Mitochondrial genomes and phylogeny of Atratus and Educator Group species of the Melanoconion Section of Culex (Melanoconion) (Diptera: Culicidae)
Tatiane Marques Porangaba de Oliveira, Peter Gordon Foster, Ivy Luizi Rodrigues de Sá, Maria Anice Mureb Sallum

TL;DR
This study sequenced mitochondrial genomes of Culex mosquitoes to better understand their genetic diversity and evolutionary relationships.
Contribution
The study provides new mitochondrial genome data for multiple Culex species in the Melanoconion subgenus and identifies phylogenetic patterns.
Findings
Partial mitochondrial genomes included 12 protein coding genes, 15 tRNA, and 1 rRNA.
Phylogenetic analysis suggested the presence of three putative species.
ND5 gene showed a shorter length due to a stop codon in the new sequences.
Abstract
Culex (Melanoconion) species are known to act as vectors for different arboviruses, and little is known about the mitochondrial genome of these species. Aiming to expand the genetic knowledge of this subgenus, a 12Kb fragment of the mitochondrial genome was sequenced from 23 specimens belonging to the Atratus and Educator Groups of the subgenus Melanoconion of Culex. The sequenced specimens were morphologically identified as Culex dunni, Culex ensiformis, Culex theobaldi, Culex trigeminatus, Culex eknomios, Culex zeteki, Culex near commevynenis, Culex angularis, Culex longistriatus, and Culex near vaxus. The reads were assembled with the reference genome of Culex quinquefasciatus and MITOS2 was used for gene annotation. Values of guanine-cytosine (GC) and adenine-thymine (AT) skews, nucleotide diversity, ratio between non-synonymous (Ka) and synonymous (Ks) substitution, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
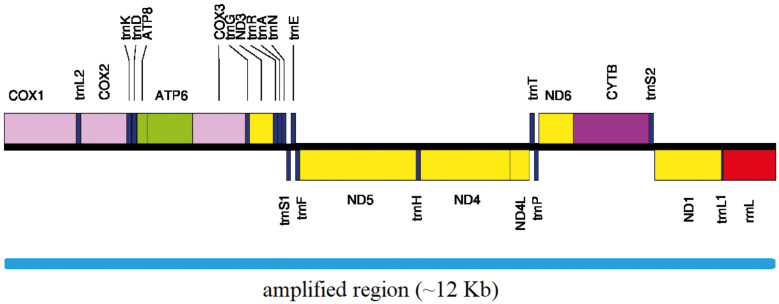 Fig. 1
Fig. 1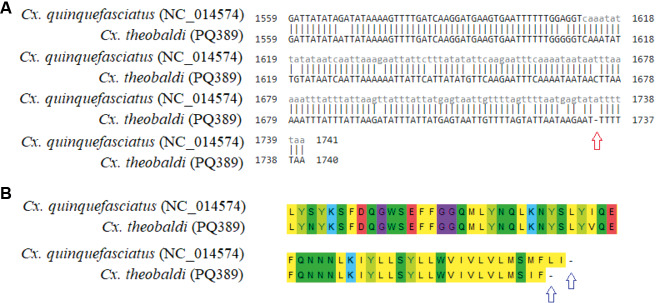 Fig. 2
