Schistosoma mansoni soluble egg antigen and its key proteins differentially affect dextran sodium sulphate-induced inflammatory bowel disease
Hsiang-Wei Fan, Ho Yin Pekkle Lam

TL;DR
This study explores how Schistosoma mansoni egg antigens, especially Smp40, can reduce inflammation in a mouse model of inflammatory bowel disease.
Contribution
The study identifies Smp40 as a key antigen in Schistosoma mansoni eggs that alleviates IBD symptoms, while other proteins like SM14 worsen the condition.
Findings
Smp40 and SEA reduced DSS-induced IBD in mice.
SM14 exacerbated IBD and caused colonic dysplasia.
GST28 had no significant effect on IBD progression.
Abstract
Inflammatory bowel disease (IBD) is an increasingly prevalent disease, affecting over seven million people worldwide and imposes a heavy burden on public health. The rising prevalence of IBD may be attributed to the hygiene hypothesis, which suggests that reduced exposure to parasites and microbes may weaken the immune system, thereby increasing susceptibility to developing IBD. Studies suggest helminths and their secretory products can modulate the host immunity and attenuate IBD. Our previous research also demonstrated that intestinal schistosomiasis can mitigate chronic IBD symptoms by restoring intestinal immune balance and dysbiosis. While the primary pathology of schistosomiasis results from egg entrapment, we hypothesised that soluble egg antigen (SEA), known for its strong immunomodulatory effect, may contribute to the improvement of IBD. Given that SEA comprises multiple…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
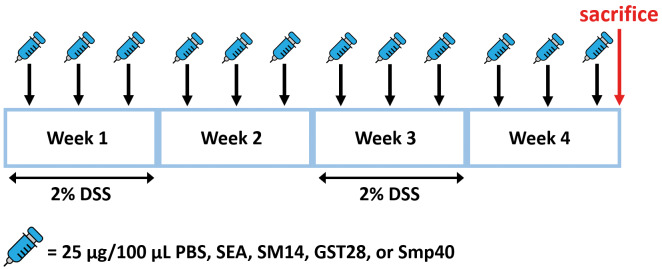 Fig. 1
Fig. 1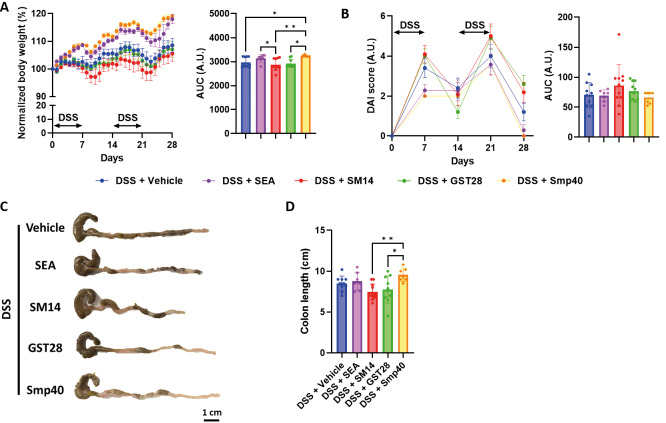 Fig. 2
Fig. 2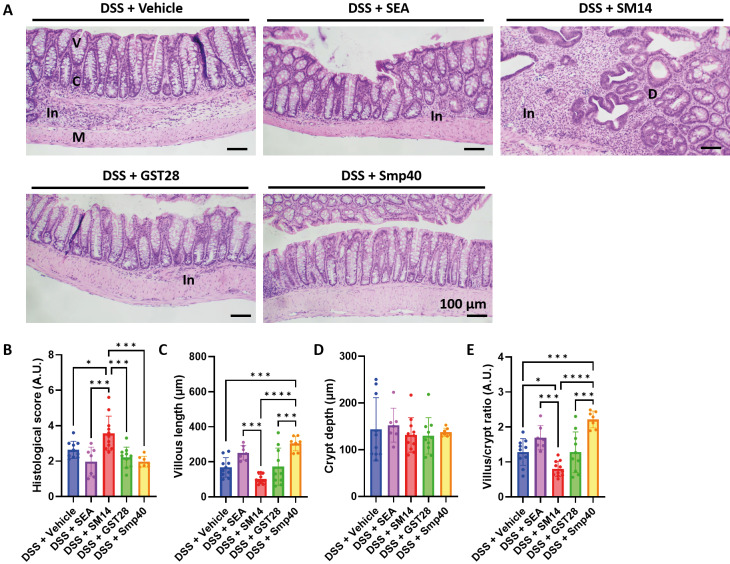 Fig. 3
Fig. 3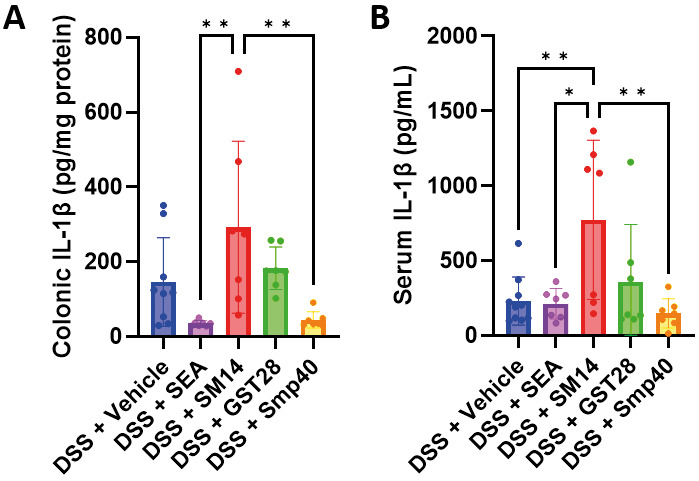 Fig. 4
Fig. 4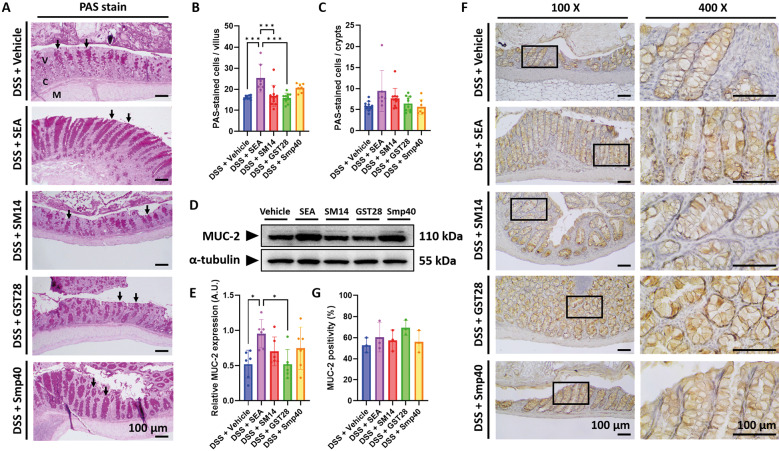 Fig. 5
Fig. 5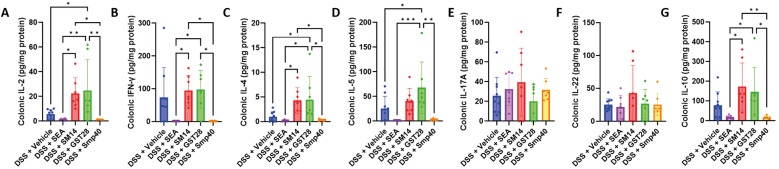 Fig. 6
Fig. 6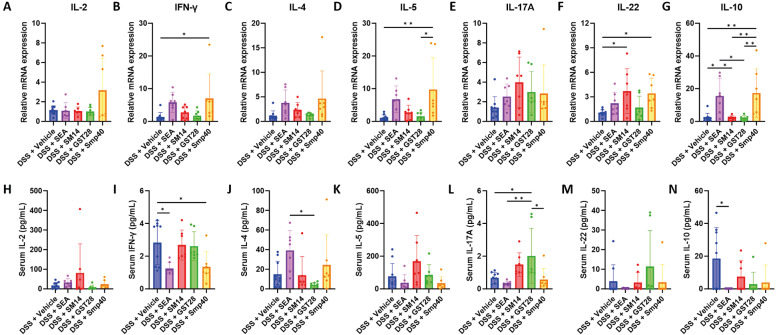 Fig. 7
Fig. 7- —National Science and Technology Council of Taiwan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasites and Host Interactions · Protease and Inhibitor Mechanisms · Liver physiology and pathology
Inflammatory bowel disease (IBD) is an increasingly prevalent chronic inflammatory condition that currently affects over seven million people worldwide.1 The disease places a substantial burden on public health, leading to high medical costs, reduced productivity, and diminished quality of life.1 The etiology of IBD is not clear, but imbalanced immunity, especially the polarisation toward the Th1 response and heightened inflammation, has been suggested as a primary reason leading to chronic intestinal inflammation and IBD.2 In the 21st century, the incidence of IBD has accelerated dramatically in both developing and developed countries. The steep increase in IBD incidence was explained by the hygiene hypothesis, which suggests that people are less likely to be exposed to microorganisms with greater urbanisation and improved hygiene practices. As a result, children may lead to an inadequate maturation of the immune system and loss of its ability to modulate the autoimmune response.3 , 4
