 are located at the N-terminal region of subgroup C (Seidl, 2008; Rajput et al., 2022).
M. anisopliae chitinases from the subgroup C exhibit multiple CBM modules in addition to a GH18 catalytic domain and an N-terminal signal peptide. For example, the amino acid sequence of ChiMaC1 and ChiMaC2 harbor two CBMs, i.e. CBM18 and CBM50, whereas ChiMaC3 has three CBMs, i.e. CBM18, CBM24, and CBM50. The only exception to this is ChiMaC4, which displays only one CBM, i.e. CBM18 (Junges et al., 2014). Interestingly, ChiMaC2 lacks the signal peptide region (Figure 2), suggesting that this enzyme is a cytoplasmic enzyme, which functions in fungal cell wall remodeling. It is also possible that ChiMaC2 is secreted via unconventional protein secretion pathways (Ding et al., 2012).
Fungal subgroup D chitinases
3.1.4
ChiMaD1 chitinase is a homolog of T. reesei chi18-15. ChiMaD1 orthologs exhibit endochitinase activity and contain a GH18 catalytic domain that is similar to the chitinase J (ChiJ) from Streptomyces species (Viterbo et al., 2001; Seidl et al., 2005; Ubhayasekera and Karlsson, 2012). The protein domain architecture is composed of an N-terminal signal peptide and a GH18 catalytic domain of bacterial origin (Figure 2) (Junges et al., 2014).
Deletion of ChMaD1-encoding gene in M. anisopliae did not affect the susceptibility of the cell wall to stresses or reduce secreted chitinases. The chMaD1-deletion mutant exhibits higher virulence rates against the cattle tick (Rhipicephalus microplus), but, surprisingly, displayed virulence rates similar to the wild type strain in the mealworm larvae (Tenebrio molitor) (Sbaraini et al., 2021). This suggests that cuticle variability across different species can induce the expression of different sets of chitinase genes.
GH20 and GH3 β-N-acetylglucosaminidases
3.2
β-N-acetylglucosaminidases (β-N-acetylhexosaminidases, NAGases, EC 3.2.1.52) are enzymes that catalyze the breakdown of chitobioses to GlcNAc monomers (Intra et al., 2008). NAGases are grouped into three GH families: GH3, GH20, and GH84. Fungal NAGases are commonly found in the GH20 family (Oliveira et al., 2018). Although the majority of GH3 NAGases are found in prokaryotes, a fungal GH3 NAGase was isolated and characterized from Rhizomucor miehei (Yang et al., 2014). Additionally, the genome of M. anisopliae harbors two NAGases from GH3 family. Therefore, only GH3 and GH20 NAGases will be covered in this section.
The characteristic domain architecture for GH20 proteins comprises a catalytic GH20 domain with an α/β TIM-barrel preceded by an N-terminal non-catalytic GH20b domain, which is required for protein expression and stabilization (Figure 2) (Liu et al., 2012; Val-Cid et al., 2015; Li et al., 2022b).
GH3 family enzymes exhibit a broad range of enzyme activities, such as β-D-glucosidases, α-L-arabinofuranosidases, β-D-xylopyranosidases, and N-acetyl-β-D-glucosaminidases (NAGases) (Macdonald et al., 2015). GH3 NAGases are considered single-domain enzymes because chitobiose hydrolysis is carried out exclusively by the N-terminal domain, which harbors the general acid/base catalytic residue and the catalytic nucleophile residue (Bacik et al., 2012; Macdonald et al., 2015). The presence of the general acid/base catalytic residue and the catalytic nucleophile residue in the GH3 catalytic domain is a distinctive feature for the identification of GH3 NAGases. In T. reesei, a protein, designated NAG3, with a GH3 NAGase domain was identified and thought to be the first T. reesei GH3 NAGase (Kappel et al., 2016). However, this protein lacks the catalytic residues responsible for the cleavage of amino sugar substrates, suggesting that it might function as a GlcNAc signal sensor/transducer. The protein was re-designated NGS1 (Ullah et al., 2025).