Fig. 2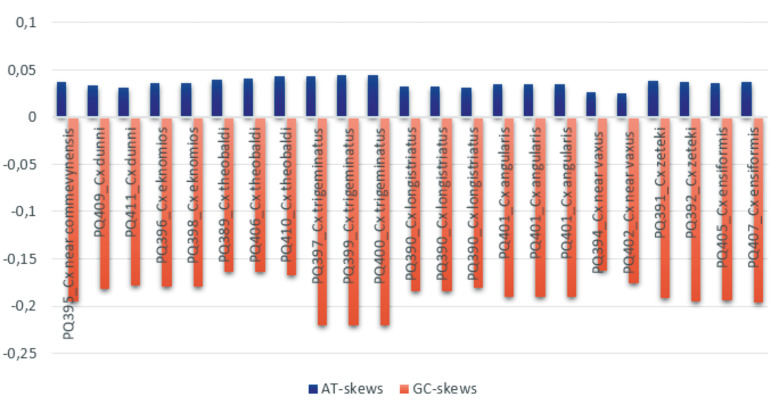 Fig. 3
Fig. 3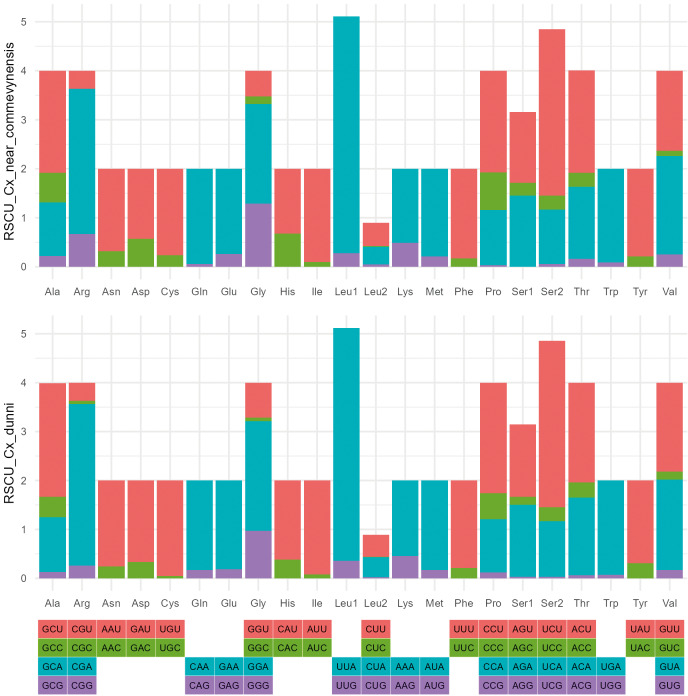 Fig. 4
Fig. 4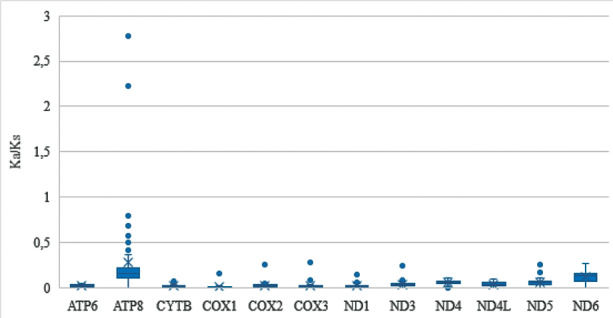 Fig. 5
Fig. 5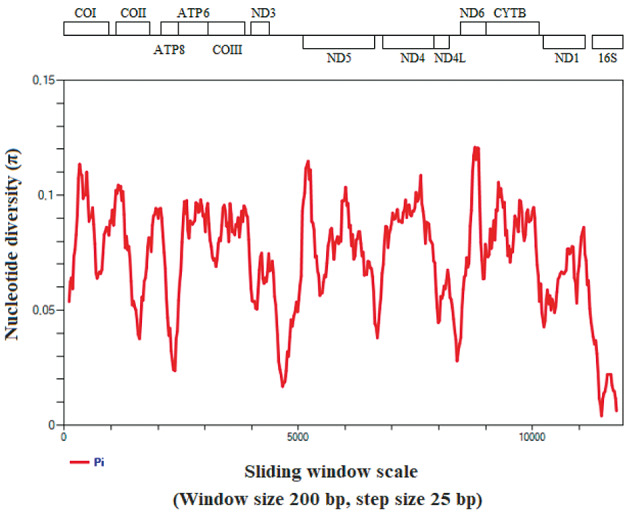 Fig. 6
Fig. 6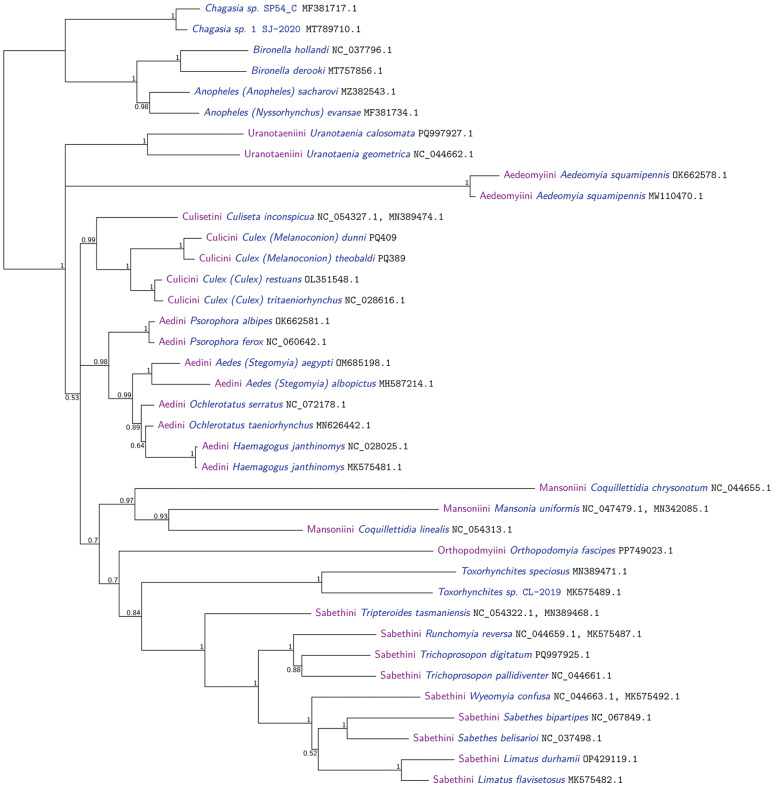 Fig. 7
Fig. 7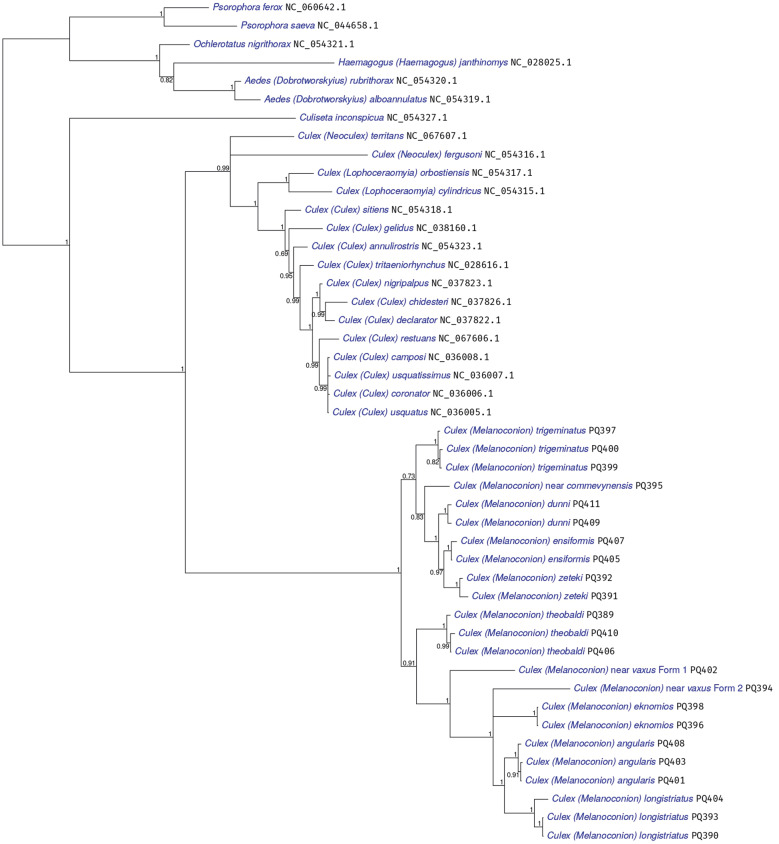 Fig. 8
Fig. 8| ID_sequences | Species | T% | C% | A% | G% | AT-skews | GC-skews |
| PQ395 |
| 36 | 15 | 38,8 | 10,1 | 0,037433 | -0,195219 |
| PQ409 |
| 36,4 | 14,6 | 38,9 | 10,1 | 0,033201 | -0,182186 |
| PQ411 |
| 36,5 | 14,5 | 38,8 | 10,1 | 0,030544 | -0,178862 |
| PQ396 |
| 36,9 | 13,8 | 39,6 | 9,6 | 0,035294 | -0,179487 |
| PQ398 |
| 36,9 | 13,8 | 39,6 | 9,6 | 0,035294 | -0,179487 |
| PQ405 |
| 36 | 15,1 | 38,7 | 10,2 | 0,036145 | -0,193676 |
| PQ407 |
| 35,9 | 15,2 | 38,7 | 10,2 | 0,037534 | -0,196850 |
| PQ389 |
| 36,3 | 14,2 | 39,3 | 10,2 | 0,039683 | -0,163934 |
| PQ406 |
| 36,3 | 14,2 | 39,4 | 10,2 | 0,040951 | -0,163934 |
| PQ410 |
| 36,1 | 14,3 | 39,3 | 10,2 | 0,042440 | -0,167347 |
| PQ397 |
| 35,7 | 15,5 | 38,9 | 9,9 | 0,042895 | -0,220472 |
| PQ399 |
| 35,6 | 15,5 | 38,9 | 9,9 | 0,044295 | -0,220472 |
| PQ400 |
| 35,6 | 15,5 | 38,9 | 9,9 | 0,044295 | -0,220472 |
| PQ390 |
| 37,1 | 13,8 | 39,6 | 9,5 | 0,032595 | -0,184549 |
| PQ393 |
| 37,1 | 13,8 | 39,6 | 9,5 | 0,032595 | -0,184549 |
| PQ404 |
| 37,2 | 13,7 | 39,6 | 9,5 | 0,031250 | -0,181034 |
| PQ401 |
| 36,8 | 14,1 | 39,4 | 9,6 | 0,034121 | -0,189873 |
| PQ403 |
| 36,8 | 14,1 | 39,4 | 9,6 | 0,034121 | -0,189873 |
| PQ408 |
| 36,8 | 14,1 | 39,4 | 9,6 | 0,034121 | -0,189873 |
| PQ394 |
| 37 | 13,9 | 39 | 10 | 0,026316 | -0,163180 |
| PQ402 |
| 37,4 | 13,7 | 39,3 | 9,6 | 0,024772 | -0,175966 |
| PQ391 |
| 36 | 14,9 | 38,9 | 10,1 | 0,038718 | -0,192000 |
| PQ392 |
| 36 | 15 | 38,8 | 10,1 | 0,037433 | -0,195219 |
- —FAPESP, CNPq
