Influence of egg traits on parasitism by Trichogramma chilonis Ishii, 1941 and Telenomus remus Nixon, 1937 against Spodoptera frugiperda (J.E. Smith, 1797)
Kushal Giri, Min Raj Pokhrel, Ghanashyam Bhandari

TL;DR
This study compares how two parasitoid species affect the fall armyworm's eggs, finding that Telenomus remus is more effective than Trichogramma chilonis, especially in older or covered eggs.
Contribution
The study provides new insights into the effectiveness of two parasitoid species against different egg traits of Spodoptera frugiperda.
Findings
Telenomus remus showed significantly higher parasitism rates than Trichogramma chilonis in both single and multilayered egg masses.
Parasitism rates decreased with increasing host egg age for both parasitoid species.
Telenomus remus had a higher adult emergence percentage regardless of egg scale coverage.
Abstract
Trichogramma chilonis Ishii, 1941 and Telenomus remus Nixon, 1937 are the most common egg parasitoids of fall armyworm (FAW), Spodoptera frugiperda (J.E. Smith, 1797) in maize growing areas. FAW lays single to multilayered egg which are covered with degrees of scale thickness. Here, we assessed the parasitism of both parasitoids over different FAW egg densities (single layered), egg mass scale coverage (multilayered), and the egg ages. Two laboratory experiments were conducted from May to November 2022 under controlled conditions (24.3 ± 0.8 °C, 69.3 ± 2.2% RH). The first experiment, using a three-factor complete randomized design (CRD), involved two egg parasitoid species, single-layered eggs at three densities (20, 43, and 60 eggs), and three egg age groups (less than 12 hours, 24–36 hours, and 48–60 hours), each replicated three times. The second experiment involved two parasitoid…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
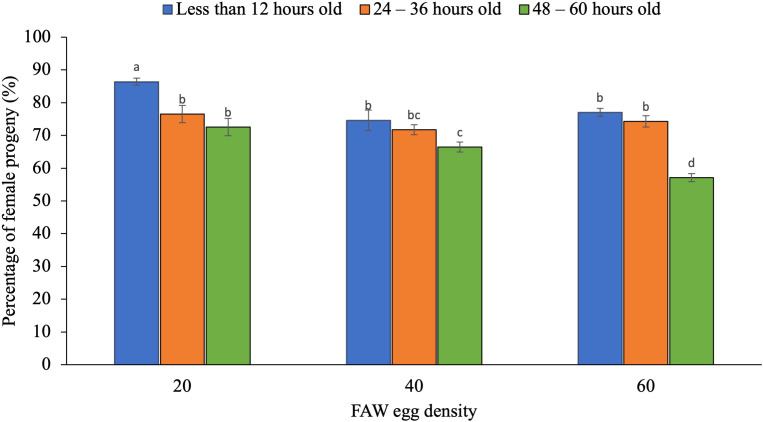 Figure 1
Figure 1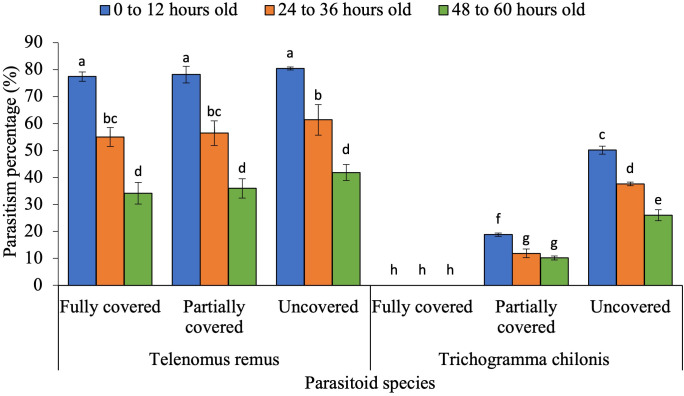 Figure 2
Figure 2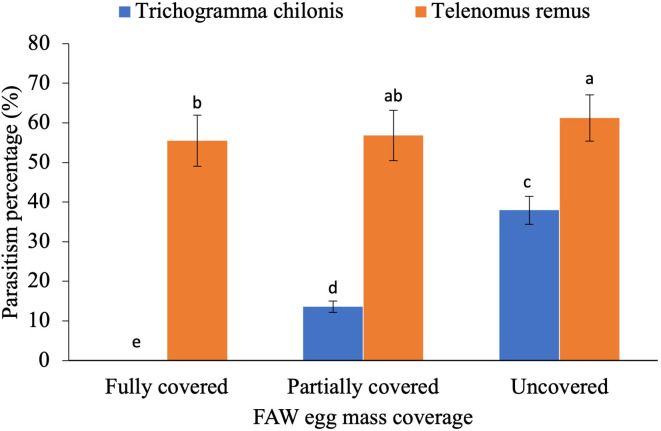 Figure 3
Figure 3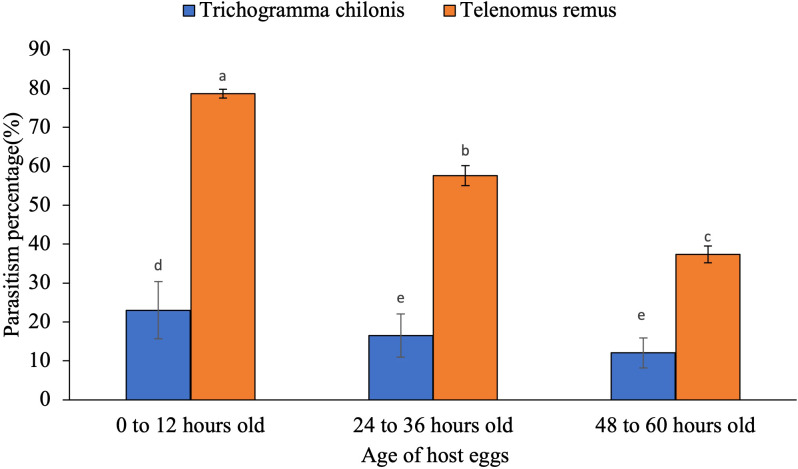 Figure 4
Figure 4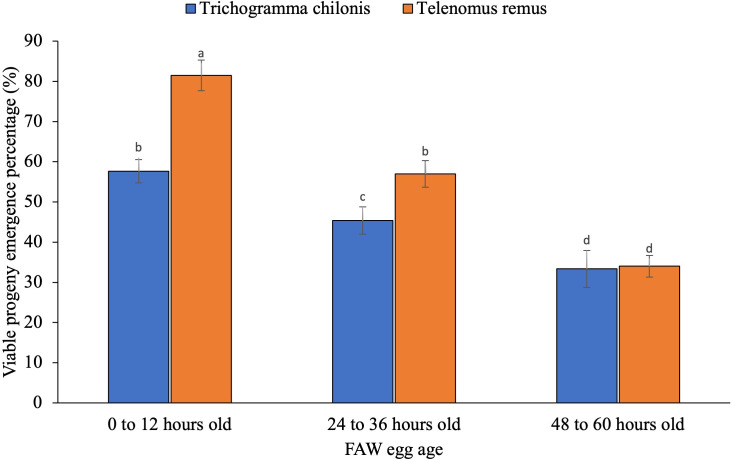 Figure 5
Figure 5| Treatments | Parasitism (%) | Viable parasitoid emergence (%) | Developmental duration (days) |
|---|---|---|---|
| Parasitoids | |||
|
| 63.95b ± 3.23 (53.72) | 69.52b ± 3.24 (57.21) | 9.8 ± 0.1 |
|
| 75.83a ± 2.65 (62.12) | 81.05a ± 2.16 (65.53) | 9.7 ± 0.1 |
| SEM (±) | 0.70 | 1.34 | 0.09 |
