Functional connectivity in infants’ visual cortex and its links to motion processing and autism
Irzam Hardiansyah, Giorgia Bussu, Sven Bölte, Emily J.H. Jones, Terje Falck-Ytter

TL;DR
This study explores how brain connectivity in infants' visual cortex relates to autism symptoms and motion processing, suggesting early markers for autism.
Contribution
The study identifies atypical visual cortical functional connectivity as a potential early marker for autism symptoms.
Findings
Gamma connectivity between midline and far-lateral visual cortex during social scenes links to later autism symptoms and motion perception.
Higher theta connectivity in non-social scenes correlates with more autistic symptoms at follow-up.
The findings suggest a shared mechanism between social perception and motion processing in early brain development.
Abstract
In a previously published study, we found atypical visual cortical laterality patterns during global motion perception in 5-month-old infants who showed high levels of autistic symptoms in toddlerhood. Here, using data from a separate experiment within the same recording session, we examined whether these results could reflect altered visual cortical functional connectivity in theta, alpha, and gamma rhythms. We assessed this in a sample of 5-month- old infants (n = 59; 39 elevated familial likelihood of autism) by means of electroencephalography (EEG) when they were watching videos showing social and non-social scenes. Gamma connectivity between midline and far-lateral visual cortex when viewing social scenes was linked to both later autism symptoms and global motion visual cortical laterality we reported in the previous study. This may indicate a shared integrative mechanism…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
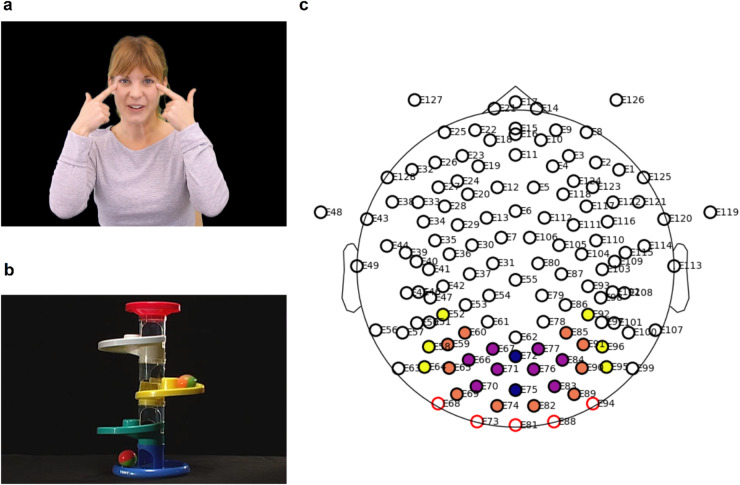 Figure 1
Figure 1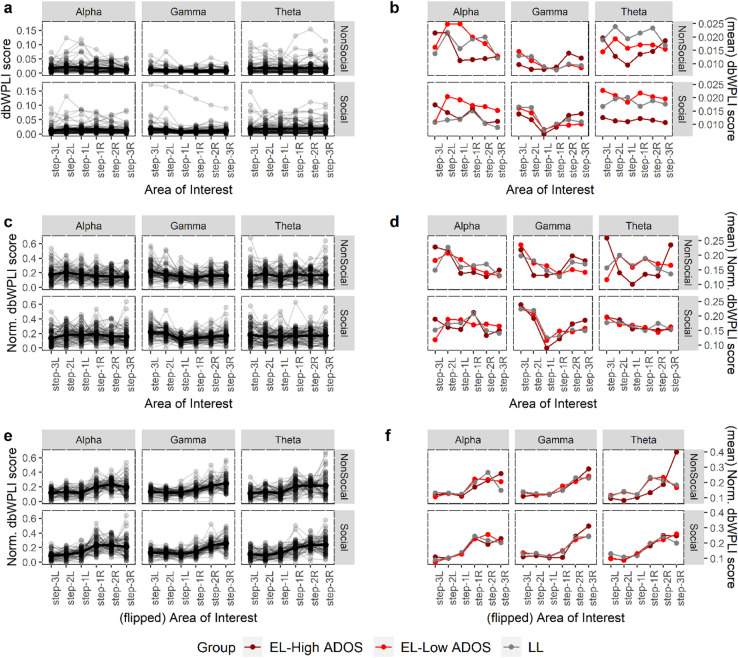 Figure 2
Figure 2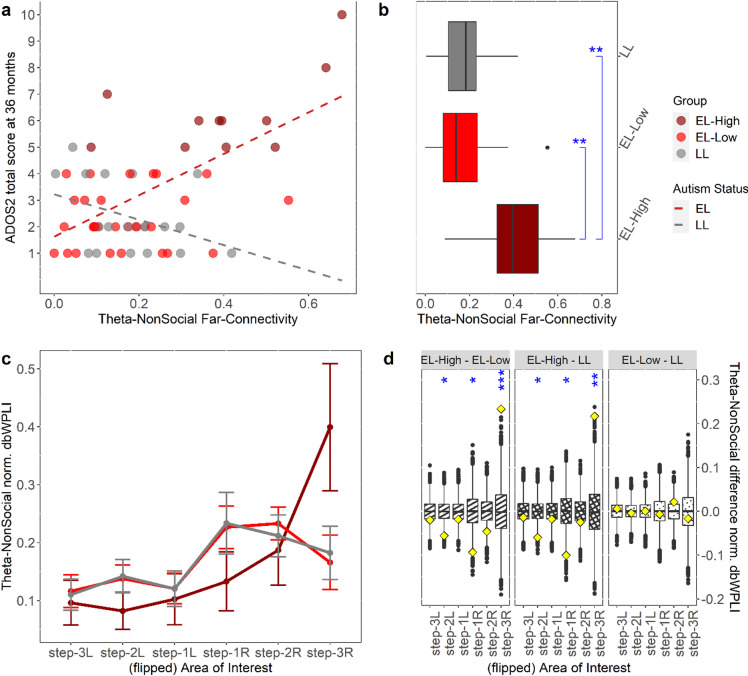 Figure 3
Figure 3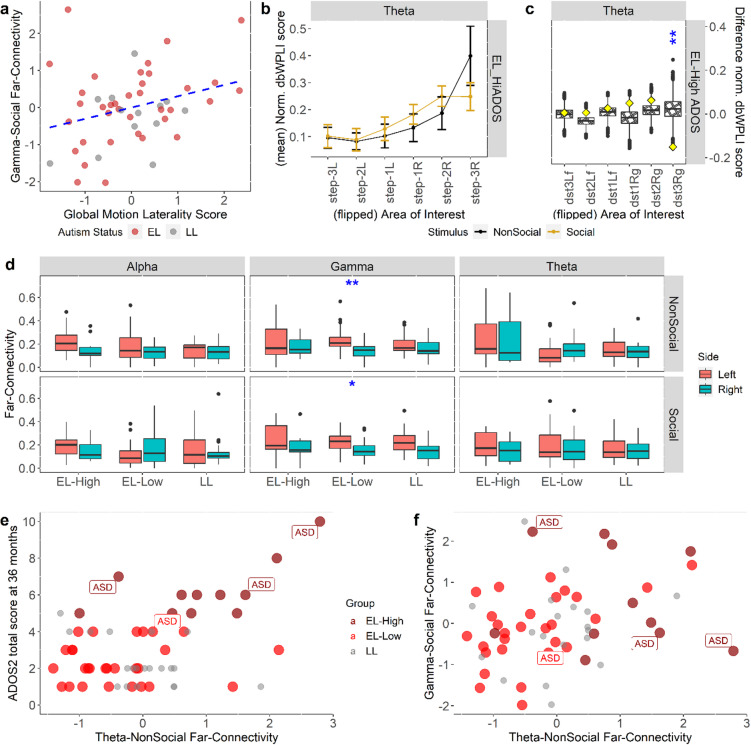 Figure 4
Figure 4- —Uppsala University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAutism Spectrum Disorder Research · Action Observation and Synchronization · Child and Animal Learning Development
Introduction
Although it is well established that the emergence of autistic symptoms is linked to differences in brain development, much remains to be understood concerning the specific nature of these differences. Both altered structural and functional brain development have been implicated^1–4^, and one infant brain imaging study suggested that the emergence of autistic symptoms was linked to a hyper-expansion of the cortex across different areas in the brain^5^, including the visual cortex. Consistently, we recently found atypical (highly lateralized) activation of the visual cortex during low-level global perceptual processing^6^ in infants with high levels of autistic symptoms in toddlerhood^7^. This study highlighted the potential developmental implication of atypical motion processing in the visual cortex of autistic children. These findings from infants are also in line with a recent study of transcriptomic alterations in the visual cortex of autistic adults^8^(see also ref^9^. We have also observed that highly lateralized EEG activation during global motion processing is linked to later language development^10^.