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Genomics and Phylogenetic Studies · Neurobiology and Insect Physiology Research
The majority of species of the subgenus Melanoconion of Culex occurs in neotropical regions. Some of the species from this subgenus are recognized for acting as important vectors of several arboviruses affecting humans and other vertebrates, such as Venezuelan Equine Encephalitis complex, West Nile Virus, and Eastern Equine Encephalitis Virus.1 2 3 4 5 6 Recently, Agua Salud alphavirus was isolated from Culex (Melanoconion) mosquitoes collected from the Brazilian Amazon (Belém municipality, Pará State),7 suggesting that they are natural hosts of these viruses.
Torres-Gutierrez and Sallum8 updated the catalogue of the subgenus Melanoconion published in 1992 by Pecor et al.9 In the new catalogue, this subgenus comprises two sections, 21 groups, 23 subgroups, and 160 valid species. Recently, revisions were carried out on the Atratus and Educator Groups of the Melanoconion Section of the subgenus Melanoconion resulting in the description of eight new species and removal of five species from synonymy.10 , 11 Thus, after the revisions this subgenus encompasses 173 valid species8 , 10 , 11 which are present in the Melanoconion and Spissipes Sections.8 , 10 , 11 , 12 , 13 The Melanoconion Section includes 150 species divided in 13 groups.8 , 10 , 11 The Atratus Group encompass the following species: Culex atratus; Culex caribeanus; Culex commevynensis; Culex columnaris; Culex comptus; Culex dunni; Culex ensiformis; Culex exedrus; Culex longisetosus; Culex longistylus; Culex loturus; Culex spinifer; Cx. trigeminatus; and Culex zeteki.10 The Educator Group includes the species Culex aneles; Culex angularis; Culex apeteticus; Culex aphyllus; Culex bibulus; Culex cristovaoi; Culex educator; Culex eknomios; Culex inadmirabilis; Culex longistriatus; Culex rachoui; Culex spiniformis; Culex theobaldi; and Culex vaxus.11