It was suggested that several helminths, such as Schistosoma mansoni,5 , 6 Trichuris suis,7 , 8 hookworms,9 and Strongyloides venezuelensis,10 can modulate the immunity and attenuates IBD. Among the diseases caused by these parasites, schistosomiasis ranks as the second most important parasitic disease after malaria, affecting more than 200 million people around the world.11 Despite its prevalence, our previous research suggests that intestinal schistosomiasis can alleviate IBD by reversing intestinal immune imbalance and gut dysbiosis.6 Because the primary pathology of schistosomiasis results from egg entrapment, the observed improvement in colitis may be attributed to proteins secreted by the eggs, which are known to exert strong immunomodulatory effects.12 In natural infection, the trapped eggs continuously secrete soluble egg antigen (SEA), which stimulates the host's T helper 2 (Th2) immune response, characterised by an increase of IL-4, IL-5, and IL-13.13 This Th2 response exerts anti-inflammatory effects that protect the host from excessive inflammation; however, it also contributes to the pathology of schistosomiasis by promoting egg-induced granulomas and fibrosis.13 , 14 When used in isolation, SEA functions as a potent immune modulator and has been explored for its therapeutic potential in autoimmune and inflammatory diseases.12 Although the direct effect of SEA on colitis has yet to be studied, exosomes derived from dendritic cells treated with SEA have been shown to prevent acute colitis by suppressing inflammatory cytokines.15
While the term "SEA" refers to "soluble egg antigen", it does not represent a single, defined antigen. Instead, SEA is a crude extract derived from schistosome eggs, comprising a heterogeneous mixture of biomolecules, including proteins, glycoproteins, polysaccharides, and glycolipids.16 These components originate from various egg-associated sources, such as the eggshell, the miracidium within the egg, and secretory products released by the egg.17 Currently, several SEA-associated antigens have been discovered, including S. mansoni 14-kDa fatty acid-binding protein (SM14), 28-kDa glutathione S-transferase (GST28), and 40-kDa heat shock protein (major egg protein; Smp40). These proteins have previously demonstrated strong immunogenicity,18 , 19 , 20 , 21 with SM14 and GST28 being developed as vaccine candidates and advancing into human clinical trials.22 , 23 GST28 has also been shown to ameliorate experimental colitis by modulating the host immune response.24 , 25 In a phase IIA clinical study, subcutaneous injections of GST28 led to a decrease in the disease activity index in patients with Crohn's disease, with only minimal GST28-related adverse effects observed.26 However, limited by the sample size in that study (only 10 patients enrolled),26 further evidence is needed to clarify the therapeutic effect of GST28 in IBD. Yet, these findings suggest a beneficial role of egg antigens and highlight their potential as a therapeutic agent for IBD.