In an early study, a 110 kDa enzyme with NAGase activity was purified from M. anisopliae. The enzyme hydrolyzes GlcNAc dimers, trimers, and tetramers to GlcNAc monomers. The enzyme was feedback inhibited by the accumulation of GlcNAc (St. Leger et al., 1991). Nevertheless, the gene encoding for this enzyme was not identified.
Genome-wide screening revealed the presence of four NAGase enzymes in the genome of M. anisopliae. Two enzymes, MaNAG1 and MaNAG2, belong to GH20 NAGases and the other two, MaNAG3 and MaNAG4, belong to GH3 NAGases (Figure 2) (Oliveira et al., 2018). The molecular masses of MaNAG1 and MaNAG2 are ~67 kDa and ~61.5 kDa, respectively. The putative GH20 NAGases exhibit the characteristic GH20 domain architecture and an N-terminal signal peptide for secretion. MaNAG3 and MaNAG4 have a predicted molecular mass of ~99 and ~60.5 kDa, respectively. Bioinformatic analysis of MaNAG3 and MaNAG4 reveals the presence of the typical GH3 catalytic domain that contains the catalytic residues (Oliveira et al., 2018). The domain composition of MaNAG1–4 is summarized in Figure 2 of this review. The expression profiles were described by others (Oliveira et al., 2018).
In contrast to chitinases, which are present in high redundancy within fungal genomes, NAGases are encoded by one or a few genes. Nevertheless, NAGases play important roles during M. anisopliae cell wall extension, branching, and degradation (Oliveira et al., 2018). In addition, NAGases play an important role in metabolizing chitin as the sole carbon source. The deletion of NAGase-encoding genes, nag1 and nag2, in the mycoparasitic fungus Trichoderma atroviride abolished its ability to grow on chitin as the sole carbon source, highlighting the significance of NAGases in metabolizing chitin. Nevertheless, the deletion of NAGase-encoding genes did not impact fungal virulence, suggesting a minor role for chitin as a carbon source during parasitism (López-Mondéjar et al., 2009).
The biological functions of chitinases during pathogenesis and saprophytic growth
4
Entomopathogenic fungi have a complex life cycle that involves a parasitic stage on different arthropods, and a saprophytic stage in the soil or on infected cadavers (Gortari and Hours, 2008; Chen et al., 2020; St. Leger and Wang, 2020). Chitinases are produced during all stages of the fungal life cycle. Besides degrading exogenous chitin, chitinases are also key enzymes in fungal cell wall remodeling (Gruber et al., 2011). The number of these enzymes varies with the trophic relationship. For example, the genomes of the mycoparasitic fungi T. atroviride and T. virens harbor a greater number of chitinase-encoding genes than those of entomopathogenic and saprophytic fungi (Table 3). In contrast, the number of these genes in the entomopathogenic fungi is approximately equal to or lower than that in the saprophytic fungus T. reesei. Interestingly, the entomopathogenic fungus B. bassiana has more NAGase-encoding genes (5 genes) than other fungi (Table 3).
Chitinases exhibit distinct biological roles during the different stages of the fungal life cycle. Entomopathogenic fungi secrete an arsenal of chitinases that hydrolyzes the cuticle, facilitating host penetration and colonization. Inside the hemocoel, they metabolize host nutrients while exhibiting limited dependence on less−preferred carbon sources, including chitin (Gruber et al., 2011). Once the host nutrients are depleted, the entomopathogenic fungi switch to the saprophytic stage, growing on the cadavers of insects. During this stage, they have a higher capacity to metabolize the less-preferred carbon sources. This suggests a distinction between chitinase-mediated virulence and chitin metabolism. This is supported by the fact that the large arsenal of chitin-degrading enzymes in mycoparasitic fungi (Table 3) did not account for a good metabolism of chitin or its partially acetylated form, chitosan. In contrast, the entomopathogenic fungus B. bassiana exhibited a better growth on chitin despite having a fewer chitinase genes (Palma-Guerrero et al., 2008; Gruber et al., 2011). Furthermore, the deletion of NAGase-encoding genes in T. atroviride results in strains with abolished growth on chitin as the sole carbon source; nevertheless, the mutants retained full virulence (López-Mondéjar et al., 2009). Because of the overlapping functions of chitinases, it is challenging to precisely delineate their relative contributions to each lifestyle (Ortiz-Urquiza and Keyhani, 2016).