Taxonomic identification of species of this subgenus by female characters is hampered by the great similarity between these characteristics and, therefore, the identification is based mainly on the morphological identification of male genitalia characters.10 , 11 , 12 , 13 DNA-based species identification is an alternative and can be used as a complementary tool. Torres-Gutierrez et al.14 verified that the 658 bp region of subunit I of the cytochrome c oxidase gene (DNA barcode)15 presented high resolution in the species delimitation of the subgenus Melanoconion. Talaga and Gendrin16 described three species of Culex (Mel.) based on both morphological characteristics of the male genitalia and molecular data of the COI gene. This mitochondrial marker is also effective in identifying cryptic Culex species.17 Demari-Silva et al.18 used a fragment of the COI gene to establish phylogenetic relationships among 17 species of the genus Culex. Although the results corroborate the monophyly of the subgenus Melanoconion, the authors suggested confirmation through further studies with nuclear genes and a greater number of samples, including species from the Pilosus Group and Spissipes Section. Torres-Gutierrez et al.19 verified the monophyly of the Melanoconion and Spissipes Sections through phylogenetic analyses with nuclear and mitochondrial genes. Although the results have been consistent with most of the morphological classification of Spissipes Section, the same was not true for Melanoconion Section. Only the Atratus and Pilosus Groups were monophyletic. Therefore, the authors report that the results for Melanoconion Section were inconclusive due to limited taxon representation and suggest future investigations with greater representation.