In this study, we aimed to investigate the effect of crude SEA, along with recombinant SM14, GST28, and Smp40, in a dextran sodium sulphate (DSS)-induced IBD model. Additionally, we sought to clarify the immunomodulatory roles of these proteins in the context of IBD.
MATERIALS AND METHODS
Animals and parasites - Animal experiments were approved by the Institutional Animal Care and Use Committees (IACUC) of Tzu Chi University (No. 113043). Male BALB/c mice were purchased from the National Laboratory Animal Centre (NLAC), NARLabs, Taiwan. All mice were housed under a 23ºC ± 1ºC and a 12-h light/dark cycle condition with 40-60% humidity. Food and water were available ad libitum.
Puerto Rico strain of S. mansoni was provided by the Biomedical Research Institute, MD, USA. The freshwater snail Biomphalaria glabrata was used as an intermediate host and male BALB/c mice were used as the final host. The S. mansoni life cycle was maintained as described previously.6
Preparation of SEA - Mice infected with S. mansoni were sacrificed eight weeks post-infection. Livers were collected and homogenised in ice-cold phosphate-buffered saline (PBS). Eggs were isolated from the liver homogenates by sequential filtration through a series of sieves with decreasing pore size: 420, 177, 105, and 25 μm. Eggs retained on the 25 μm sieve were collected in ice-cold PBS and centrifuged at 370× g for 2 min. The pellet was resuspended in ice-cold PBS, and SEA was prepared by homogenising the eggs with a glass homogeniser. All procedures were conducted under sterile conditions.
Preparation of recombinant SM14, GST28, and Smp40 - Plasmids of the SM14 and GST28 were constructed as previously described.19 , 27 The plasmid of the Smp40 was newly designed and constructed in this experiment. Briefly, a polymerase chain reaction (PCR) was performed on the cDNA of S. mansoni worm with the primer: Forward 5' GAGACATATGTCTGGTGGGAAACAACATAAC 3' and Reverse 5' GATACTCGAGGTGAGTAATTGCATGTTGCTTC 3'. The PCR product was run on a 1% agarose gel and purified by EasyPure PCR/Gel extraction Kit (Bioman Scientific, Taipei, Taiwan). The resulting product was digested with NdeI and XhoI and ligated into plasmid pET-28a [Supplementary data (Figure A)].
All plasmids were transformed into the Escherichia coli BL21 (DE3) for protein expression. Transformed BL21 (DE3) was plated onto LB agar containing 50 μg/mL kanamycin and incubated overnight. Single colonies were inoculated into LB broth and cultured at 37ºC with shaking until an OD_600_ of 0.8 was reached. Protein expression was induced by adding 1 mM isopropyl-beta-D-thiogalactopyranoside (IPTG), followed by incubation at 37ºC with shaking for 4 h. Cultures were harvested by centrifugation at 10,000 × g for 20 min at 4ºC. The supernatant was discarded, and the remaining pellets were resuspended in lysis buffer (50 mM NaH_2_PO_4_, 300 mM NaCl, 10 mM imidazole, pH 8). The cells were lysed using a MiniBead-beater and the lysate was collected and centrifuged at 20,000 × g for 15 min at 4ºC. The supernatant containing soluble proteins was collected for purification by metal affinity chromatography (Cat#: 7880011; Bio-Rad, CA, USA). The eluted proteins were analysed on a 13.5% sodium dodecyl sulphate-polyacrylamide gel (SDS-PAGE) and stained with 0.1% Coomassie blue. An SDS-PAGE gel showing the bacterial lysates for Smp40 with and without IPTG induction, as well as the eluted Smp40 protein, is presented in Supplementary data (Figure B-D).