Differences have also been noted between parasitic and saprophytic fungi with respect to the GH domain architecture. The genomes of the mycoparasitic fungi harbor a larger set of CBMs than those of the saprophytic fungi. These modules facilitate stronger attachment of GH proteins to insoluble substrates (Kubicek et al., 2011; Xiao et al., 2012; St. Leger and Wang, 2020). In M. anisopliae, the GH18 subgroup C chitinases have CBM-rich domain architecture (Figure 2), suggesting a major role for these proteins during pathogenicity.
The chitinolytic enzymes of the entomopathogenic fungi M. anisopliae and Isaria cicadae, and the mycoparasitic fungi belonging to Trichoderma spp. exhibit a conserved domain architecture in subgroups A and B of GH18 chitinases (Figure 3). Subgroup A enzymes contain a GH18 catalytic domain and frequently have a signal peptide at the N-terminus; however, these enzymes lack CBM modules. Subgroups B share a homologous domain organization that comprises a signal peptide followed by a GH18 module. Some subgroup B chitinases might also have a CBM1 module at the C-terminus (Figure 3). Interestingly, only a third of the subgroup B chitinases from I. cicadae have a signal peptide (Peng et al., 2020). In Trichoderma spp., subgroup D chitinase lacks signal peptide at the N-terminus. Peng et al. (2020) did not report a subgroup D chitinase in I. cicadae. However, we identified a homolog of subgroup D chitinase when we ran a BLAST analysis (Table 3). Subgroup C chitinases display a divergent domain organization across all three fungi (Figure 3). Although they play a key role in the degradation of the exogenous chitin, surprisingly, the subgroup C chitinases from I. cicadae lack the signal peptide. The domain architecture of GH20 NAGases is conserved except for the I. cicadae homolog, which lacks the signal peptide (Figure 3). The lack of the signal peptide in several chitinolytic enzymes from I. cicadae suggests different biological roles or that the proteins are secreted via unconventional pathways (Ding et al., 2012). Interestingly, homologs of GH3 NAGases from M. anisopliae have not been identified in Trichoderma spp. or I. cicadae. However, our BLAST analysis identified GH3 NAGase proteins in these fungi (Table 3). For reviews on the contributions of chitinase diversity to fungal virulence, we recommend the reader to refer to Hartl et al. (2012), Gruber and Seidl-Seiboth (2012), and Ferreira and Soares (2023).
Regulation of gene expression of chitin-degrading enzymes in M. anisopliae
5
Chitinase expression has been shown to correlate with virulence in entomopathogenic fungi. Highly virulent M. anisopliae strains produced significant amounts of chitinases in comparison with low-virulence strains (Bhanu Prakash et al., 2012). The expression of M. anisopliae chitinases is induced by different substrates such as insect cuticle, chitin, and GlcNAc (Bogo et al., 1998; Krieger de Moraes et al., 2003; Barreto et al., 2004; Santi et al., 2010b; Beys-da-Silva et al., 2014; Rustiguel et al., 2016). Unmasking chitin is a prerequisite for the induction of chitinase expression. Chitinase activities of entomopathogenic fungi, including M. anisopliae, are detected at low levels on the cuticle surface during the initial penetration step. However, chitinase levels increase once chitin becomes accessible following the removal of masking proteins through proteolytic hydrolysis (St. Leger et al., 1996). Because of the dual nature of M. anisopliae as an insect pathogen and a soil saprophyte, chitinases are secreted in response to cuticles derived from living as well as dead insects. For example, the expression of chitinase genes was induced during cuticle penetration of the living Helicoverpa armigera insects, as well as during saprophytic growth on the cuticle of dead Spodoptera litura (Bhanu Prakash et al., 2012). In contrast, chitinase and NAGase activities are repressed in the presence of elevated concentrations of GlcNAc, suggesting a feedback inhibition mechanism regulating the expression of these genes (St. Leger et al., 1991; Krieger de Moraes et al., 2003; Barreto et al., 2004).