| MSD | 2.00 | 3.83 | 0.26 |
| F-test | 72.49* | 19.42* | 0.08 NS |
| Egg density | |||
| Twenty eggs | 81.67a ± 2.89 (66.71) | 84.97a ± 1.96 (68.38) | 9.8 ± 0.1 |
| Forty eggs | 68.75b ± 3.14 (56.48) | 75.28b ± 3.33 (61.10) | 9.9 ± 0.1 |
| Sixty eggs | 59.26c ± 3.64 (50.58) | 65.60c ± 3.81 (54.77) | 9.6 ± 0.1 |
| SEm (±) | 0.85 | 1.64 | 0.11 |
| MSD | 2.95 | 5.66 | 0.39 |
| F-test | 91.29* | 17.65* | 1.58NS |
| Egg age | |||
| 0–12 hours old | 83.24a ±2.49 (67.68) | 85.03a ± 1.82 (69.93) | 9.9 ± 0.1 |
| 24–36 hours old | 69.81b ± 3.01 (57.15) | 72.18b ± 4.25 (59.90) | 9.8 ± 0.1 |
| 48–60 hours old | 56.62c ± 3.20 (48.93) | 68.65b ± 3.22 (56.41) | 9.6 ± 0.1 |
| SEm (±) | 0.85 | 1.64 | 0.11 |
| MSD | 2.95 | 5.66 | 0.39 |
| F-test | 121.04* | 12.73* | 2.58NS |
| CV (%) | 6.26 | 11.31 | 4.83 |
| Grand mean | 69.89 | 75.28 | 9.76 |
| Treatments | Percentage of | Developmental |
|---|---|---|
| Parasitoids | ||
|
| 70.56 ± 1.46 (57.25) | 9.89 ± 0.18 |
|
| 73.18 ± 1.69 (58.99) | 9.72 ± 0.11 |
| SEm (±) | 0.73 | 0.15 |
| MSD | 2.13 | 0.44 |
| F-test | 2.85NS | 0.60NS |
| Scale covering | ||
| Fully covered | – | – |
| Partially covered | 72.66 ± 1.33 (58.57) | 9.67 ± 0.16 |
| Uncovered | 71.08 ± 1.83 (57.65) | 9.94 ± 0.13 |
| SEm (±) | 0.73 | 0.15 |
| MSD | 2.13 | 0.44 |
| F-test | 0.81 NS | 1.67NS |
| Egg age | ||
| 0–12 hours old | 75.49a ± 1.44 (60.43) | 9.83 ± 0.21 |
| 24–36 hours old | 74.83a ± 0.93 (59.93) | 9.91 ± 0.19 |
| 48–60 hours old | 65.30b ± 1.79 (53.99) | 9.67 ± 0.14 |
| SEm (±) | 0.89 | 0.19 |
| MSD | 2.61 | 0.54 |
| F-test | 16.1* | 0.47NS |
| CV (%) | 5.32 | 6.58 |
| Grand mean | 71.87 | 9.80 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Insect Resistance and Genetics · Insect Pheromone Research and Control
Introduction
1
The fall armyworm (FAW), Spodoptera frugiperda (J.E. Smith, 1797), is an invasive pest species native to tropical and subtropical regions of the America (1). Until 2015, FAW restricted to its native range (2). Its outbreak was first recorded in Africa in early 2016 (1) and in India in 2018 (3). Then, it was officially documented in Nepal in 2019 in the Nawalpur district (4). This polyphagous and devastating nature of FAW threatens maize production systems and national food security (5). In Nepal, FAW has the potential to cause 20-25% damage to maize, resulting in a loss of approximately 0.5 million tons of total maize production, valued