Animal treatment - Mice were six weeks of age at the beginning of the experiments. Mice were divided randomly into five groups. The sample size was calculated using the resource equation, resulting in the allocation of three to five mice per group. The experiment was independently repeated twice. All the mice were given 2% DSS (Cat#: J63606.22, Thermo Fisher Scientific, MA, USA) in their drinking water at week 1 and week 3, whereas normal drinking water was provided at week 2 and week 3. Different egg antigen proteins (at a concentration of 250 μg/mL) were peritoneally injected into the mice three times a week on alternate days during the experimental duration. All the mice were sacrificed after four weeks (Fig. 1). At the time of sacrifice, blood was collected through cardiac puncture, and organs were collected for subsequent experiments.
Disease activity index (DAI) - The DAI of the mice was assessed weekly based on three criteria: percentage of weight loss, presence of rectal bleeding, and stool consistency. Each parameter was scored on a scale from 0 to 4 [Supplementary data (Table I)], and the DAI score was calculated as the sum of the three individual scores.
Tissue processing, staining, and histopathology - Tissues were fixed with 10% formalin, embedded in paraffin, and sectioned into thin slices for haematoxylin & eosin (H&E) and periodic acid-Schiff (PAS) staining as previously described.6 Colonic sections stained with H&E were scored for epithelial damage, lamina propria inflammation, muscularis propria thickening, and fibrosis. Each criterion was assigned a score of 0, not observed; 1, mild; 2, moderate; 3, intensive. Measurements were also done on villous height, crypt depth, and villi-to-crypt ratio. Colonic sections stained with PAS were counted for the number of positive cells. At least ten random fields were examined and scored in each section.
Colonic sections were also stained for MUC-2 by immunohistochemistry staining. Briefly, antigens were retrieved from the sections by incubating them in boiling EDTA buffer for 20 min. Subsequently, the sections were treated with 3% H_2_O_2_ for 10 min and incubated overnight at 4ºC with MUC-2 (1:300; Cat#: A14659; ABclonal). The sections were then incubated with HRP-conjugated anti-rabbit secondary antibody (1:1000; Cat#: C04003; Croyez Biosciences) for 30 min and 3, 30-diaminobenzidine (DAB; Cat#: 34000; Thermo Fisher Scientific) for 12 min. Sections were counterstained with haematoxylin and rehydrated with increasing ethanol concentration.
RNA extraction, cDNA synthesis, and quantitative PCR (qPCR) - Total RNA was extracted by homogenising tissues in TRIzol reagent (Invitrogen; Thermo Fisher Scientific) and purified using the standard chloroform extraction method. Five micrograms of total RNA were used to generate cDNA using a GScript First-Strand Synthesis Kit (GeneDireX, Taiwan). The qPCR reaction was performed by 2× qPCRBIO SyGreen Blue Mix Lo-ROX (PCR Biosystems, London, UK) using the Roche LightCycler 480 system. Amplification and detection were performed as follows: 55 cycles of denaturation at 95ºC for 10 s, 58 or 60ºC for 15 s, and extension at 72ºC for 25 s. The oligonucleotide primers used are shown in Supplementary data (Table II). Relative gene expression was calculated using the 2^-ΔΔCT^ method with β-actin as the housekeeping gene.
Measurement of cytokine levels in serum and colonic tissues - Serum was separated by centrifuging whole blood at 1,500 × g for 15 min. The colon was homogenised in PBS and centrifuged at 12,000 × g for 15 min at 4ºC. Levels of IL-1β (Cat#: 432604; BioLegend, San Diego, CA, USA), IL-2 (Cat#: 431001; BioLegend), IFN-γ (Cat#: 430801; BioLegend), IL-4 (Cat#: 431101; BioLegend), IL-5 (Cat#: 431204; BioLegend), IL-17A (Cat#: 88-7371; Thermo Fisher Scientific), IL-22 (Cat#: 88-7422; Thermo Fisher Scientific), and IL-10 (Cat#: 431411; BioLegend) in the sera or colonic homogenate were measured using a standard sandwich enzyme-linked immunosorbent assay (ELISA) kit. Protein concentrations of colonic homogenates were determined by the Bradford method using a Bio-Rad Protein Assay Dye (Bio-Rad Laboratories, Hercules, CA, USA).
Western blot - Total proteins were extracted, separated by 10% SDS-PAGE, and transferred onto PVDF membranes (EMD Millipore, Burlington, MA, USA). After blocking with 5% non-fat milk, the membranes were incubated with MUC-2 (1:1000; Cat#: A14659; Abclonal) or α-tubulin (1:5000; Cat#: AC012; Abclonal) at 4ºC overnight. Membranes were then incubated with HRP-conjugated anti-rabbit (1:5000; Cat#: C04003; Croyez Biosciences) or HRP-conjugated anti-mouse (1:5000; Cat#: C04001; Croyez Biosciences) secondary antibody for 1 h. Membranes were developed using ECL detection reagent (EMD Millipore). Relative protein levels were quantified using Image J (Version 1.46, National Institute of Health, Bethesda, MD, USA), and protein densitometry was expressed relative to that of α-tubulin.