Altered connectivity has been postulated to be a functional brain signature in autism^11–14^, and it is possible that the above-mentioned differences in global motion processing are associated with differences in connectivity in the visual cortex among infants later diagnosed with autism. Therefore, we investigated the functional connectivity in the visual cortex in a cohort of 5-month-old infants from the Early Autism in Sweden (EASE). The cohort resembled the one from earlier study^7^, but was larger, and included additional participants that had reached the study endpoint (see Results, also Supplementary Tables 1–2 for analysis-specific exclusions and sample sizes). We aimed to assess (i) if atypical functional connectivity in the visual cortex is linked to later autism diagnosis and (ii) if there is a correlation between the connectivity and global motion processing. Specifically, we performed analyses of within-visual cortex resting-state connectivity based on infants’ EEG data, using the debiased Weighted Phase-Lag Index (dbWPLI)^15^, a phase-lag measure that had been reported to be robust for use with infants^16,17^. An analysis plan was registered prior to data analysis https://osf.io/e8y5x.
Given that our previous findings indicated atypical balance between midline and lateral (peripheral) activity in the visual cortex during motion processing in infants subsequently showing autistic symptoms^7^, we focussed on patterns of synchronization between the primary visual cortex (V1) and lateral extrastriate cortices (Fig. 1c). We assessed EEG connectivity within the visual cortex by adapting a graph-based approach that has previously been used with fMRI data^18,19^. Our approach is similar to that used in ref^20^ but with simplified graph measures instead of the traditional ones (e.g., path length, clustering, etc.). These simplified measures were defined in our analysis plan and allowed us to make specific inferences about communications between midline and lateral areas of the visual cortex (see full descriptions in Methods).
The infants were watching short movie clips, either showing a social (a woman singing nursery rhymes; Fig. 1a) or a non-social (colourful rotating toys; Fig. 1b) scene. Thus, in contrast to our previous experimental stimuli used to probe global motion processing specifically^7^, the current stimuli showed visual motion in a (for infants) ecologically meaningful context. We assessed functional connectivity of theta, alpha, and gamma brain rhythms; see Methods for more details. The three frequency bands were slightly shifted to adjust for the young age of the participants and were chosen for being the most commonly investigated brain rhythms in the contexts of cognitive development^21,22^, social attention^23^, and autism^24–26^, also in the context of infancy research^27,28^. Furthermore, infants’ theta and alpha rhythms has been shown to possess similar functional significance as the analogous rhythms in adults^29,30^. We chose not to assess functional connectivity during the previously reported global motion experiment^7^, which employed steady-state visually evoked potential (SSVEP) method, because of the strong oscillations elicited by the stimuli themselves (alternating between signal and noise several times per second).
As in the previous report^7^, we compared three groups: (i) infants with elevated likelihood of autism and significant later autistic symptoms (EL-High ADOS), (ii) infants with elevated autism likelihood and few or no later symptoms (EL-Low ADOS), (iii) and infants with low likelihood of autism (LL). The categorization of autistic symptom presentation within the EL group was based on established cut offs on the infants’ Calibrated Severity Scores (CSS)^31^ from the Autism Diagnostic Observation Schedule 2nd edition (ADOS-2)^32^ assessed at 36 months of age (see Methods). Following our analysis plan, we created analytical variables called the Near-Connectivity and Far-Connectivity, defined as the score of dbWPLI connectivity from the midline occipital cortex to the lateral occipital region immediately adjacent to it (“step-1 distance”) and from the midline to the furthest lateral occipital region (“step-3 distance”) included in our areas of interest, respectively (see Fig. 1c, also described fully in Methods). Thus, in analogy to the previous Laterality Score^7^, the two variables should enable us to probe whether atypical balance exists between functional connectivity in the proximity of V1 and that in the lateral (extrastriate) areas. Finally, we used the same Global Motion Laterality Score variable as in the previous work^7^ to represent (lateral) global motion processing and analyzed its relation to the dbWPLI connectivity variables we derived in this study. Notably, in this paper we use the term “laterality” to denote peripheral versus central (midline) area of the visual cortex, rather than left versus right.
Fig. 1. Experimental setup. (a) A screenshot of stimulus video clip for the social condition (see Supplementary Video 1). (b) Screenshots from the non-social (toy) stimulus video clip (see Supplementary Video 2). (c) EEG electrode layout and the part of occipital scalp region that we defined as visual cortex (described fully in Methods) were divided into 6 areas of interest (AoIs) relative to, but excluding, the two midline “seed” electrodes coloured in dark blue. These 6 AoIs extended bilaterally and covered successively further regions of the occipital cortex, forming step-1 (purple), step-2 (orange), and step-3 (yellow) block of electrodes, where the “step” reflected the average (binned) Euclidian distance of the corresponding block from the seed electrodes. Crossed channels were discarded. The Euclidean distances were calculated from the 3-dimensional coordinates of each electrode estimated by EEGLab. A mean dbWPLI functional connectivity score was then calculated for each of the 6 AoIs relative to the seed region, and scores for the step-1 area (left and right, separately) were defined as the Near-Connectivity and those for the step-3 area (left and right, separately) as the Far-Connectivity. Note: A written informed consent was obtained from each of our collaborators who acted as a model in Fig. 1a and in Supplementary Video 1 (see also Methods).