at around 200 million dollars (6). Given the conducive climatic conditions, uncontrolled FAW infestation could completely decimate maize crops (7). Various methods exist to tackle the FAW problems in maize at farmers level with major dominance of chemical measures (8, 9). Their overuse has adverse effect on the environment, non-target organisms, and human health (10). Moreover, it triggers problems like resistance development, resurgence of pests, and the destruction of natural enemies (11). Thus, there is a global need for environmentally friendly alternatives to address the FAW problem without harming the environment and human health (12).
Biological control is an eco-friendly approach to manage FAW (13). Being a self-propelling and self-perpetuating system, this method proves to be economic and sustainable in the long run (12). It contributes as an important component of the Integrated Pest Management (IPM) approach (14). This method in combination with various other control methods carries the potential to significantly decrease the amount of pesticides being used currently (15). It makes a use of predators, parasitoids, entomopathogens, and biopesticides that attack on specific pest stages (12). The use of natural enemies like parasitoids (egg and larval), predators and microorganisms, can be the best alternative to chemical insecticides for FAW suppression (2). Egg parasitoids can control pest which makes them a potential candidate in biological control (16). Besides, they can be easily mass-produced in small area using both natural and laboratory host (17). Field surveys at various parts of the world have recorded the presence of egg parasitoids (Trichogramma spp., and Telenomus spp.), larval parasitoids (Cotesia spp., Charops spp., and dipteran species), egg-larval parasitoid (Chelonus curvimaculatus), as well as entomopathogenic fungi (Beauveria bassiana and Metarhizium anisopliae) with proven efficiency against FAW (18–21). These biocontrol agents are easy to rear in the laboratory and suitable for field release (14, 22). Among different biological control agents, eggs parasitoids are mostly explored and used against major agricultural pests (23). Among them, Trichogramma chilonis Ishii, 1941 and Telenomus remus Nixon, 1937 are proven to be the most promising egg parasitoids for FAW management (24) and have been recorded in different parts of Nepal (25, 26).