Statistical analysis - Two independent experimental replicates were performed. Data from the same groups across both repeats were pooled, and a statistical comparison was performed on the combined dataset without applying interim statistical adjustments. Data are presented as mean ± standard deviation (SD) unless otherwise specified. Statistical significance between groups was evaluated using one-way analysis of variance (ANOVA) followed by Tukey's honest significant difference test. Differences were considered statistically significant at p < 0.05 (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001). All analyses were performed by GraphPad Prism software version 9.4.1.
RESULTS
Soluble egg antigen and Smp40 improve DSS-induced colitis, whereas SM14 worsens DSS-induced colitis - Mice were treated with two repeated cycles of 2% DSS to develop colitis. At the same time, mice were intraperitoneally injected with either crude SEA or recombinant egg antigens, SM14, GST28, and Smp40, at a concentration of 250 μg/mL three times a week (Fig. 1). Injection of different egg antigens had minimal but different effects on the body weight of the mice. SEA- and Smp40-treated mice have improved body weight compared to the vehicle-treated mice, but only Smp40-treated mice reached statistical significance (Fig. 2A). Regarding the severity of colitis, mice treated with SM14 showed a slightly higher, although not statistically significant, DAI score (Fig. 2B) and shortened colon length (Fig. 2C-D). Histopathological analysis of the colon suggested inflammatory infiltration in all the groups, though it was notably reduced in the SEA-, GST-, and Smp40-treated mice. In contrast, SM14-treated mice showed increased inflammatory infiltrations and were the only group in which colonic dysplasia was observed (Fig. 3A). Histological analysis also revealed a worsened histological score in the SM14-treated group (Fig. 3B). Further analysis suggested that Smp40-treated mice exhibited increased villus length, while SM14-treated mice had a shorter villus length (Fig. 3C). While these antigens did not affect the depth of the crypt (Fig. 3D), Smp40-treated mice had a significantly higher villus-to-crypt ratio (Fig. 3E), suggesting improved intestinal architecture and functions. SM14-treated mice, on the other hand, exhibited a reduced villus-to-crypt ratio compared to other groups (Fig. 3E), indicating colonic damage and compromised intestinal function.
experimental scheme. Mice were given 2% dextran sulphate sodium (DSS) in their drinking water at weeks 1 and 3, and normal drinking water was provided at weeks 2 and 4. Different egg antigens (at a concentration of 250 μg/mL) were peritoneally injected into the mice three times a week on alternate days during the experimental duration. All the mice were sacrificed after four weeks.
*schistosome egg antigen differently alters dextran sulphate sodium (DSS)-induced inflammatory bowel disease in mice. (A) Body weight change of the mice and the corresponding area under curve (AUC) chart. (B) Disease activity index and the corresponding AUC chart. (C) Representative colon images of the mice. (D) Colon length. n = 7-11 mice. Data are presented as mean ± standard deviation (SD). *p < 0.05 and *p < 0.01. Significance determined by one-way analysis of variance (ANOVA).
*schistosome egg antigen differently alters colon histopathology in dextran sulphate sodium (DSS)-induced colitis in mice. (A) Representative histological images of the colon tissue. V: villus; C: crypts; M: muscularis mucosae; In: inflammation; D: dysplasia. (B) Histological score of the colon. (C) Measurement of villi length, (D) crypt depth, and (E) villi-to-crypt ratio. n = 7-11 mice. Data are presented as mean ± standard deviation (SD). *p < 0.05; ***p < 0.001; and ***p < 0.0001. Significance determined by one-way analysis of variance (ANOVA).
To further assess inflammation, colonic IL-1β levels were measured. SEA- and Smp40-treated groups exhibited decreased IL-1β levels, while the SM14-treated group showed increased levels (Fig. 4A). Similarly, the SM14-treated group showed a higher serum IL-1β level compared to the other groups (Fig. 4B). Collectively, these results suggest that different schistosome egg antigens may exert differential effects on DSS-induced IBD. While SEA and Smp40 appear to alleviate colitis, SM14 worsens colitis.
*SM14 exacerbates colonic inflammation in dextran sulphate sodium (DSS)-induced mice. (A) Colonic IL-1β levels. (B) Serum IL-1β levels. n = 7-11 mice. Data are presented as mean ± standard deviation (SD). *p < 0.05 and *p < 0.01. Significance determined by one-way analysis of variance (ANOVA).
SEA improves intestinal barrier by increasing goblet cells and upregulating MUC-2 expression - To further investigate the function of the intestinal barrier, PAS staining was used to identify mucin within goblet cells, which appeared dark pink in contrast to the pale pink background. The result suggested that SEA-treated mice had significantly higher colonic goblet cells in the villus compared to the vehicle and other treatment groups (Fig. 5A-B). Yet, goblet cell numbers were not significantly different in the crypt (Fig. 5C). Western blot analysis revealed increased MUC-2 expression in the SEA-treated group (Fig. 5D-E). However, immunohistochemistry staining for MUC-2 in colonic tissue revealed only minimal and non-significant changes (Fig. 5F-G). While SEA treatment showed a substantial effect on the intestinal barrier, changes in other treatment groups were not as evident as those in SEA-treated groups.