The spatiotemporal expression pattern of chitinases in M. anisopliae varies with the substrate. Gene expression analyses deciphered the spatiotemporal expression pattern of putative GH18 chitinases, and GH3 and GH20 NAGases in M. anisopliae. Autolysis of mycelia induced the highest number of chitinases. Additionally, chitin and GlcNAc induced the highest expression levels of subgroups A and B chitinases in mycelia. In contrast, subgroup C chitinases displayed significantly lower expression levels than subgroups A and B under the same induction conditions. Fungal conidia, appressoria, and blastospores displayed lower expression levels than mycelia (Junges et al., 2014). Variability in gene expression levels was also noted in GH3 and GH20 NAGases. In mycelia, chitin induced various expression levels of NAGase-encoding genes. MaNAG1 and MaNAG4 showed the highest expression levels, reaching ~50-fold and ~13-fold, respectively. Interestingly, the expression of MaNAG2 and MaNAG4 genes was constantly detected in the appressoria in the absence of chitin or cuticle, and in mycelia after challenging with chitin, suggesting a major role in host cuticle penetration. MaNAG3 seems to have a dual role in fungal pathobiology and cell wall remodeling during hyphal growth. This can be attributed to the fact that the MaNAG3-encoding gene was constitutively expressed in conidia and appressoria; however, was induced in mycelia after challenging with chitin and GlcNAc (Oliveira et al., 2018). The spatiotemporal regulation of chitinases in other fungi has been reviewed by Langner and Göhre (2016).
The expression of chitinases is also triggered by inducers other than GlcNAc-containing substrates. For example, the expression of M. anisopliae chi3 (ChiMaB2) gene is regulated in response to heat-shock stress conditions (at 42°C for 1 h). It is therefore presumed that the chi3 gene is a component of the fungal adaptation mechanism to heat-shock stress conditions (Staats et al., 2013).
Interestingly, infection with mycoviruses is another factor that affects chitinase activity. Mycoviruses are viruses that infect fungi, reducing their growth and virulence. Virus-infected Metarhizium majus and B. bassiana strains displayed an attenuated virulence and cuticular penetration (Wang et al., 2023; Zhang et al., 2023). Similarly, viral infection of M. anisopliae resulted in a significant reduction in endochitinase levels but not in N-acetylglucosaminidase levels (de la Paz Giménez-Pecci et al., 2002). However, such infection outcomes were found to be inconsistent across different studies. For example, Perinotto et al. (2014) showed that the virus-infected M. anisopliae strains exhibited higher virulence levels against Rhipicephalus microplus ticks and produced a significant amount of chitinases and proteases. These findings were supported by another study, which found that M. anisopliae and M. pingshaense mycovirus-infected strains exhibited elevated levels of virulence against the lepidopteran larvae of diamondback moth (Plutella xylostella) and the fall armyworm (Spodoptera frugiperda) (Guo et al., 2024). For more information on mycoviruses in entomopathogenic fungi, we recommend the reader to refer to recent reviews (Hough et al., 2023; Camargo et al., 2025).
M. anisopliae reacts to environmental changes via several sensory systems and a network of transcription factors. Screening of M. anisopliae genome identified 161 protein kinases and 510 putative transcription factors (Gao et al., 2011). However, the role of these proteins was not investigated in M. anisopliae. In this section, we will integrate key findings from other entomopathogenic fungi to identify conserved molecular mechanisms regulating gene expression of chitinases and NAGases in M. anisopliae (Figure 4).