Parasitism by egg parasitoids is influenced by various traits of host eggs (27). Egg density affects parasitism by influencing encounter rates with small to moderate densities enhancing the efficiency, whereas higher density reducing it due to competition and superparasitism (28, 29). Host egg age also influences parasitoid preference and outcomes—fresh host can lead to higher parasitism whereas older eggs with hardened chorion and developed embryo may limit parasitoids fitness (30, 31). Scale covering on FAW egg massess acts as a physical barrier for the parasitoids like T. chilonis having short ovipositor (24), whereas T. remus are known for its ability to penetrate dense, scaly and multilayered egg masses (13).
Several studies have explored the parasitism of egg parasitoids under different conditions (13, 29, 32), however, a comprehensive study of how these host egg traits—egg density, scale cover, and egg age—influence parasitism remains limited. Moreover, comparative assessments between T. chilonis and T. remus under these conditions are scarce, despite their co-existence and overlapping use in FAW biocontrol programs. In this study, we systematically assessed and compared the parasitism, adult emergence, percentage of female progeny, and developmental duration of T. chilonis and T. remus across different densities, scale covering, and age groups of FAW eggs under laboratory conditions.
Materials and methods
2
Experimental location
2.1
The experiments were conducted during the maize-growing season (May–November 2022) under controlled laboratory conditions (mean temperature 24.3 ± 0.8 °C and RH 69.3 ± 2.2%) at the Entomology Laboratory of the National Maize Research Program (NMRP), Rampur, Chitwan, Nepal (27°39′17″N, 84°21′2″E; 228 m above sea level).
FAW colony establishment
2.2
Late-instar S. frugiperda larvae were collected from pesticide-free maize fields at NMRP and reared on fresh maize leaves in transparent plastic containers (19 x 14 x 7 cm^3^) for cohort-based rearing (33). Fresh green leaves were provided every one to two days, depending on their condition, until the larvae pupated. The pupae were collected daily and transferred to separate boxes (14 × 10 × 4 cm³). This rearing process was repeated to ensure a continuous supply of pupae for adult emergence.
Egg collection chamber
2.3
The adults after emergence were transferred to oviposition cages (30 × 30 × 30 cm3) supplied with water-soaked cotton, honey, and young maize plants for egg laying. The next morning, maize plants with eggs on their leaves were carefully removed. Leaves with egg masses were clipped and placed in a well-ventilated box. Medium-sized multilayered egg masses (roughly 100 eggs) were then selected and categorized into three groups based on scale covering: fully covered, partially covered, and uncovered. Similarly, single-layer egg masses were counted under a microscope using a fine camel hairbrush, and batches of 20, 40, and 60 eggs were prepared. This process was repeated every day to obtain eggs of different ages for each group. Finally, the eggs were glued onto cardboard pieces (3 × 5 cm²) to prepare standardized egg cards for the experiment.
Laboratory rearing of egg parasitoids
2.4
A wild population of T. remus were collected from the NMRP maize field and a nucleus colony was established in the laboratory. FAW egg masses were randomly collected and checked for the presence of parasitoids. The unparasitized eggs yields only FAW larvae whereas, parasitized eggs produced few or no FAW larvae after 1–3 days and adult parasitoids emerged after about 7–10 days. The emerged parasitoids were supplied with a thin layer of honey as source of energy and later identified to confirm the species. To multiply them, fresh FAW egg masses (from egg collection chamber) were irradiated in a UV chamber for 15–20 minutes to halt the development of embryo. The egg cards were then exposed to the test tube containing parasitoids at a ratio of one parasitoid per 20 eggs for 24 hours, with fresh FAW eggs supplied on alternate days until the parasitoids died, as described by Tefera et al. (2019) (14). The parasitized egg masses were kept in separate test tube, and the process was repeated for 3–4 generations. The parasitized egg masses (master’s card) were prepared and refrigerated at 4–6 °C for 1–2 days at parasitoid pupal stage. These master cards were then taken out each day to make sure that freshly emerged adult parasitoids are available for research every day.
The other species of egg parasitoid, T. chilonis was being successfully reared in the entomological division of NMRP, Rampur. Tricho-master cards (card with glued eggs of Corcyra cephalonica (Stainton, 1866) already parasitized by T. chilonis) were used to parasitize few fresh batches of FAW eggs. The parasitized egg masses were then used to parasitized and multiply other batches of eggs as explained for T. remus and after 3–4 generations, multiple batches were made and refrigerated to ensure a continuous supply of new parasitoid every day in the laboratory.