*schistosome egg antigen alters intestinal barrier in mice with dextran sulphate sodium (DSS)-induced colitis. (A) Representative periodic acid-Schiff (PAS)-stained colon section. V: villus; C: crypts; M: muscularis mucosae. The black arrows indicate goblet cells containing mucin (PAS-stained cells), which appear as darker pink-coloured cells. (B-C) Quantification of PAS-stained cells per (B) villus and (C) crypt. (D) Representative western blot image of MUC-2 expression. (E) Relative protein expression of MUC-2. Densitometric values were normalised to α-tubulin. (F) Representative immunohistochemistry staining of MUC-2. (G) Percentage of positive expression of MUC-2. For (A-C), n = 7-11 mice; for (D-E), n = 6 mice; for (F-G), n = 3 mice. Data are presented as mean ± standard deviation (SD). *p < 0.05 and **p < 0.001. Significance determined by one-way analysis of variance (ANOVA).
Schistosome egg antigen differently modulates immune responses of colitic mice - Given the critical role of immune imbalance in the pathogenesis of IBD2 and the immunoregulatory effects of schistosome eggs and their antigens,12 we next aimed to investigate the immune profile in the mice. Measuring the cytokines in the colon revealed significant suppression of IL-2, IFN-γ, IL-4, IL-5, and IL-10 levels in the SEA- and Smp40-treated groups compared to SM14- and GST28-treated groups. Although a noticeable decrease in the cytokine levels was also seen in the SEA- and Smp40-treated group compared to the vehicle-treated group, the results were statistical insignificant (Fig. 6A-D, G). In contrast, SM14-treated group showed a significant increase in IL-2, IFN-γ, IL-4, and IL-10, but only when compared to the SEA- and Smp40-treated groups (Fig. 6A-C, G). Additionally, GST28-treated group exhibited a significant increase in the colonic IL-2, IL-4, and IL-5 levels compared to the vehicle-treated group (Fig. 6A, C-D). It is worth noting that IL-17A and IL-22 levels were not altered by any of the egg antigens (Fig. 6E-F).
*schistosome egg antigen differently modulates colonic immune responses in dextran sulphate sodium (DSS)-induced mice. Colonic levels of (A) IL-2, (B) IFN-γ, (C) IL-4, (D) IL-5, (E) IL-17A, (F) IL-22, and (G) IL-10. Cytokine levels were relative to the total protein levels. n = 7-10 mice. Data are presented as mean ± standard deviation (SD). *p < 0.05; **p < 0.01; and **p < 0.001. Significance determined by one-way analysis of variance (ANOVA).
The immune profile in the spleen and serum was also analysed to assess systemic immunity. As noted, the Smp40-treated group showed a significant upregulation of splenic expression of IFN-γ, IL-5, IL-22, and IL-10 (Fig. 7B, D, F-G). IL-2, although not showing statistical significance (p = 0.0597), also revealed a similar increase in the Smp40-treated group (Fig. 7A). The SM14-treated group also exhibited increased IL-22 expression compared to the vehicle-treated group (Fig. 7F), while the SEA-treated group significantly upregulated splenic IL-10 expression (Fig. 7G). On the other hand, SEA-treated group significantly suppressed serum IFN-γ and IL-10 levels while Smp40-treated group suppressed serum IFN-γ levels (Fig. 7I, N). In addition, the GST28-treated group exhibited an increase in serum IL-17A levels (Fig. 5L). These results suggest that distinct Schistosoma egg antigens can differentially modulate both local intestinal immune response and systemic immunity.
*schistosome egg antigen differently modulates splenic and serum immune responses in dextran sulphate sodium (DSS)-induced mice. Splenic mRNA expression of (A) IL-2, (B) IFN-γ, (C) IL-4, (D) IL-5, (E) IL-17A, (F) IL-22, and (G) IL-10. Serum levels of (H) IL-2, (I) IFN-γ, (J) IL-4, (K) IL-5, (L) IL-17A, (M) IL-22, and (N) IL-10. n = 7-10 mice. Data are presented as mean ± standard deviation (SD). *p < 0.05 and *p < 0.01. Significance determined by one-way analysis of variance (ANOVA).
DISCUSSION
Soluble egg antigen of Schistosoma has demonstrated good immunogenic properties and has been investigated as a potential therapy for various autoimmune diseases.12 Previous studies have shown that natural Schistosoma infection or exposure to SEA can alleviate colitis by modulating the host immune response.6 , 28 , 29 , 30 However, the specific active components within the SEA responsible for this effect remained unknown, considering the fact that SEA is a mixture of many different proteins.16 In this study, we identified that both the crude SEA extract and recombinant Smp40 alleviated DSS-induced colitis, whereas recombinant SM14 exacerbated the disease. At the same time, recombinant GST28 had no significant effect on colitis (Figs 2-4). Further investigation revealed that the observed differences in the IBD outcomes may be due to the modulation of immune response by different antigens, albeit in distinct ways (Figs 5-7).