The two-component system (TCS) is a key sensory system in fungi. The TCS system is a signal transduction pathway that consists of a membrane-bound sensor histidine kinase (Sln1) and two response regulator proteins, designated Ssk1 and Skn7 (Schaller et al., 2011). The histidine kinase (Sln1) is a hybrid protein that contains two domains: the kinase domain and the receiver domain. Sln1 senses environmental changes, and stimulates autophosphorylation of the kinase domain, which passes the phosphate group to the receiver domain of Sln1. Orthologs of Sln1 are involved in adaptation to different stress conditions (Catlett et al., 2003). In the entomopathogenic fungus Metarhizium rileyi, Sln1 is a key sensor of growth stress (Song et al., 2013, Song et al., 2015). Ypd1 is the next protein in the TCS. Ypd1 receives the phosphate group from Sln1 and transfers it to the downstream response regulators Ssk1 and Skn7. The later protein, Skn7, is a transcription factor that upregulates the expression of stress response genes and cell wall integrity in fungi (Fassler and West, 2011). The role of Skn7 in entomopathogenicity was analyzed in Metarhizium robertsii. Deletion of the skn7 gene in M. robertsii induced cell autolysis and the activation of extracellular chitinases and proteases, which implies that Skn7 functions as a repressor of chitinase and protease gene expression. In addition, bioinformatic analysis revealed the presence of the Skn7 binding site 5′-GGC(C/G)(A/G)-3′ in the promoter region of two chitinases (Shang et al., 2015). The homologs of TCS proteins and the two chitinases are conserved in the genome of M. anisopliae (Tables 4 and 5). This suggests similar sensory and regulatory mechanisms in both fungi (Figure 4).
Putative regulatory pathways and transcription factors mediating the expression of chitin-hydrolyzing enzymes in M. anisopliae. GlcNAc is imported to the cytoplasm via the N‐acetylglucosamine transporter NGT1, causing the activation of the Ron1 transcription factor. The accumulation of GlcNAc in the cell causes a feedback inhibition of chitinolytic enzyme activities. The accumulation of glucose in the cell activates the carbon catabolite repressor CreA, which represses the activity of chitinases and NAGases. Changes in environmental conditions activate the two-component system sensor histidine kinase (Sln1), which activates downstream proteins. The activation of the Skn7 transcription factor represses the chitinase genes. Fus3 MAPK activates the RNS1 and Ste12 transcription factors, which up-regulate the expression of chitinolytic enzymes. Alkaline pH activates the PacC protein, which up-regulates the expression of chitinases and NAGases. In addition, Sre1 and COH2 are transcription factors that regulate the expression of chitinases in M. anisopliae.
PacC is another regulator of chitinase expression in entomopathogenic fungi. PacC is a pH-dependent transcription factor that governs fungal adaptation to ambient environment. Under acidic pH conditions, PacC exhibits a closed protein conformation that protects the C-terminal domain from protease hydrolysis, and thus PacC remains in the cytoplasm. When pH changes to alkaline, PacC is activated by two proteolysis steps of the C-terminal region, which causes the protein to localize to the nucleus. Inside the nucleus, PacC binds to the consensus sequence 5′-GCCA(A/G)G-3′ (Li et al., 2022a). Deletion of pacC in M. robertsii resulted in a strain with impaired virulence and reduced adaptation to changes in pH. Additionally, the strain showed reduced chitinase activity; however, protease activity was not affected. In silico analysis of the promoter region of M. robertsii chitinases revealed the presence of the PacC consensus binding sequence in nine chitinase-encoding genes and one GH20 NAGase (Huang et al., 2015). Homologs of PacC transcription factor and PacC-regulated chitinases and NAGases have been identified in M. anisopliae (Tables 4 and 5). This suggests similar roles for PacC in M. anisopliae (Figure 4).
Gao et al. (2011) found that MAP kinase and cAMP dependent protein kinase A (PKA) pathways are up-regulated in M. anisopliae after challenging mycelia with cuticle from cockroach and locust, suggesting a role for both kinases in regulating the chitinase gene expression. Nevertheless, the relative contributions of both pathways in regulating chitinase expression were not investigated. Similarly, Chen et al. (2016) have shown that the MAPK cascade regulates the expression of chitinase genes in M. robertsii during infection, with two chitinases up-regulated (MAA_00157 and MAA_10509) and four chitinases down-regulated (MAA_08315, MAA_02744, MAA_03460, and MAA_00424). However, the downstream transcription factors that regulate chitinase gene expression were not identified in this study. In addition, the MAPK pathway has been reported to be conserved in several Metarhizium species other than M. anisopliae (Tang et al., 2022). We identified homologs of MAPK cascade proteins and the MAPK-regulated chitinases in M. anisopliae E6 strain (Tables 4 and 5).