Results
Sample sizes and descriptive statistics
Due to the different approaches of EEG data pre-processing and analysis employed to compute the Near-/Far-Connectivity and Global Motion Laterality Score^7^ variables (see Methods for more details), the sample size differed for the analyses: N_Connectivity_ = 68, N_LatScore_ = 95 (Supplementary Tables 1–2). Descriptive statistics for each resulting dataset are presented in Supplementary Tables 3–4.
Topographical distribution of midline-to-lateral functional connectivity within the visual cortex
Plots of topographical functional connectivity across the 6 AoIs defined in Fig. 1c are shown in Fig. 2 (n = 59; nEL−HiADOS = 11, nEL−LoADOS = 28, nLL = 20). The left column (2a,** c**,** e**) showed individual topographical plots, while the right column (2b,** d**,** f**) show group average topographies. The plots are presented in the chronology of our analysis steps (see Methods): original dbWPLI scores (top panel, 2a,** b**), vector-normalized dbWPLI scores (middle panel, 2c,** d**), flipped (all maxima arbitrarily allocated to the right side) and normalized dbWPLI scores (bottom panel, 2e,** f**).
The EL-High ADOS group had substantially lower overall raw Theta-Social connectivity (lower-right panel of Fig. 2b), and a different raw Theta-NonSocial topographic morphology (upper-right panel of Fig. 2b) compared to the other two groups. The flipped and normalized plots revealed clear patterns of unilateral, i.e., only right or left but not both, connectivity between midline and lateral occipital areas. The Theta-NonSocial topography for the EL-High ADOS group remained distinctive in the flipped plots (upper-right panel of Fig. 2f). See Supplementary Table 5 for formal tests of across-group differences.
Fig. 2. Topographical theta, alpha, and gamma connectivity during observation of social and non-social stimuli. a. Raw (unnormalized, unflipped) topographical dbWPLI scores across the occipital electrodes in theta, alpha, and gamma rhythms for the two stimulus conditions from all individual participants irrespective of group. b. Group average plots (EL-High ADOS = dark red; EL-Low ADOS = red; LL = grey) from the topographical plots in a for the six experimental (3 bands x 2 stimuli) conditions. c-d. Corresponding normalized (see Methods) topographical dbWPLI scores across the occipital scalp. e-f. Corresponding flipped topographical dbWPLI scores. Here all individual maxima are always to the right (hence ‘left’ and ‘right’ are arbitrary; see Methods). The bilaterality patterns seen in b and d, especially for Gamma-NonSocial, Gamma-Social, and Theta-NonSocial conditions, were due to the almost equal numbers of infants having a peak either on the left or right side of the topography, but very rarely on both, which was made apparent in f. For all plots, n = 59. See Fig. 3c and Supplementary Fig. 2 for the error bars (for b,** d**,f) and their corresponding non-parametric difference testing for the flipped normalized topographies in c. Note: In all figures’ x-axis labels, ‘-L’ and ‘-R’ (e.g., in “step3L”) denote left and right, respectively.
Association between infants’ visual cortical functional connectivity and autistic symptoms in toddlerhood
For infants who had complete data for dbWPLI scores and ADOS-2 score (n = 59; nEL−HiADOS = 11, nEL−LoADOS = 28, nLL = 20), the non-parametric variable selection (see Methods) provided 5 connectivity variables –Theta-NonSocial, Gamma-Social, Gamma-NonSocial Far-Connectivities and Theta-NonSocial, Alpha-Social Near-Connectivities– as the most predictive of ADOS-2 CS total score at 36 months within the repeated 10-fold cross-validation scheme. In the subsequent model-fitting (see “Modelling association between functional connectivity and autism” in Methods), however, only the three Far-Connectivity variables were found significant: Theta-NonSocial (β = 1.12, partial-η^2^ = 0.23, ***p < .001, n = 59), Gamma-Social (β = 0.60, partial-η^2^ = 0.09, *p = .036, n = 59), and Gamma-NonSocial (β = − 0.59, partial-η^2^ = 0.08, *p = .047, n = 59), together accounting for ~ 44% of variability in ADOS-2 score. Two covariates, autism likelihood status (EL = 1, LL = 0) (β = 0.98, partial-η^2^ = 0.09, *p = .034, n = 59) and sex (M = 1, F = 0) (β = 0.87, partial-η^2^ = 0.08, *p = .047, n = 59) were found significant. Following the preregistered analysis plan, we did an analysis of moderation by autism likelihood status. This analysis revealed a significant interaction between likelihood status and Far-Connectivity in the Theta-NonSocial condition, such that the effect is stronger in the EL group (β = 1.91, partial-η^2^ = 0.14, **p = .008, n = 59), while in both Gamma-Social and Gamma-NonSocial (both p > .070) the effects were no longer significant at α = 0.050. The resulting model now explained more variability (R^2^ = 49%) in ADOS-2 scores compared the main-effect model. See Supplementary Table 5 for more details on these modelling results.
A follow up analysis revealed that the association of Theta-NonSocial Far-Connectivity with ADOS-2 score was driven by the EL sub-sample (Fig. 3a). Further group-level analyses showed that the EL-High ADOS group displayed greater Theta-NonSocial Far-Connectivity scores overall (Fig. 3b), which reflect their distinctive average topographical connectivity pattern in that condition (Fig. 3c; see also Supplementary Table 5).