Both naturally-occurring IBD and experimentally-induced colitis are driven by an altered and imbalanced immune response, typically characterised by an exaggerated Th1 response.31 Therefore, suppressing this skewed immunity has been considered a potential therapeutic strategy. Treatment of colitic mice with SEA and Smp40 resulted in a significant reduction of colonic Th1 cytokines, IL-2 and IFN-γ (Fig. 6A-B), which was accompanied by an improvement of the colitis (Figs 2-3). This result is consistent with previous studies showing that SEA can suppress Th1 cytokines, leading to the amelioration of autoimmune diseases.32 However, Th2 cytokines, IL-4 and IL-5, and Treg cytokine IL-10 were also suppressed in the colons of mice treated with SEA and Smp40 (Fig. 6C-D, G). Although SEA is generally known to induce a Th2 response,33 it has also been suggested that SEA might lead to a substantial reduction of Th2 response under certain inflammatory settings, including colitis.30 This effect could possibly be due to a pre-existing Th1-dominant immune milieu that constrains SEA from driving an excessive Th2 immune response, thereby restoring Th1 and Th2 balance rather than amplifying Th2 immune responses. The effect of IL-10, a multi-functional cytokine, in IBD is controversial. Although IL-10 has been shown to have a protective effect in IBD,33 elevated levels were also found in IBD patients and could contribute to disease risk.34 , 35 Previous studies have suggested that S. mansoni infection can suppress IL-10 expression, leading to protection from colitis.5 Similarly, the suppression of IL-10 by SEA and Smp40 observed in this study may contribute to a similar protection. In addition, SEA has been suggested to modulate the gut microbiome and intestinal metabolism in colitic mice, suggesting that the gut microbiome may play a role in SEA-mediated inhibition of inflammation in IBD.36
On the other hand, Smp40 is an immunomodulatory protein homologous to heat shock protein (HSP),21 and several HSPs have already been identified in Schistosoma species.37 Although there is no direct evidence that schistosome-related HSPs influence IBD, various studies have shown that HSPs, such as HSP70, may help ameliorate the disease.38 Future studies focusing on the protein and molecular characterisation of the Smp40 may provide further knowledge into its role in IBD.
In colitic mice treated with SM14, increased colonic levels of IL-2, IL-4, and IL-10 were observed (Fig. 6A, C, G). Notably, SM14 led to a more pronounced inflammation and the development of colonic dysplasia (Fig. 3A), a known risk factor for colorectal cancer.39 These findings therefore underscore the potential adverse effects of SM14. We hypothesise that SM14 promotes significant inflammatory cell infiltration and elevated colonic IL-2 levels, which together create a pro-inflammatory environment that may contribute to the development of dysplasia. The increase of IL-4, on the other hand, is crucial for the development of colorectal cancer.40 , 41 It has been shown to induce epithelial-mesenchymal transition and promote the aggressiveness of colorectal cancer cells.42 , 43 Although IL-10 is generally known for its potential to suppress inflammation and control tumour-promoting inflammation,44 it has also been associated with colorectal cancer development.45 In fact, deficiency of IL-10 has been reported to enhance the efficacy of dendritic cell-based immunotherapy,45 and increased serum IL-10 levels in colorectal cancer patients have been associated with a higher recurrence rate and poorer prognosis.34 Furthermore, SM14 is a fatty acid-binding protein.12 While intestinal fatty acid-binding protein that primarily expressed on intestinal epithelial cells plays a pivotal role in intestinal inflammation,46 exogenous injection of SM14, although differing in their host sources, may also elicit a similar adverse effect. Collectively, these results suggest that SM14 may exacerbate colitis and potentially promote progression toward colorectal cancer. Although SM14 has been used as a vaccine candidate against schistosomiasis and has progressed through phase I and phase II clinical trials with a safe and strong immunogenic profile,22 , 47 caution should be exercised in the future when considering its use in IBD patients.
We also observed that GST28-treated mice had increased colonic levels of IL-2, IL-4, and IL-5 (Fig. 6A, C-D); however, no changes in colitis severity were seen (Fig. 2). The therapeutic potential of GST28 in IBD has already been investigated in several experimental and clinical studies.24 , 25 , 26 It has been suggested that GST28 enhances colonic Th2 response, recruits eosinophils, and suppresses Th1 response in order to alleviate colitic symptoms.24 , 25 Our findings of increased IL-4 and IL-5 are consistence with these previously reported effects.24 Previously, GST28 has been shown to increase IL-2 expression in S. mansoni infection.27 Given that IL-2 is a cytokine that is important for the expansion and function of Treg cells,48 its elevation may provide immunoregulatory benefits. In addition, low-dose IL-2 treatment has been shown to reduce disease severity in 2,4-dinitrobenzene sulfonic acid (DNBS)-induced colitic mice49 and in patients with moderate to severe ulcerative colitis.50 Despite the changes in the immune response, GST28 treatment did not improve IBD outcomes in our study. One possible reason is the absence of an adjuvant, which may be necessary to fully activate the anti-inflammatory effect of GST28.24 , 51 Additionally, dosing of GST28 appears to be an important factor. In a previous study, the use of GST28 at doses of 5 and 50 μg/kg significantly suppressed colitis, whereas a higher dose of 500 μg/kg had no effect on colitis.25 In our current study, the dosage used (250 μg/mL, approximately equal to 830 μg/kg) may have exceeded the effective therapeutic range. However, our previous work showed that GST28 at this same dose, when combined with heat-killed Cutibacterium acnes as an adjuvant, induced significant immune modulation and reduced disease severity of schistosomiasis,27 emphasising the importance of adjuvant inclusion. Therefore, future studies may involve the combination use of GST28 with appropriate adjuvants in the context of IBD. The use of adjuvants may also be applied and investigated for other egg antigens to enhance their immune modulatory potential.