Parasitism at different egg densities and ages (Experiment 1)
2.5
In nature, egg parasitoids come across FAW eggs in different numbers and at different ages. The number of eggs depends on the age of the female FAW—newly mated females lay many eggs, while older ones lay fewer. FAW eggs also hatch in about 3 days, so their age varies depending on the oviposition time. To study how these differences in egg number and age affect parasitism, two egg parasitoids—T. chilonis and T. remus—were tested on FAW eggs of three different densities (20, 40, and 60 eggs) and three age groups (less than 12 hours, 24–36 hours, and 48–60 hours old). The experiment was set up using a completely randomized design (CRD) with three replications. For each test, a one-day-old mated female parasitoid was placed inside a glass test tube (15 mm × 100 mm). Data were collected on the proportion of parasitism, viable adult emergence, sex ratio (percentage of female progeny), and the developmental period of both parasitoid species.
Parasitism at different egg scale covering and ages (Experiment 2)
2.6
Newly mated female lays egg mass with fully covered scale and, the proportion of scale decreases and old female lays eggs without scale covering. To study how these differences in egg scale covering and age affect parasitism, T. chilonis and T. remus were evaluated on FAW eggs of three different scale covering (Fully covered, partially covered, and uncovered) and three age groups (less than 12 hours, 24–36 hours, and 48–60 hours old). The experiment was also set up using a completely randomized design (CRD) with three replications. Uniform sized egg masses (nearly 100 ± 10) were kept in a test tube and a one-day-old mated female parasitoid was placed inside a glass test tube (15 mm × 100 mm). Data were collected on the proportion of parasitism, viable adult emergence, sex ratio (percentage of female progeny), and the developmental period of both parasitoid species.
Statistical analysis
2.7
All data were entered into MS Excel (Office 365) for basic analysis and graph generation. Normality and homogeneity were tested using the GVLMA package, and data not meeting ANOVA assumptions were arcsine-transformed. ANOVA was conducted in RStudio (v2022.03.1), and significant results were separated using Tukey’s HSD at a 5% significance level. Univariate outliers present on the datasets were identified using box and whisker plot techniques. The influential outliers compromising and questioning the credibility of the result were handled by the variable deletion techniques i.e., data exclusion as explained by Mowbray et al. (2019) (34). Data on percentage emergence of parasitoids, female progeny percentage and developmental period for experiment 2 were compromised because of the undefined value of T. chilonis on one variable (fully covered). The variable was excluded for the unbiased estimate of those parameters.
Results
3
Parasitism across egg densities and host egg age
3.1
Parasitism
3.1.1
Parasitism was significantly influenced by the parasitoid species, host egg density and egg age (Table 1). Across all egg densities and egg ages, T. remus parasitized 75.8% of FAW eggs, which was statistically higher than T. chilonis, which parasitized 63.9% of the FAW eggs (F_1,36_ = 72.49; p < 0.05). Parasitism was also influenced by egg density. Egg masses with a density of 20 eggs had the highest parasitism (81.7%), which was significantly higher than those with 40 eggs (68.8%) and 60 eggs (59.3%) (F_2_,36 = 91.29; p < 0.05). Host egg age directly affected parasitism, regardless of the parasitoid species. Eggs less than 12 hours old had significantly higher parasitism (83.24%), followed by eggs aged 24–36 hours (69.8%), and 48–60 hours (56.6%) (F_2_,36 = 121.04; p < 0.05).
Viable parasitoid adult emergence
3.1.2
The percentage of viable parasitoid adult emergence was significantly influenced by the parasitoid species, egg density, and the egg age (Table 1). T. remus showed higher adult emergence (81.1%) compared to T. chilonis (69.5%) (F_1,36_ = 19.42; p < 0.05). Emergence percentages were highest in egg masses of 20-egg density (84.9%), followed by 40-egg density (75.3%) and 60-eggs (65.6%) (F_2,36_ = 17.65; p < 0.05). Similarly, emergence was significantly higher in eggs less than 12 hours old (85.03%), followed by eggs aged 24–36 hours (72.18%) and 48–60 hours (68.65%) (F_2,36_ = 12.73; p < 0.05).