Several regulators control the expression of chitinase genes in other entomopathogenic fungi. Ron1 is a transcription factor and a key regulator of GlcNAc catabolism (Figure 4). In addition, Ron1 plays an important role in fungal virulence, extracellular chitinase activities, and cell wall biosynthesis of the entomopathogenic fungus B. bassiana (Qiu et al., 2022). Sre1 is another transcription factor that regulates chitinase gene expression in fungi (Gómez et al., 2020). Deletion of sre1 in the mycoparasitic fungus Clonostachys rosea down-regulated the expression of chitinase genes (Piombo et al., 2024). In addition, RNS1 is a Myb transcription factor that is activated by Fus3 MAPK. Activated RNS1 binds to the 5’-ACCAGAC-3’ sequence in the promoter region and up-regulates the expression of cuticle-degrading genes, including one chitinase and one NAGase (Meng et al., 2021). Ste12 is a transcription factor that is also activated by Fus3 MAPK. Deletion of ste12 in M. rileyi reduced chitin content in conidia; however, the target chitinases were not investigated (Lin et al., 2021). Homologs of Ron1, Sre1, RNS1, and Ste12, and the chitinolytic enzymes that are controlled by these regulators, are conserved in M. anisopliae (Tables 4 and 5). This suggests a similar role for their homologs in M. anisopliae (Figure 4); however, the molecular characterization of these proteins is required. Furthermore, two novel transcription factors, designated COH1 and COH2, were shown to regulate chitinases in M. robertsii. Both transcription factors are required for fungal colonization of the hemocoel. In the Δcoh2 mutant, gene expression of three chitinases (MAA_00157, MAA_10456, and MAA_11305) was reported to be down-regulated (Zhang et al., 2021). However, when we ran BLAST analysis of these three chitinases, only a homolog of MAA_00157 was identified in the M. anisopliae genome. In contrast, MAA_10456 showed no homology to any known proteins. Our BLAST analysis of MAA_11305 identified a hypothetical protein MANI_008911 (accession number: KFG79813) with no GH18 catalytic domain. Instead, the protein harbors an aminoglycoside 3’-phosphotransferase and Choline kinase domain, which suggests that this protein is not a chitinase. For reviews on the role of transcription factors in entomopathogenic fungi, we recommend the reader to refer to a recent review (Song et al., 2025).
The mechanism that mediates GlcNAc transportation into the cell has been explored in T. reesei. Extracellular GlcNAc is imported to the cell via the N‐acetylglucosamine transporter (NGT1), which belongs to the major facilitator superfamily (MFS). Once GlcNAc is internalized in the cell, it activates catabolic gene expression of approximately 2100 genes. Ron1 serves as a major regulator for these genes, while the carbon-catabolite repressor CRE1 is a key repressor (Figure 4) (Song et al., 2019; Ullah et al., 2025). The M. anisopliae genome harbors a putative homolog of NGT1 (Table 4). In addition, the carbon catabolite-repression mechanism has been proposed in M. anisopliae based on the fact that chitinase activity is abrogated with increasing concentrations of glucose alone or in combination with chitin (Krieger de Moraes et al., 2003; Barreto et al., 2004). However, these studies lack comprehensive molecular analyses.
Perspectives and applications of chitinases from M. anisopliae
6
The potential of M. anisopliae as a biocontrol agent against agricultural pests have been reviewed by many reports (Ferreira and Soares, 2023; Mesquita et al., 2023). Chitinases from M. anisopliae have been extensively characterized at the molecular and biochemical levels; however, relatively few studies have explored their broader applications beyond insect pathogenicity, particularly in plant protection and strain improvement strategies. Emerging research demonstrates that M. anisopliae chitinases can be exploited both directly and indirectly to enhance resistance against plant pathogens and insect pests.