Th17 responses are also involved in the pathogenesis of IBD by promoting IL-17-mediated intestinal inflammation.52 However, our study did not reveal any significant changes in the intestinal Th17 response following treatment with any of the tested egg antigens (Fig. 6E-F). Previous studies have suggested that SEA can protect against skin transplant rejection by modulating Th1 and Th2 responses, without affecting IL-17^+^CD4^+^ T cells.53 Similarly, SM14 has been shown to drive the development of IL-10-producing T cells, but not IL-17-producing T cells, in a C57BL/6 mouse model.18 Moreover, the choice of mouse strain may also influence the immune outcome. A prior study comparing Giardia infection in BALB/c and C57BL/6 mice showed a higher parasitic burden in BALB/c mice, which is associated with a lower Th17 activity.54 Another report suggested that Mycoplasma pneumoniae infection in BALB/c mice exhibited a lower Th17 response compared to DBA/2 mice.55 Therefore, while Th17 responses play a role in IBD, the tested egg antigens may not appear to significantly influence colonic Th17 activity. Additionally, variations in immune responses among different mouse strains highlight the importance of host genetic background in interpreting antigen-specific immunological outcomes in IBD.56 , 57 Therefore, future studies comparing different mouse models may be warranted.
Finally, we investigated the systemic immune response in the mice, which the results differed from the local response as observed in the colon. Smp40-treated mice showed a significant increase in splenic IL-2, IFN-γ, IL-5, IL-22, and IL-10 levels, whereas SEA-treated mice had higher splenic IL-10 levels, and SM14-treated mice had higher splenic IL-22 levels (Fig. 7A-G). A previous study has shown that Smp40 can elicit a Th1 response, characterised by increased secretion of IL-2 and IFN-γ from splenic lymphocytes, even as the overall immune response shifts toward Th2 dominance.58 Additionally, Smp40 has been shown to increase IL-10 expression in peripheral blood mononuclear cells from S. mansoni-infected patients.21 SM14 induction of splenic IL-22 may contribute to the excess inflammation seen in the colon (Fig. 3A), as IL-22 is known to recruit neutrophils and promote an inflammatory environment in ulcerative colitis.59 SEA has been shown to stimulate IL-10 production in splenic B cells, which aligns with our results.18 From the same study, it was suggested that glycosylated molecules in high molecular weight fractions of SEA are responsible for this effect,18 suggesting a specific component may underlie its immunomodulatory effect.
In this study, different egg antigens demonstrated different immunomodulatory effects in the context of IBD, leading to varying disease outcomes. While many studies have highlighted the promising therapeutic potential of SEA in treating IBD, it is important to recognise that SEA is a complex mixture of many different molecules, each of which may exert both beneficial and detrimental effects. Therefore, a deeper understanding of the safety and mechanisms of these individual antigens is necessary before considering their use in clinical settings.
SUPPLEMENTARY MATERIALS
Supplementary material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Solitano V Bernstein CN Dotan I Dignass A Domilici R Dubinsky MC Shaping the future of inflammatory bowel disease: a global research agenda for better management and public health response Nat Rev Gastroenterol Hepatol 202522438524026343310.1038/s 41575-025-01063-x · doi ↗ · pubmed ↗
- 2Hu C Liao S Lv L Li C Mei Z Intestinal immune imbalance is an alarm in the development of IBD Mediators Inflamm 2023202310739843755455210.1155/2023/1073984 PMC 10406561 · doi ↗ · pubmed ↗
- 3Koloski NA Bret L Radford-Smith G Hygiene hypothesis in inflammatory bowel disease: a critical review of the literature World J Gastroenterol 200814165731818654910.3748/wjg.14.165PMC 2675108 · doi ↗ · pubmed ↗
- 4Weinstock JV Elliott DE Helminths and the IBD hygiene hypothesis Inflamm Bowel Dis 200915128331868019810.1002/ibd.20633 · doi ↗ · pubmed ↗
- 5Ruyssers NE De Winter BY De Man JG Loukas A Pearson MS Weinstock JV Therapeutic potential of helminth soluble proteins in TNBS-induced colitis in mice Inflamm Bowel Dis 2009154915001902390010.1002/ibd.20787 · doi ↗ · pubmed ↗
- 6Lin YR Lam LY Chang CM Lam HYP Concomitant occurrence of chronic Schistosoma mansoni infection and chronic colitis restore immune imbalance and dysbiosis leading to protection against intestinal colitis and schistosome egg-induced intestinal fibrosis Mem Inst Oswaldo Cruz 2025120 e 2400454033218710.1590/0074-02760240045 PMC 12051921 · doi ↗ · pubmed ↗
- 7Summers RW Elliott DE Qadir K Urban Jr JF Thompson R Weinstock JV Trichuris suis seems to be safe and possibly effective in the treatment of inflammatory bowel disease Am J Gastroenterol 2003982034411449978410.1111/j.1572-0241.2003.07660.x · doi ↗ · pubmed ↗
- 8Sandborn WJ Elliott DE Weinstock J Summers RW Landry-Wheeler A Silver N Randomised clinical trial: the safety and tolerability of Trichuris suis ova in patients with Crohn's disease Aliment Pharmacol Ther 201338255632373095610.1111/apt.12366 · doi ↗ · pubmed ↗