Sex ratio (Percentage of female progeny)
3.1.3
The average percentage of emerged female parasitoids in T. chilonis (72.73%) was statistically similar to that of T remus (73.20%) (F_1,36_ = 0.19; p = 0.67). The percentage of female was significantly influenced by the interaction between egg density and egg age (Figure 1; F_4,36_ = 3.78; p = 0.01).
Percentage of female progeny of egg parasitoids as influenced by interaction between host egg density and egg age. The standard error bar with the same letter (s) over the bar are not significantly different from each other (Tukey HSD test, α = 0.05)
Developmental period
3.1.4
The developmental duration of the parasitoids was not significantly affected by parasitoid species, egg density, or egg age (Table 1). The developmental period of T. chilonis was 9.8 days and 9.7 days for T. remus (F_1,36_ = 0.08; p = 0.77). For both species, adult emerged after 9.8 days from 20-egg density, 9.9 days from 40-egg density and 9.6 days from 60-egg density (F_2,36_ = 1.58; p = 0.22). Parasitoids emerged after 9.9 days from less than 12 hours old egg FAW eggs, 9.8 days from 24–36 hours old eggs and 9.6 days from 48–60 hours old eggs (F_2,36_ = 2.58; p = 0.09).
Parasitism across egg scale covering and host egg age
3.2
Parasitism
3.2.1
Parasitism was significantly influenced by the three-way interaction among the parasitoid species, host egg scale coverage, and host egg age (Figure 2; F_4,36_ = 3.56, p = 0.02). The parasitism percentage was also affected by the interaction between parasitoid species and host egg scale covering (Figure 3; F_2,36_ = 160.46, p < 0.001), as well as between parasitoid species and egg age (Figure 4; F_2,36_ = 43.06, p < 0.05).
Parasitism percentage of egg parasitoids as influenced by interaction between parasitoid species and host egg age. The standard error bar with the same letter (s) over the bar are not significantly different from each other based on (Tukey HSD test, α = 0.05).
Parasitism percentage of egg parasitoids as influenced by interaction between parasitoid species and host egg scale covering. The standard error bar with the same letter (s) over the bar are not significantly different from each other (Tukey HSD test, α = 0.05).
Parasitism percentage of egg parasitoids as influenced by interaction between parasitoid species, host egg scale covering and host egg age. The standard error bar with the same letter (s) over the bar are not significantly different from each other (Tukey HSD test, α = 0.05).
Viable parasitoid adult emergence
3.2.2
The percentage of viable adult emergence was significantly influenced by the interaction between parasitoid species and host egg age (Figure 5; F_2,24_ = 6.22; p = 0.007) but not influenced by the egg scale covering (F_1,24_ = 3.34; p = 0.08).
Viable parasitoid adult emergence percentage as influenced by interaction between parasitoid species, and host egg age. The standard error bar with the same letter (s) over the bar are not significantly different from each other (Tukey HSD test, α = 0.05).
Sex ratio (Percentage of female progeny)
3.2.3
The percentage of emerged female was 73.19% for T. remus and 70.56% for T. chilonis, with no significant difference (Table 2) (F_1,24_ = 2.85; p = 0.10). Similarly, the percentage of emerged female was 72.66% in partially covered egg masses and 71.08% in uncovered egg masses for both parasitoid species, which were also statistically similar (F_1,24_ = 0.81; p = 0.38). However, egg age positively affected the emerged female. Eggs less than 12 hours old (75.49%) and 24–36 hours old (74.83%) produced significantly higher females than 48–60 hours old eggs (65.30%) (F_2_,24 = 16.10; p < 0.05).
Developmental period
3.2.4
There was no significant influence of parasitoid species, FAW egg mass coverage and ages on developmental duration of parasitoids (Table 2). The developmental days of T. chilonis was 9.9 days and for T. remus was 9.7 days. (F_1,24_ = 0.60; p = 0.45). Similarly, the developmental duration was 9.9 days for uncovered eggs and 9.7 days for partially covered eggs for both parasitoid species (F_1,24_ = 1.67; p = 0.21). Likewise, parasitoids emerge after 9.8 days in less than 12 hours old eggs, 9.9 days in 24–36 hours old eggs, and 9.7 days in 48–60 hours old eggs (F_2,24_ = 0.47; p = 0.63).