Fig. 3. Different midline-to-far lateral visual cortical theta connectivity during non-social stimulus viewing is associated with having more symptoms at three years of age. (a) Correlation between Theta-NonSocial Far-Connectivity and ADOS-2 CSS total score (r = .50, ***p < .001, n = 59; regression line not shown) was driven by a strong positive correlation (r = .64, ***p < .001, n = 39) in the EL sub-sample (LL sample: r = − .38, p = .095, n = 20). (b) Accordingly, group difference analysis of Theta-NonSocial Far-Connectivity revealed significant differences between the EL-High ADOS group (n = 11) and both EL-Low ADOS (d = 1.61, **p = .005, n = 28) and LL (d = 1.57, **p = .008, n = 20) groups, but no difference was seen between the latter two groups (p = .492); see also Supplementary Fig. 1 for plots of the other connectivity variables. (c) Mean flipped normalized full topographic patterns of the functional connectivity for the three groups (crf. Figure 2f), showing that the EL-High ADOS group’s connectivity result is distinctive from the other two groups (see d for statistical group comparisons). (d) Results of non-parametric permutation testing of pairwise group differences (see Methods) of topographic patterns shown in c, done for each of the six areas of interest making up the full topography, where each panel presents a comparison of two groups (boxplots are simulated distributions using 10,000 randomization of the group labels, diamonds actual point values in our data, stars mark significance); see also Supplementary Fig. 2 for plots of the other 5 conditions. Compared to EL-Low ADOS infants, the EL-High ADOS infants had on average stronger connectivity in step-3R AoI (M = 0.234, d = 4.16, ***p < .001) but weaker in step-1R AoI (M = − 0.094, d = -2.41, *p = .050) and in step-2 L AoI (M = − 0.056 d = -2.36, *p = .050) in the theta band during non-social video viewing. Comparison of EL-High ADOS group with LL infants produced similarly pronounced differences (step-3R AoI: M = 0.217, d = 3.67, **p = .004; step-1R AoI: M = − 0.100, d = -2.44, *p = .050; step-2 L AoI: M = − 0.060, d = -2.44, *p = .050) in theta band in response to this non-social stimulus. Comparison between EL-Low ADOS and LL infants did not produce any difference (all p > .450). All comparisons were FDR-corrected. (Notes: EL = Elevated Likelihood; LL = Low Likelihood; EL-High = EL-High ADOS; EL-Low = EL-Low ADOS; step-dL/-R = step-dLeft/-Right, d = 1, 2, 3; * = p < .05; ** = p < .01; *** = p < .001)
Linking laterality of visual cortical functional connectivity and lateral global motion processing in infancy to later autism
With n = 43 (nEL−HiADOS = 7, nEL−LoADOS = 21, nLL = 15) infants who had complete data for Near- and Far-Connectivity, Global Motion Laterality Score, and ADOS-2 score, only Theta-NonSocial (β = 0.56, partial-η^2^ = 0.11, *p = .039, n = 43) and Gamma-NonSocial (β = − 0.59, partial-η^2^ = 0.11, *p = .046, n = 43) Far-Connectivities were significant when we refitted the main-effect model of functional connectivity association with autism. The model had a R^2^ of 32.2%. We did not fit the interaction model due to low power to detect an interaction effect. Expanding the main-effect model by adding Global Motion Laterality Score as an extra predictor (see “Inferring relationship between functional connectivity and global motion processing” in Methods), both Theta-NonSocial (β = 0.63, partial-η^2^ = 0.15, *p = .017, n = 43) and Gamma-NonSocial (β = − 0.57, partial-η^2^ = 0.11, *p = .042, n = 43) Far-Connectivities were still significant, along with the Global Motion Laterality Score (β = 0.47, partial-η^2^ = 0.15, *p = .045, n = 43) itself. The model’s R^2^ was 39.7% and a subsequent nested likelihood ratio test of the models before and after the expansion showed that this increase in variance explained was significant (χ_1_^2^ = 5.01, p = .025). No covariate turned out significant in either model. Notably, comparison of ANOVA results of these two models indicated that variance explained by Global Motion Laterality Score was distinct from that explained by either Theta-NonSocial or Gamma-NonSocial Far-Connectivity; however, it reduced the variance explained attributed to Gamma-Social Far-Connectivity (see Table 1). A subsequent analysis of association between Global Motion Laterality Score and all 12 Near-/Far-Connectivity variables, using the same variable selection procedure as in the previous section (also see Methods), revealed Gamma-Social Far-Connectivity (β = 0.36, partial-η^2^ = 0.12, *p = .021, n = 49) as the only connectivity variable significantly associated with Global Motion Laterality Score (Fig. 4a), consistent with the above results. Complete modelling results can be found in Supplementary Table 5.
As a supplementary analysis, we confirmed that for the expanded sample of n = 82 (nEL−HiADOS = 19, nEL−LoADOS = 44, nLL = 19) infants, the previously reported association between Global Motion Laterality Score and ADOS-2 total score at 36 months still held (β = 0.53, partial-η^2^ = 0.07, **p = .018, n = 82); see Supplementary Table 5.
Table 1. Analysis of variance of functional connectivity association models of ADOS CSS at 36 months without and with global motion laterality score (GMLS).PredictorModel without GMLSModel with GMLSSum of Squarep-valueSum of Squarep-valueSex3.440.2101.520.381Autism Likelihood Status1.960.3410.820.519Age (5 months)7.010.0776.680.072Theta-NonSocial Far-Connectivity9.670.03912.160.017Gamma-NonSocial Far-Connectivity9.070.0468.600.042Gamma-Social Far-Connectivity4.460.1551.010.474Global Motion Laterality Score8.370.045Residuals76.0567.68Degrees of freedom3635
Functional connectivity differences between stimulus types and between left and right hemispheres
Although not specified in the preregistered Analysis Plan, we performed two additional analyses to aid the interpretation of the results: (i) social vs. non-social topographic connectivity (both raw and flipped-normalized) differences in each frequency band by group; (ii) left/right far-lateral connectivity difference in each experimental condition by group. Only in the flipped topographies, we found lower social relative to non-social theta connectivity at step-3R AoI in EL-High ADOS group (Figs. 4b, c), but the reverse (higher social relative to non-social theta connectivity) at the same AoI in the EL-Low ADOS group (Supplementary Fig. 3). No lateralization (left vs. right) difference was found in any condition or group (Fig. 4d). See Supplementary Table 5 for detailed results.
Autism diagnosis and non-social theta and social gamma far-lateral connectivity
There were only four individuals diagnosed with autism in our sample, therefore we did not perform a formal analysis of autism diagnosis. Nonetheless, we provided plots (Figs. 4e, f) which showed that all but one case corresponds to an extreme score in either Theta-NonSocial or Gamma-Social Far-Connectivity variable.