Several studies have evaluated the potential of M. anisopliae chitinases in engineering transgenic plants with improved resistance to fungal diseases and insect infestations. For example, transient expression of the chit1 gene from M. anisopliae in cotton plants resulted in 9% mortality of adult whiteflies, Bemisia tabaci, feeding on the transformed plants (Anwar et al., 2019). Similarly, tobacco plants expressing chit1 under the control of the CaMV 35S promoter exhibited enhanced resistance to infection by the plant pathogenic fungus Rhizoctonia solani (Kern et al., 2010). These findings indicate that Metarhizium-derived chitinases can function effectively in heterologous plant systems to confer protection against both fungal and insect threats.
In addition to plant-based approaches, chitinases from M. anisopliae have been used to enhance antagonistic activity in mycoparasitic fungi, thereby improving biological control potential. Overexpression of the chit1 gene in T. harzianum significantly increased antagonism against the plant pathogen Botrytis cinerea (Li et al., 2012). Likewise, expression of M. anisopliae chitinases in T. koningi enhanced lethality against the Asian corn borer larvae (Xia et al., 2018). These studies highlight the versatility of M. anisopliae chitinases as functional tools for improving fungal biocontrol agents across different host systems.
Beyond these applied perspectives, chitinases play a central mechanistic role in fungal biocontrol efficacy. They are considered key virulence determinants that directly influence the performance of entomopathogenic fungi by mediating degradation of the insect cuticle during host penetration. In M. anisopliae, chitinases act synergistically with proteases and lipases to enable successful entry into the insect hemocoel, a prerequisite for infection and host colonization (Ortiz-Urquiza and Keyhani, 2013; Wang and Wang, 2017). The secretion, regulation, and enzymatic activity of chitinases are therefore critical factors governing biocontrol success.
Experimental evidence further demonstrates that enhancing chitinase expression can improve fungal virulence and infection efficiency. Overexpression of the chitinase gene Bbchit1 in Beauveria bassiana resulted in accelerated host mortality and increased pathogenicity (Fang et al., 2005). These results provide proof-of-concept that strengthening cuticle-degrading capacity can enhance the effectiveness of fungal biocontrol agents, although such approaches must be carefully evaluated for ecological safety and regulatory compliance.
Finally, successful biocontrol applications depend not only on the genetic potential but also on the effective enzyme activity at the host surface. Sustained contact between fungal propagules and the insect cuticle is essential for sufficient enzymatic degradation to occur, emphasizing the importance of formulation and application strategies that support chitinase-mediated penetration (Lovett and St. Leger, 2017). Consequently, integrating molecular insights into chitinase function with practical deployment strategies provides a rational framework for improving the consistency, reliability, and performance of Metarhizium-based biological control systems (Wang and Wang, 2017).
Discussion
7
M. anisopliae is a promising entomopathogenic fungus that is commercially used in the biological control of insect pests. Overall, this review emphasizes the diversity of M. anisopliae chitinases, and highlights their fungal subgroup distribution, domain architecture, and the conserved mechanisms underlying gene expression. M. anisopliae possesses a complex arsenal of chitin-degrading enzymes. Nevertheless, only three chitinases were characterized. Therefore, a significant amount of work is still required to characterize the remaining chitinases. Similarly, the identification of signaling pathways that regulate the expression of chitinase genes remains elusive. Chitinase genes encode enzymes with overlapping activities, and therefore other chitinases can mask the phenotype of the deleted gene, a mechanism known as functional redundancy. Because of the redundancy of chitinases in the genome, it is challenging to identify the precise function of an individual chitinase gene using a single knockout mutant. Therefore, double- or triple-knockout mutants should be generated. However, the disruption of multiple chitinase genes in a single strain is limited by the number of selectable markers currently available. One way to overcome this challenge is by recycling the selectable marker genes using the Cre–loxP recombination system. Another approach is to employ CRISPR–Cas9 technology to simultaneously knock out multiple chitinase genes or to develop a marker-free gene-deletion strategy.