Discussion
4
Parasitism
4.1
This study aimed to evaluate the parasitism efficiency of T. remus and T. chilonis on S. frugiperda egg masses of varying structures and ages. Our finding demonstrated that T. remus exhibited significantly higher parasitism in both single and multilayered egg masses. Parasitism was higher for T. remus than T. chilonis in both single and multilayered egg masses, with the highest rates in single layered egg mass due to easier accessibility for parasitoids (28). The parasitism declines with egg mass layers and scale covering, reflecting the physical challenge of overcoming scales and penetrating deeper layers. These findings are supported by Beserra and Parra (2005) (28), Jin et al. (2021) (29), Mohamed (2021) (35) and Mohamed et al. (2023) (36), all of them confirmed the parasitism rates by different Trichogramma species decreased based on the number of egg mass layer.
The superior performance of T. remus is largely due to its robust and aggressive nature enabling it to overcome the physical barrier of scale coverings and penetrate deeper into multilayered egg masses (13). In contrast, T. chilonis was hindered by the scale covering, restricting it to parasitize only the upper layers or eggs on the edges (24, 30).
Parasitism decreased with increasing host egg age due to depletion of nutrient and host embryo development, which rendered older eggs unsuitable for parasitoid development (31). Also, the egg scale covering harden with age, making it more challenging for parasitoids to penetrate through it (37). Although these results demonstrate T. remus’s superiority over T. chilonis as a biocontrol agent, field complexities may not be fully captured by laboratory conditions. Future studies should evaluate their parasitic performance under natural conditions and investigate combined control methods that use both parasitoids.
Parasitoid adult emergence
4.2
This study investigated the emergence success of T. remus and T. chilonis from S. frugiperda egg masses of varying structures and ages. The results revealed significantly higher adult emergence percentage for T. remus compared to T. chilonis, which may be due to its stronger adaptation to S. frugiperda eggs. These findings are supported by Carneiro and Fernandes (2012) (24), who also documented higher emergence rate of T. remus than T. pretiosum on similar host systems.
Parasitoids emergence is influenced by host egg age. As host egg age increased, emergence rates declined, indicating that older eggs likely became less suitable for parasitoid development. This is likely due to nutrient depletion and progressive host embryo development in older eggs, which fail to support parasitoid growth and lead to higher mortality before emergence —a pattern also observed by Tuncbilek and Ayvaz (2003) (31). The findings are also supported by Priyanka et al. (2023) (32), who reported a decrease in parasitoid emergence with increasing host egg age. The emergence rates of both species were not significantly impacted by egg scale covering, suggesting this physical barrier does not hinder larval development after oviposition. This finding is supported by Laminou et al. (2020) (13) and Mohamed et al. (2023), who reported no effects of FAW egg scale coverage on emergence of egg parasitoids. These results imply that fresh or young host eggs are suitable for mass rearing of parasitoids. Additionally, T. remus seems to be a good option for field release against S. frugiperda, particularly in cases where host egg age varies. Future research should validate these findings in the field and look into possible explanations for the developmental advantages of T. remus offspring over those of other species.
Sex ratio (Percentage of female progeny)
4.3
This study investigated the influence of host egg density, egg scale covering and egg age on the percentage of female progeny in two egg parasitoids, T. chilonis and T. remus. Our study reveals that the percentage of female progeny in both species of egg parasitoids are statistically similar. This finding is in consistent with the finding of Jin et al. (2021) (29) and Chen et al. (2021) (38), who too reported the average percentage of female progeny in T. chilonis and T. remus around 70 percent.
The proportion of female offspring decreased with increasing host egg densities. This might be explained by the fact that more unfertilized S. frugiperda eggs were parasitized in high-density environments, which probably produced more male progeny (29). Additionally, the percentage of female progeny declined with increasing egg age. This may be possibly due to declining egg quality and reduced resources in older eggs, which negatively impact the development of female parasitoids. This is supported by findings from Sun et al. (2021) (30), who observed a drop in Trichogramma female progeny percentage from 82% to 69% as egg age increased from 0 to 2 days. Similarly, Priyanka et al. (2023) (32) noted a reduction in T. remus female progeny from 66% to 64% as egg age increased from 24 to 48 hours. Our results demonstrate that the sex ratio of emerging parasitoids is significantly influenced by the age and density of the host egg. Because more females can increase the success of parasitism, this is beneficial for mass-rearing and biocontrol initiatives. These effects should be tested in other host-parasitoid systems and in the field.