Fig. 4. Linking functional connectivity laterality to global motion processing and ASD diagnosis. (a) Association between Gamma-Social Far-Connectivity and Global Motion Laterality Score was significant (r = .30, *p = .038, n = 49) for all participants who had data for both variables. (b) Mean flipped normalized topographic patterns of theta functional connectivity for the EL-High ADOS group, showing an elevated far-connectivity during the viewing of non-social relative to social stimuli (see c for the formal statistical comparison). (c) Accordingly, non-parametric permutation testing of between-stimulus difference (see Methods) of topographic patterns shown in c revealed a significant difference (social – non-social) in Step-3R AoI (d = -3.05, *p = .002, n = 11); boxplots are simulated distributions using 10,000 randomization of the stimulus labels, diamonds actual point values in our data, stars mark significance). See also Supplementary Fig. 3 for the complete plots and permutation testing of between-stimulus differences for all the groups and bands. (d) Wilcoxon tests of group differences in left or right (i.e., right minus left) connectivity asymmetry was performed to see whether left-sidedness or right-sidedness in any of the three frequency bands and two stimulus types is more related to autism. No group difference was found, except for in gamma frequency band and in both stimulus types (Gamma-NonSocial: d = -1.03, p = .004, n = 58; Gamma-Social: d = − 0.82, p = .031, n = 58), but only for the EL-Low ADOS group. In both cases, left asymmetry was more prevalent. Boxplots show the sample median, and the first and third quartiles; whiskers show minimum and maximum (± 1.5 s.d.); dots are outliers (± 1.96 s.d.). (e) Reproduction of Fig. 3a but now with autism diagnosis cases shown. (f) Individuals diagnosed with autism are shown as a function of both non-social theta connectivity laterality (x-axis) and social gamma connectivity laterality (y-axis). All cases in EL-High ADOS group (dark red) correspond to extreme laterality in either non-social theta or social gamma connectivity. See Supplementary Table 5 for numerical details of all results presented here. All p-values were FDR-corrected. (Notes: EL = Elevated Likelihood; LL = Low Likelihood; EL-High = EL-High ADOS; EL-Low = EL-Low ADOS; * = p < .05; ** = p < .01; *** = p < .001).
Discussion
We investigated infants’ visual cortical functional connectivity and tested its link to both our earlier global motion processing findings^7^ and to later presentation of autistic symptoms. As the current study was motivated by our previous report^7^ on global motion processing in the visual cortex, we focussed exclusively on this area of the brain. We found that a stronger functional connectivity between midline and far lateral areas of the visual cortex in gamma band during viewing of social scenes (smiling and singing women) was associated with a stronger discrepancy of lateral-to-midline activity during global motion processing, as well as with later presentation of autistic symptoms. Our results also revealed that infants’ midline-to-far lateral visual cortical functional connectivity in theta band during viewing of non-social scenes (colourful rotating toys) was positively associated with the extent of autistic symptoms later in toddlerhood. This association was strong in infants who had an elevated familial likelihood of developing autism, but non-significant in the low-likelihood group. Furthermore, among infants who later developed more autistic symptoms, this far lateral functional connectivity was seen to be stronger compared to infants who did not develop symptoms regardless of familial likelihood. We observed no association between alpha lateral connectivity, for either stimulus type, with later autism symptoms.
Consolidating our findings here with those from the previous study^7^(which involved mostly the same infants but with data from a different experiment), we have now converging evidence in support of atypical neural organization (as reflected in both SSVEP band power discrepancy and synchronized activation between midline and peripheral lateral areas) of the visual cortex of young infants who later present with significant autistic symptoms. In the previous study^7^, the stronger lateral relative to midline SSVEP power was seen during the processing of global motion (coherent moving dots). Here, we found stronger theta but weaker gamma midline-and-peripheral lateral synchronization during non-social scene processing and stronger gamma midline-and-peripheral lateral synchronization during social scene processing.
In infants’ visual cortex, global motion laterality was found to be correlated with far-lateral gamma connectivity during processing of social scenes, but not with far-lateral theta connectivity during processing of non-social scenes. These findings point to the possibility that laterality of non-social theta connectivity and laterality of social gamma connectivity/global motion processing reflect independent neural processes which are both linked to autism. Finally, the main model with only a handful of predictors representing visual cortical functional connectivity, i.e., non-social theta and both social and non-social gamma, could explain almost 50% of the variability in later symptom severity of autism.
Gamma connectivity in the visual cortex, links to global motion processing and autism
We found that the midline-to-far lateral visual cortical gamma connectivity was associated, albeit in different directions for social and non-social stimuli, with later autistic symptoms. These effects were only seen in the main-effect model and were not significant when moderation by the autism likelihood status was included.
Gamma rhythm activities in infancy have been linked to eye-gaze and dynamic face processing^33^, both of which have been reported to be different (from neurotypicals) in autism^34–36^. Atypical facial motion processing in autism has been thought to be of perceptual nature, which could originate from altered integration of cross-domain (i.e. spatial and motion) features^37^. Consistent with this, gamma rhythm has been linked to perceptual binding, needed to perceive connected parts as a single percept, and which has been shown to be present in 8-month-old infants’ EEG readings, with spatial distribution around the parieto-occipital region^38^. We found that the global motion laterality was positively linked to laterality of gamma connectivity during social scene processing, and both in turn were linked to later autistic symptoms. This may suggest that the two perceptual functions (global motion and facial motion perception) rely on some shared perceptual integrative processes that could be different in autism^39,40^. Altered unisensory and multisensory temporal integration have been well studied in adults and children with autism^41–43^. Nevertheless, the two functions were operationalized using different analytical constructs (i.e., phase-lagged connectivity vs. SSVEP power) each of which carries a different neurological meaning, which makes the inference about the nature of the association difficult (i.e., they can be different processes relying on the same underlying mechanism or just different aspects of the same process). It is noteworthy that in a previous related study^10^ we found the same global motion perception was associated with receptive language ability – a skill that could be affected by differences in facial motion perception – at 18 months (in largely the same infants).
A recent study^44^ suggested the existence of a third visual cortical processing pathway specialized for social perception, on top of the well-known dorsal and ventral pathways^45^, with ascribed functionality largely consistent with our interpretation here. In particular, the midline-to-far lateral functional connectivity used in our analysis could reflect the direct connectivity between V1 and MT/V5 reported by the authors and gamma rhythm synchronization between these two areas could serve as the underlying mechanism mediating social perception through visual motion integration^44^. Our results, thus, could provide information about the early development of this putative third pathway as well as insights to address its autism-related questions.
We found that gamma connectivity laterality for non-social stimuli was negatively associated with later symptom intensity. A general explanation for gamma connectivity results would be related to the neuronal inhibitory process, which are known to be indexed by the visual cortical gamma synchronization as seen in animal experiments^46^. The higher social gamma and lower non-social gamma connectivity associated with higher later autism symptoms might indicate an atypical balance of inhibitory processes in the visual cortex when responding to social and non-social stimuli^47^.