Mitochondrial genome has also been used to phylogeny in Culicidae.20 , 21 Demari-Silva et al.20 performed phylogenetic analysis with mitochondrial protein-coding genes using four of the six species of the Coronator Group of the subgenus Culex and observed the monophyly of this Group, corroborating with the morphological hypothesis. Mitochondrial genomes in Culex corroborates that of other metazoan organisms, presenting a length of approximately 15 Kb, with 13 protein coding genes (PCGs), two rRNA (rrnL and rrnS), 22 tRNA genes, and a region rich in adenine (A) and thymine (T).20 , 21 Mitogenome sequences of Culex mosquitoes of the subgenera Culex; Lophoceraomyia; Neoculex; and Culiciomyia are available in National Centre for Biotecnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/). Although mosquitoes of the subgenus Melanoconion are of medical importance and are used in phylogeny and taxonomy studies,14 , 19 information about the mitochondrial genome is scant. Phylogenetic analyses with mitochondrial protein-coding genes together with morphological data can contribute to a more accurate identification of the species of Melanoconion Section. Thus, this study aims to: (1) obtain, describe, and analyse mitochondrial protein coding genes from different species of the Educator and Atratus Groups of the subgenus Melanoconion; (2) verify the phylogenetic relationships of both Groups.
MATERIALS AND METHODS
Mosquito Sampling - Mosquitoes of the genus Culex subgenus Melanoconion from different localities of Brazil were used in this study. Details about mosquito collections are shown in Supplementary data (Table I). Specimens were identified according to the keys proposed by Sá et al.10 and Rodrigues de Sá et al.,11 placed in tubes containing 95% ethanol and stored at -80ºC. Male genitalia were dissected and mounted on microscope slide, covered with fine coverslip, and deposited in the Coleção Entomológica de Referência, Faculdade de Saúde Pública, Universidade de São Paulo. Genomic DNA from whole mosquitoes was individually extracted using the Qiagen DNeasy Blood & Tissue Kit (Qiagen), following the manufacturer's instructions. The extracted DNA was stored at -80ºC as part of the frozen entomological collection of the Faculdade de Saúde Pública, Universidade de São Paulo, Brazil.
Polymerase chain reaction (PCR) amplification and sequencing - The largest region of the mitochondrial genome between the 16S rRNA and cytochrome c oxidase subunit I (COX1) genes was amplified with the primers 16Sa (5′ CGCCTGTTTATCAAAACAT 3′)22 and LCO1490 (5′ GGTCAACAAATCATAAAGATATTGG 3′).23 This region encompasses 12 PCGs and has approximately 12 Kb. For each polymerase reaction was used GoTaq® Long PCR Master Mix 1x, 0.2 mM of each primer, 1 µL of DNA and ultrapure water to the final volume of 50 µL. Thermal cycler conditions were 94ºC for 2 min, 42 cycles of 94ºC for 30 s, 45ºC for 20 s and 65ºC for 13 min and a final extension at 72ºC for 7 min. Amplicons were purified using DNA Clean & Concentrador^TM^ (Zymo Research, California, USA) and quantified using a Qubit 2.0 fluorometer (LifeTechnologies, Oregon, USA), according to the manufacturer's instructions. Long PCR amplicon libraries were prepared using the Nextera® XT DNA Sample Preparation Kit (Illumina, Illinois, USA) and paired-end fragments (150 bp) were sequenced on the Illumina MiSeq platform. The quality of the generated reads was assessed using FastQC v0.11.9.24
Mitochondrial genome assembly and annotation - Mapping to reference method was used to genome assembly. Geneious Prime 2023.2.1 (https://www.geneious.com) was used for paired-end reads assembly using default parameters, mapper Geneious method and Culex quinquefasciatus mitochondrial genome sequence (Genbank accession NC_014574) as reference. Annotation of the genes was performed using MITOS 2 Web Server25 with invertebrate genetic code and confirmed manually with the alignment of each gene with Culex sequences available in Genbank (Genbank accessions NC_036006 and NC_014574).