The identification of chitinases and their regulatory proteins can also be achieved via a combination of genomics, transcriptomics, and proteomics approaches, which allow for high-throughput analyses of chitinases and their upstream regulators. For example, investigating the expression profiling of chitinase genes under different insect cuticular components, host developmental stages, or environmental stresses provides insight into the conditions under which chitinase genes are turned on. Virulence assays across multiple insect hosts and environmental conditions reveal context-dependent redundancy. In addition, heterologous expression of M. anisopliae chitinases in bacteria, filamentous fungi, and yeast allows for deciphering the biochemical properties of chitinases and their biological function. Transgenic plants that are resistant to insect pests or fungal pathogens can be generated by expressing M. anisopliae chitinases in plants. Domain engineering is another approach to analyze the function of domains and engineer chimeric chitinases for industrial degradation of chitin.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdulhameed A. S. Al Omari R. H. Abdullah S. Al-Masud A. A. Abualhaija M. Algburi S. (2025). Nanoarchitectonics of biohybrid polymer nanocomposite of carboxylated chitosan-phthalate/Sn O 2 nanoparticles for dye wastewater treatment: characterisation, isotherm, kinetic, and adsorption optimisation. Int. J. Environ. Anal. Chem. 105, 6799–6820. doi: 10.1080/03067319.2024.2429792 · doi ↗
- 2Al-Nasrawi H. Shalan N. Abualsoud B. M. Nsairat H. (2024). Preparation, characterization and in vitro evaluation of 5-fluorouracil loaded into chitosan–acacia gum nanoparticles. Ther. Deliv. 15, 339–353. doi: 10.4155/tde-2023-0136, PMID: 38469691 PMC 11160445 · doi ↗ · pubmed ↗
- 3Andersen S. O. (2009). “ Chapter 94 - exoskeleton,” in Encyclopedia of insects, 2nd ed.Eds. Resh V. H. CardéR. T. ( Academic Press, San Diego), 339–342. doi: 10.1016/B 978-0-12-374144-8.00103-X · doi ↗
- 4Anwar W. Javed M. A. Shahid A. A. Nawaz K. Akhter A. Ur Rehman M. Z. . (2019). Chitinase genes from Metarhizium anisopliae for the control of whitefly in cotton. R. Soc Open Sci. 6, 190412. doi: 10.1098/rsos.190412, PMID: 31598241 PMC 6731705 · doi ↗ · pubmed ↗
- 5Bacik J.-P. Whitworth G. E. Stubbs K. A. Vocadlo D. J. Mark B. L. (2012). Active site plasticity within the glycoside hydrolase nag Z underlies a dynamic mechanism of substrate distortion. Chem. Biol. 19, 1471–1482. doi: 10.1016/j.chembiol.2012.09.016, PMID: 23177201 · doi ↗ · pubmed ↗
- 6Baratto C. M. Dutra V. Boldo J. T. Leiria L. B. Vainstein M. H. Schrank A. (2006). Isolation, Characterization, and Transcriptional Analysis of the Chitinase chi 2 Gene (DQ 011663) from the Biocontrol Fungus Metarhizium anisopliae var. anisopliae. Curr. Microbiol. 53, 217–221. doi: 10.1007/s 00284-006-0078-6, PMID: 16874542 · doi ↗ · pubmed ↗
- 7Baratto C. M. Vanusa da Silva M. Santi L. Passaglia L. Schrank I. S. Vainstein M. H. . (2003). Expression and characterization of the 42 k Da chitinase of the biocontrol fungus Metarhizium anisopliae in Escherichia coli. Can. J. Microbiol. 49, 723–726. doi: 10.1139/w 03-085, PMID: 14735222 · doi ↗ · pubmed ↗
- 8Barreto C. C. Staats C. C. Schrank A. Vainstein M. H. (2004). Distribution of chitinases in the entomopathogen metarhizium anisopliae and effect of N-acetylglucosamine in protein secretion. Curr. Microbiol. 48, 102–107. doi: 10.1007/s 00284-003-4063-z, PMID: 15057476 · doi ↗ · pubmed ↗