Strong lateral theta connectivity in infants with later symptoms of autism
Theta oscillations (3–5 Hz) within the occipital/occipitoparietal regions of infants aged 3–6 months^48,49^, measured via experimental manipulations, have been associated with infants’ emotional arousal^27,50,51^, reward experience^28^(also in visual cortex of animal models^52^, affection-inducing cognitive processing (e.g., during exploration of novelties^23^ or a social engagement^49,53^, and sustained anticipatory attention^29,54^. While these findings stemmed from different study designs compared to ours, it is possible that the spontaneous theta connectivity arising from watching the non-social video is linked to similar processes. Specifically, the higher non-social theta connectivity in the EL-High ADOS infants might reflect increased recruitment of affect-driven attentional resources^23,54^ during the processing of the colourful rotating toy video, consistent with evidence of stronger non-social responses from other studies using a similar paradigm but with different analytical or imaging modalities (e.g., band power^53^, ERPs^55^, and fNIRS^56,57^. Furthermore, using similar social/non-social stimuli as ours, a recent study^49^ showed that in typically-developing infants of age 9 months, a reconfiguration and expansion of the cortical network synchronized in theta band^58^ occurred, making it resemble the adult’s theta network^49^. Before such reconfiguration, theta functional connectivity showed no selectivity (modulation) to either types of stimulus^49^. In line with this, the theta connectivity responses of the LL and the EL-Low-ADOS groups showed similar (flipped) normalized topographical patterns for both stimulus types. In contrast, the midline-to-far lateral theta connectivity was stronger in response to non-social relative to social stimuli in the EL-High-ADOS infants. Note: No group difference was found in theta connectivity for the social stimulus. It is possible that this finding in the EL-high-ADOS group reflects early atypical selectivity in the cortical regions (e.g., lateral occipital and posterior temporal sites^59,60^ known to be selective for social stimuli which occurred already at age 5 months^61^.
Another possible explanation for our theta result in non-social stimulus condition relates to altered cortico-subcortical functional connectivity observed in autism^62,63^, that has been found to influence sensory information processing^64–66^. Increased excitatory connectivity from the thalamus to a posterior area of temporal lobe (close to MT/V5) has been found in autistic adults^65,67^. Because the thalamus also projects to V1, an elevated connectivity to the posterior temporal lobe in autism could lead to stronger synchronization between the midline and corresponding peripheral area of the visual cortex in this group. There is evidence of thalamic regulation of cortical sensory processing (and perceptual binding) via theta-gamma^68^ and theta-beta^69^ cross-frequency coupling. Nonetheless, the specific result we saw in infants during non-social stimulus viewing needs to be explained by further investigation.
Alpha connectivity and general attentional level to stimuli
We did not find any group or condition difference in the topographical alpha connectivity patterns. As alpha rhythm is associated with general attention and inactivity^30,70^, this may indicate that our attention coding and minimum epoch strategies worked well to ensure all infants, regardless of group, contributed data with the same level of general attentiveness to both stimulus types. Furthermore, the result also implied that the phenomena observed with both theta and gamma rhythms were not confounded by infants’ differential engagement behaviour during the session. Previous studies using similar paradigm as ours found alpha hyperconnectivity in the frontocentral scalp areas, which we did not assess here, in infants of age 14 months and found it was linked to the repetitive and restricted behaviour domain of autism^16,17^.
Finally, the link between autism and functional connectivity in general could reflect atypical cortical lateralization and asymmetry^4,71^ due to, among others, differences in early development of white matter tracts^72–75^. Yet it is notable that our results from 5-month-old infants showed the individual topographic connectivity patterns to be strongly unilateral, reflecting that the lateralization was highly individualized^76^(not consistently to one side of the brain).
Limitations
We did not include analysis of scalp areas associated with attention (e.g., posterior parietal, prefrontal) because the focus of our study was to follow up of our previous finding, linked to the visual cortex. Nonetheless, information about the general state of infants’ attention can be inferred from the variability of occipital alpha connectivity, which we found to be similar across experimental conditions and groups. Additionally, we have tried to minimize the risk that our analytical variables are impacted by overt attentional differences by implementing the attention coding and minimum epoch number strategies.
Our sample size suffered from high exclusion rate due to the minimum number of (≥ 90) epochs criterion which disproportionately excluded infants with high level autistic symptoms and/or an autism diagnosis. As a result, we did not have enough individuals diagnosed with autism to perform an analysis/modelling of autism diagnosis.
Although we labelled our two stimulus categories as social and non-social, a limitation of the current study is that these were not closely matched in terms of low-level perceptual properties. Both categories included rich, dynamic audiovisual content of equal duration and sound volume, but other features may have differed. Our intention was not to definitively test domain specificity, but rather to include two broad classes of stimuli that are both infant-relevant and infant-friendly. An advantage of this approach is that findings which generalize across such diverse stimulus categories may reflect highly robust effects, which is informative for understanding early cognitive processing. Finally, it is notable that the inherent limitations of EEG methods precluded us from drawing any conclusion about specific neuronal sources of the theta, alpha, and gamma oscillations observed in our study.
Conclusion
The link between global motion and social perception has been a longstanding question in autism research^77^. Here, we illuminated one possible link between the two perceptual functions by demonstrating a correlation between the laterality of global motion processing (represented by the Global Motion Laterality Score) and laterality of gamma connectivity during social stimulus viewing (represented by the Gamma-Social Far-Connectivity). Additionally, we found another independent neural process indexed by the laterality of theta connectivity during non-social stimulus viewing which was a strong predictor of later autism. This study and our previous report^7^ converge on highlighting the atypical functional organisation within the visual cortex as an important factor in early autistic development^9^ and the potential for this brain area for future biomarker discovery.
Methods
Participants
The EASE sample has been comprehensively described in past studies from our group^6,7,10^, thus we will only provide a cursory overview here. Participants were recruited via a direct phone call or adverts in paediatrics clinics around the Greater Stockholm area (for the Elevated Likelihood for Autism / EL infants) and from the Swedish Population Registry (for the Low Likelihood for Autism / LL infants). EL infants were determined from the presence of an older sibling, or any second-degree family member diagnosed with ASD. In addition to previously reported 55 infants^7^, 36th month ADOS-2 scores from additional 27 infants now became available as they reached suitable age to be clinically assessed. Detailed breakdowns of analysis-level exclusion (“data attrition”) are given in Supplementary Tables 1–2.
Ethics approval
All methods were performed in accordance with the relevant guidelines and regulations. Ethical permit for the EASE study was obtained in advance from the Stockholm Regional Ethics Committee.
Informed consent
All parents or legal guardians of the participating infants signed a written informed consent on behalf of their infants for study participation at the start of study.
Measures
General EEG setup
EEG recording was performed at one time-point only, when the participant was 5 months old, using the same EEG setup as reported previously^7^.
Visual stimuli
During the EEG session, infants watched various interchanging short video clips while sitting on their parents’ lap. A 1-minute clip showing a woman smiling and singing a Swedish nursery rhyme against a plain dark backdrop was used as our social-scene stimulus. Another 1-minute clip of two different colourful toys rotating around its axis shown one after another was used as the non-social stimulus. Both clips were played for three times in an interleaving manner, together with the form and motion stimulus videos^7^, during the 10-minute EEG session. Screenshots of both stimuli are shown in Figs. 1a, b and the video clips are included as Supplementary Videos 1–2 to this manuscript. Written informed consents were obtained from all our collaborators who acted as a model in our social stimulus video (shown in Fig. 1a and Supplementary Video 1) to be published in an open-access journal article.