Sequence analysis - All new sequences were aligned using ClustalW in MEGA 11.0.13 software.26 The GC-skews and AT-skews were measured using the following formulas: guanine-cytosine (GC-skews) (G - C) / (G + C) and adenine (A) and thymine (T) (AT-skews) (A - T) / (A + T). These values can range from -1.0 to 1.0 and indicate compositional asymmetries in DNA sequences. Nucleotide diversity (π) was generated in DnaSP v.6.12.0.3,27 using all mitochondrial sequences obtained and with a sliding window of 200 bp and steps of 25 bp. The ratio between non-synonymous (Ka) and synonymous (Ks) substitution in PCG sequences was also calculated. Values Ka/Ks were obtained from pairwise sequence comparisons of each PCG in DnaSP v.6.12.03. This analysis allows estimating whether certain PCG are under (1) positive selection (Ka/Ks > 1), (2) negative selection (Ka/Ks < 1), or (3) neutral evolution (Ka/Ks = 1).
The invertebrate mitochondrial genetic code was used to translate the PCG sequences in amino acid sequences. Nucleotide composition and relative synonymous codon usage (RSCU) were calculated using MEGA 11.0.13 software.
Phylogenetic analysis - An overview phylogeny of Culicidae was made using translations from samples of Culicidae species and the newly-sequenced Melanoconion species. Complete mitochondrial genomes from Culicidae were obtained from Genbank, which included 21 genera with at least one mitogenome. Two mitogenomes were sampled randomly from each such genus (with only one from Orthopodomyia, as that was all that was available), from which translations of protein coding genes were obtained. New Melanoconion sequences PQ389 and PQ409 from this study were added to the other Culicidae sequences. Alignments of the translations were made using Clustalo,28 but alignments for ATP8, ND3, and ND4L were too short (lengths 54, 118, and 99, respectively) and were not used. A concatenated alignment of the remaining nine translations was made. There were five pairs of sequences that were identical; one of each was removed from the alignment. The alignment of the remaining 38 sequences, with a length of 3137 characters, was analysed with Phylobayes MPI V1.9, using two runs, using the CAT+GTR+G(4) model.29 A phylogeny of the Melanoconion subgenus was made in a similar way to the Culicidae phylogeny described above. Alignments were made using protein-coding gene translations of the 23 new Melanoconion sequences from this study, to which were added Culex and other near outgroup sequences. This alignment of 46 sequences, of length 3127 AAs, was analysed as described above for Culicidae, using Phylobayes MPI V1.9 using the CAT+GTR+G(8) model.
RESULTS
Mosquito sampling - Of the total specimens used in this study, two were morphologically identified as Cx. dunni, two as Cx. ensiformis, three as Cx. theobaldi, three as Culex trigeminatus, two as Cx. eknomios, two as Cx. zeteki, one as Culex near commevynenis, two as Culex near vaxus, three as Cx. angularis, and three as Cx. longistriatus. All specimens were male except one Cx. zeteki [Supplementary data (Table I)].
PCR amplification and sequencing - A region of the mitochondrial genome of approximately 12 kb was amplified from each of the 23 specimens. Next-Generation sequencing generated a total of 2,684,992 paired-end reads. The number of reads generated for each specimen are in Supplementary data (Table II).
Mitochondrial genome assembly and annotation - After assembly, the sequences had length between 11,801 and 11,819 base pairs [Supplementary data (Table II)] and the PCGs were composed of 3,392 codons. The partial mitochondrial genome of all samples contains 28 genes, including 12 PCGs (COX1, COX2, ATP8, ATP6, COX3, ND3, ND5, ND4, ND4L, ND6, CYTB, and ND1), 15 tRNA, and 1 rRNA (rrnL), with partial sequences of COX1 and rrnL (Fig. 1). Protein coding and tRNA genes were located in both strands (H or L) and rrnL on the L-strand (Fig. 1). In H-strand are following PCGs and tRNA: COX1, COX2, ATP8, ATP6, COX3, ND3, ND6, CYTB, trnL2, trnK, trnD, trnG, trnR, trnA, trnN, trnE, trnT, trnS2, therefore in L-strand are: ND5, ND4, ND4L, ND1, trnS1, trnF, trnH, trnP, trnL1. The following nine mitochondrial genes were not sequenced: ND2 (PCG); trnI, trnQ, trnM, trnW, trnC, trnY, and trnV (tRNA); rrnS (rRNA).