Developmental period of parasitoids
4.4
This study investigated the influence of host egg density, egg scale covering and egg age on the developmental duration of two egg parasitoids, T. chilonis and T. remus. Our finding reveals that the developmental time from egg to adult remained consistent across both T. chilonis and T. remus, regardless of treatment conditions. This is supported by findings of Sultan et al. (2013) (39) who reported a development period of 9.60 days for T. chilonis, while Oktaviani et al. (2021) (40), observed a similar duration of 9.61 days for T. remus.
The reason behind this stability in developmental time may lie in the biology of the parasitoids. As noted by Bueno et al. (2008) (41) these parasitoid species lay their eggs singly inside individual FAW eggs and superparasitism or multiple parasitism are rare (42). Therefore, each parasitoid larva develops independently within a single host egg, external factors such as egg density or the presence of scale covering do not appear to influence their development. Moreover, our findings are consistent with Priyanka et al. (2023) (32), who recorded developmental durations of T. remus as 9.61 days on 24-hour-old eggs and 9.52 days on 48-hour-old eggs. These results further support the conclusion that the age of the host egg does not significantly impact the time required for parasitoid development. Although the developmental duration of these two parasitoids is consistent across different egg traits, the small sample size and laboratory setting necessitate larger field-based research to validate and generalize the findings.
Conclusions
5
The higher parasitism of T. remus on multilayered and scaly egg masses of S. frugiperda indicates its potential as biological control agent for scaly egg masses. However, T. chilonis fails to overcome the layers and scales barrier of eggs of S. frugiperda. The parasitism of both parasitoid species decreases with the increasing age of host eggs, highlighting the importance of synchronization in augmentative biological control programs. For both species, laboratory mass production should coincide with the freshly laid fall armyworm eggs (within 24 hours) for efficient parasitism and sustainable female populations in the environment.
However, this study was conducted in lab condition using single female in single tube setup, which may not fully represent actual field situation. In field condition, parasitism is shaped by competition, host searching behavior, and heterogenous egg masses all of which can change parasitism compared to laboratory conditions. In addition, host egg scale coverage was measured qualitatively (fully covered, partially covered, uncovered) rather than quantitatively. Because “partial” versus “full” coverage can be subjective, and scale thickness may vary among egg masses, future studies should quantify scale coverage and thickness and validate parasitoid performance under semi-field and field conditions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Goergen G Kumar PL Sankung SB Togola A Tamo M . First report of outbreaks of the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. Plo S One. (2016) 11:1–9. doi: 10.1371/journal.pone.0165632, PMID: 27788251 PMC 5082806 · doi ↗ · pubmed ↗
- 2Ogunfunmilayo AO Kazeem SA Idoko JE Adebayo RA Fayemi EY Adedibu OB . Occurrence of natural enemies of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) in Nigeria. Plo S One. (2021) 16:1–13. doi: 10.1371/journal.pone.0254328, PMID: 34252127 PMC 8274871 · doi ↗ · pubmed ↗
- 3Sharanabasappa D Kalleshwarmy CM Asokan R Swamy HM Maruti MS Pavithra HB . First report of the Fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manage Hortic Ecosys. (2018) 24:23–9.
- 4Bajracharya ASR Bhat B Sharma P Shashank PR Meshram NM Hashmi TR . First record of fall armyworm Spodoptera frugiperda (J. E. Smith) from Nepal. Indian J Entomol. (2019) 81:635–9. doi: 10.5958/0974-8172.2019.00137.8 · doi ↗
- 5Ghimire P Bhetwal N . FAW: An emerging threat to agriculture. Food Agri Econ Rev. (2021) 1:93–100. doi: 10.26480/faer.02.2021.93.100 · doi ↗
- 6Pradhan B . Collective efforts to fight fall armyworm in Nepal (2020). Available online at: https://www.cimmyt.org/news/collective-efforts-to-fight-fall-armyworm-in-Nepal/ (Accessed April 7, 2020).
- 7Sah LP Lamichhaney D KCHB Acharya MC Humagain SP Bhandari G . Fall armyworm (Spodoptera frugiperda) in maize: Current status and collaborative efforts for its management in Nepal. J Plant Prot Soc. (2020) 6:53–64. doi: 10.3126/jpps.v 6i 0.36472 · doi ↗
- 8Matova PM Kamutando CN Magorokosho C Kutywayo D Gutsa F Labuschagne M . Fall-armyworm invasion, control practices and prospects of breeding fall-armyworm- resistant maize in Sub-Saharan Africa. Crop Sci. (2020) 60:2951–70. doi: 10.1002/csc 2.20317, PMID: 33328691 PMC 7702106 · doi ↗ · pubmed ↗