Functional connectivity measure and frequency bands
Infants’ visual cortical functional connectivity was measured using dbWPLI^15^ due to its proven record for use with infants^16,17^, especially to cope with infants’ small head size that would aggravate the problems of spurious short-range connectivity. A dbWPLI score is calculated by first computing the cross-spectrum between waves measured at two different EEG electrodes and then estimating their mean phase difference (lag/lead) for a specific frequency. Mean phase difference for a range (“band”) of frequencies is calculated by averaging the scores over the frequencies. For the purposes of our study, we only take the absolute dbWPLI scores, which represent the strength of the connectivity but not the direction (i.e., leading or lagging).
Three frequency bands, adjusted for infancy^53,78^, were included in our analyses: theta (3–5 Hz), alpha (6–8 Hz), and whole gamma (20–60 Hz). These three brain rhythms were chosen due to their relevance to social cognitive development and autism in infants^22–26^. Therefore, together with the two stimulus types (social & non-social, see “Visual stimuli” above) we had 6 stimulus conditions (theta-social, theta-non-social, alpha-social, alpha-non-social, gamma-social, gamma-non-social) that were used in analyses.
Laterality score
We used the same Global Motion Laterality Score variable reported in our previous study^7^. This variable represents activation differences between midline area (presumably, corresponds to V1) and lateral area of the visual cortex^7^ and was calculated based on the T^2^-circular (T2circ) statistic^79^. The Global Motion condition corresponds to activation in response to coherent visual motion stimulus presented once every 0.5 s (2 Hz)^6,7^ to infants.
Assessment of autistic symptoms
A clinical assessment of infants’ autistic symptoms was performed at age of 36 months. As part of this assessment, ADOS-2 was administered to evaluate the severity of autistic symptoms by research-reliable clinicians. We used the total score of ADOS-2 CS as the outcome variable in a regression model (see “Analysis: Statistical modelling” below). The clinical diagnostic categorization (DSM-5) was based on a comprehensive assessment of all clinical information and consensus between two clinical experts, but as noted above only four children in the final sample received a formal DSM-5 diagnosis of autism spectrum disorder.
Analysis
Coding of infants’ attention to stimuli
We performed coding of infants’ attention to the visual stimuli from the recorded infant behaviour videos taken during the sessions. For each infant, using the Datavyu^80^ software, we manually marked segments of the video where the infant either (i) made gross movements (e.g., moving the whole body, an abrupt and large limb swing), or (ii) looked away from the presented stimulus, where in most cases both co-occurred. The obtained time markers were then used to exclude the corresponding EEG segments from the subsequent analysis. One infant was excluded from the beginning due to having no video.
Exclusion of bad EEG data
Due to equipment malfunction and other causes, two infants only had less than 1 min EEG data; the normal duration was around 10 min). These two infants were removed from further analysis.
EEG pre-processing
To pre-process the collected EEG data, we used the Harvard Automated Processing Pipeline for EEG (HAPPE)^81,82^ version 3.0 (https://github.com/PINE-Lab/HAPPE), that has been shown to provide a high data retention rate from developmental populations^83,84^. The pipeline runs on top of EEGLab^85^ and takes a raw EEG data (e.g., .RAW), processes it through various standardized steps (e.g., filtering, removing and interpolating bad EEG channels, re-referencing, segmenting, etc.) as well as applying more sophisticated techniques (e.g., 2-stage independent component analysis) to reject artefacts, and provides clean, segmented data (“epochs”) as its output^82^. Details of parameter settings used for running HAPPE in this study as well as the number of epochs produced for each processed participant’s data can be found in Supplementary Note 1. One participant was excluded due to having corrupted data and completely rejected by HAPPE.
Selection of visual cortex electrodes and definition of areas of interest
We inferred the EEG electrodes (within a 128-channel Geodesic Sensor Net HCGSN 130 [Electrical Geodesic Inc., OR]) that would approximately cover the entire visual cortex from past studies investigating the source localizations of EEG channels^86–88^ as well as studies of EEG functional connectivity in infants^16,17^. Twenty-four electrodes, symmetrically from left (12) and right (12) hemispheres, were selected from the occipital lobe that approximately corresponds to visual cortical areas relevant for our research questions^86,88^. The subsequent steps were generally inspired by approaches described in ref^18,19^ but adapted for use with EEG data. From this set of electrodes, we assigned two midline electrodes, Oz and POz, to define our seed area. Then, using the positional XYZ-coordinates generated by EEGLab for each electrode, we defined three areas of interest (AoIs) on each lateral side around the seed area (thus, a total of 6 AoIs) based on the Euclidean distance of each non-seed electrode to the average coordinates of the two seed electrodes. These AoIs were then assigned as areas at 1-, 2-, and 3-step distance away from the seed area, respectively, on left and right hemispheres. Please see Fig. 1c for a schematic diagram of these arrangements.
Computation of functional connectivity scores
Following the recommendations in refs^89,90^, we segmented the EEG data into 1-second 50%-overlapping epochs and, after rejecting epochs where the infant was not looking at stimuli (in more than 50% of an epoch’s length), retained only participants having at least 90 surviving epochs (equivalent to 1.5 min of uninterrupted data). Each 1-second epoch was tapered at the edges by a Hanning window to minimize “crosstalk” between successive epochs (especially so due to the 50%-overlap) in the subsequent dbWPLI score calculation. Thereafter, we applied a Fast Fourier Transform (FFT) to obtain the power spectrum from each epoch.
Next, we proceeded with a 4-step procedure to obtain our functional connectivity variables for modelling: (1) we computed a bivariate (electrode-to-electrode) dbWPLI score, as described in ref.^15^, between each of the 22 non-seed electrodes in our “visual cortex” to each of the two seeds; (2) one single functional connectivity score for each non-seed electrode was computed by averaging the two scores from the previous step; (3) a connectivity score for an AoI was computed by averaging the scores over all non-seed electrodes belonging to that AoI; (4) these steps were repeated for each of the six experimental conditions; see “Functional connectivity measure and frequency bands” above. See Fig. 2a and b for the plots of resulting individual and group-mean topographical dbWPLI patterns, respectively.
Vector-normalization and flipped topography
We applied a vector-normalization to each participant’s topographical (across the 6 AoIs) connectivity scores to focus on the relative patterns over the visual cortex and eliminate the influence of magnitude differences that can vary widely across or even within individuals, and to maintain consistency with the approach in ref^7^. This approach also had the effect of eliminating the very-high correlations (r ≥ .6) between the unnormalized AoI scores, which would render them practically unusable for regression modelling. Plots of these normalized topographical patterns are presented in Fig. 2c and d.
Next, to see whether the patterns of topographical connectivity involved bilaterally strong synchronizations between V1 and the lateral extrastriate areas, i.e., if the “peak” connectivity occurred in both hemispheres or only in one, we flipped the individual topographical patterns as such that all the (normalized) maximum scores lay on the same (right, chosen arbitrarily) side of the plot. Thus, for patterns having the maximum on left hemisphere, we rotated the entire patterns around the y-axis, while those having the maximum on right hemisphere were left unchanged. This approach, which was also used in ref^7^ as a supplementary analysis, revealed a strongly unilateral nature of the topographical connectivity, with much of the group differences visible on the hemisphere where the maximum occurred (see Fig. 2e and f). Consequently, we used these flipped patterns in all subsequent modelling and focussed only on the 3 AoIs from the (right) lateral side where the maxima are located. Although this decision removed any information about hemispheric specificity of the connectivity patterns, we considered it acceptable as it would play no role in addressing our research questions (see Introduction and also published as our analysis plan at https://osf.io/e8y5x).
Testing group differences
To infer group differences of the whole connectivity topography (all 6 AoIs included), we resorted to using permutation tests^91^ due to the highly-correlated nature of dbWPLI scores across visual cortex, even after vector-normalization and flipping. To do this, we shuffled the group labels (LL, EL-LoADOS, EL-HiADOS) and randomly assigned them among the participants, computed the difference in mean normalized dbWPLI scores for each pair of “new” groups in the 6 AoIs and 6 conditions (see “Functional connectivity measure and frequency bands”), and repeated the process for 10,000 times. The resulting distributions of normalized scores (in 6 AoIs and 6 conditions) were then tested against our data using their empirical quantiles. Consequently, this procedure performed three comparisons, one for each possible pairing of our three groups (see Fig. 3d and Supplementary Fig. 2).
To test differences in each of the 12 modelling variables (see “Statistical modelling” below) across the three participant groups, we used boxplots together with Kruskal-Wallis tests (see Fig. 3b and Supplementary Fig. 1).
Modelling association between functional connectivity and autism
For each frequency band (theta, alpha, gamma) and each stimulus condition (social, non-social), we took the 1-step distance AoI (on the maxima-side) scores as the “Near-Connectivity” variable and the 3-step distance AoI scores (on the maxima-side) as the “Far-Connectivity” variable, giving us a total of 12 modelling variables. We skipped using the 2-step distance AoI scores to further minimize any possible “crosstalk” between neighbouring scalp regions. For simplicity, we do not report the ratio of far-to-near connectivity, which we originally planned to do (https://osf.io/e8y5x), because eventually it did not provide any additional information beyond what can be provided by the Far- and Near-Connectivity combined. The resulting correlations among these 12 variables were more amenable (see Supplementary Note 1) for fitting simple ordinary least square (OLS) regression models (see below).
Subsequently, a non-parametric variable selection procedure based on cross-validation^55,92,93^ was run to identify a subset of the 12 variables that best explains the variability in ADOS-2 total score. A collection of all 4,096 (= 2^12^) possible subsets of the 12 variables, including the empty set and full set, was generated and then an OLS linear regression model with ADOS-2 total score as the outcome was fitted on each of these subsets as regressors. For each fitted model, we performed a 10-fold cross-validation (CV) with 10 repetitions (each time generating a different random 9:1 split of the dataset) to obtain an average coefficient of determination (R^2^) score, selected 100 models with the best R^2^ scores, and ranked the frequency of occurrences of each variable in these 100 models. The top 5 variables were then fitted in the final linear regression model for inferring the association with ADOS-2 total score.
Relation between functional connectivity and global motion processing
To infer whether variance (of ADOS-2 total score) explained by the dbWPLI variables can be explained by the Global Motion Laterality Score, we added the latter to the existing linear model of ADOS-2 total score vs. functional connectivity (see “Modelling association between functional connectivity and autism”) as an extra predictor. Then, we (i) noted how the sum-of-square (SS) of each dbWPLI predictor changed and (ii) tested whether the resulting increase (if any) of the model’s R^2^ is significant. For (i), we performed an analysis of variance (ANOVA) on the model before and after addition of the extra predictor, and for (ii) a nested likelihood ratio test on the same two models was done. A substantial overlap in variances explained by Global Motion Laterality Score and any of the dbWPLI predictors would point to a possibility that the lateral activation we saw in the previous study of global motion processing^7^ has a basis in and/or an implication for functional connectivity in the brain rhythms studied here. Finally, to better understand the nature of this hypothesized relation between global motion processing and visual cortical functional connectivity in the three brain rhythms, we fitted an OLS linear regression of Global Motion Laterality Score regressing on dbWPLI predictors. Before the model-fitting, we again ran the same non-parametric variable selection procedure as before (see “Modelling association between functional connectivity and autism”), but now with Global Motion Laterality Score as the outcome, to trim down the number of plausible dbWPLI predictors to include in the model from 12 to only 5. The eventual model fitting included only the selected 5 dbWPLI predictors and no covariates, because sex, age (at 5 months), and likelihood for autism status had been regressed out from all dbWPLI variables and Global Motion Laterality score.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
Supplementary Material 3
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gandal, M. J. et al. Broad transcriptomic dysregulation occurs across the cerebral cortex in ASD. Nature, 611, 532–539 (2022).10.1038/s 41586-022-05377-7PMC 966874836323788 · doi ↗ · pubmed ↗
- 2Falck-Ytter, T. & Bussu, G. The sensory-first account of autism. Neuroscience Biobehavioral Reviews 153 105405 (2023).10.1016/j.neubiorev.2023.10540537742990 · doi ↗ · pubmed ↗
- 3Hedenius, M., Hardiansyah, I. & Falck-Ytter, T. Visual global processing and subsequent verbal and non-verbal development: An EEG study of infants at elevated versus low likelihood for autism spectrum disorder. Journal autism Dev. disorders, 53, 3700-3709 (2023).10.1007/s 10803-022-05470-w PMC 1046565935353335 · doi ↗ · pubmed ↗
- 4Lord, C. et al. Autism diagnostic observation schedule–2nd edition (ADOS-2). Los Angeles CA: Western Psychol. Corporation 284 (2012).
- 5Nakano, T., Ota, H., Kato, N. & Kitazawa, S. Deficit in visual temporal integration in autism spectrum disorders. Proceedings of the Royal Society B: Biological Sciences 277, 1027–1030 (2010).10.1098/rspb.2009.1713 PMC 284275619955150 · doi ↗ · pubmed ↗
- 6Team, D. & Datavyu A video coding tool. Databrary Project, New York University. URL http://datavyu. org (2014).
- 7Monachino, A., Lopez, K., Pierce, L. & Gabard-Durnam, L. The HAPPE plus event-related (HAPPE + ER) software: a standardized processing pipeline for event-related potential analyses. Bio Rxiv, 2007. 2002.450946. (2021).10.1016/j.dcn.2022.101140 PMC 935614935926469 · doi ↗ · pubmed ↗