diagram graphic of the mitochondrial region amplified by the primers mentioned in the study. The region comprises 12 protein coding genes
All PCGs of species sequenced possessed the start codon ATN, except for the COXI, which was not possible to determine due to the partial sequencing of this gene (Supplementary data (Table III)). The partial stop codon (T_) was observed in COX1 and COX2 genes, whereas ATP6, ATP8, ND1, ND3, ND4, ND4L, ND5, ND6, COX3 and CYTB showed a complete stop codon (TAA) [Supplementary data (Table III)].
The PCGs showed no length variation between the species studied. One difference was observed in the length of the ND5 gene in relation to the Cx. quinquefasciatus sequence (Genbank accession NC_014574), used as a reference for genome assembly. The ND5 gene of the new sequenced samples presented one less base, which generated a stop codon and, consequently, a shorter length compared to the NC_014574 sequence (Fig. 2). Because of the absence of this base occurred in a polyT region, Sanger sequencing was performed to verify whether this difference could have been a result of the assembly of the reads. So, the new sequences were aligned and primers in the ND3 and ND5 regions were designed (Primer F: TTGGACTTTATCATGAATG (ND3 region) and Primer R: GATCAAGGGTGAAGTGAA (ND5 region)) to amplify the fragment containing the polyT region mentioned above. Amplification and sequencing of this fragment (~ 622 bp) was performed in the following samples: PQ401 (IG20-5), PQ403 (IG20-6), and PQ408 (SP-Ciduni-02). The result of the alignment of the ND5 fragment between Sanger sequencing and Illumina sequencing can be seen in Supplementary data (Fig. 1), confirming the absence of the base.
schematic diagram showing the end of the ND5 gene nucleotide and amino acid alignment of Culex quinquefasciatus (NC_014574) with Culex theobaldi (PQ389). (A) Both nucleotide strands are sense direction. The red arrow indicates the lack of a base in PQ389. (B) The blue arrows indicate the stop codon of each specimen for ND5.
Sequence analysis - The nucleotide frequency in the fragment sequenced for each specimen can be seen in Table. All species showed the positive value for AT-skew and negative for GC-skew (Fig. 3), showing an excess of A over T and C over G.
graph of adenine-thymine (AT) skews and guanine-cytosine (GC) skews values for each species addressed in this study.
For all species under study, Leu and Ser were the most abundant amino acids and the most frequent codons (RSCU > 1.0) were those ending with A or T, as can be seen for Cx. dunni and Cx. near commevynenis in Fig. 4 and for the other species in Supplementary data (Table IV).
relative synonymous codon usage (RSCU) of the mitochondrial partial genome of species of Culex dunni and Culex near commevynensis. The RSCU values can observed on the y-axis.
Ka/Ks values ranged from 0.0 to 2.775 for species under study. Most mitochondrial PCGs presented Ka/Ks values lower than 1, indicating under negative selection pressure (Fig. 5). The ATP8 gene was the one that presented the greatest Ka/Ks variation, being greater than 1 in the following pairwise sequence comparisons: Cx. longistriatus (PQ404, PQ390, PQ393) x Cx. angularis (PQ401, PQ403, PQ408), indicating positive selection pressure [Fig. 5 and Supplementary data (Fig. 2)].
box plot of the Ka/Ks values of each protein coding gene of Culex (Melanoconion) species.
The degree of polymorphism between the different species of Culex (Mel.) was observed throughout the mitochondrial sequences by nucleotide diversity (π), which varied between 0.00407 and 0.12085 [Supplementary data (Table V)]. The most polymorphic PCGs were ND6, ND5, and COX1 (Fig. 6).
nucleotide diversity of mitochondrial partial genome of Culex (Mel.).
Phylogenetic analysis - Two phylogenetic analyses were performed using translations from PCGs. Only partial sequence of COX1 was considered in the analyses, since their complete sequence was not obtained in the 23 sequenced samples under study. The 12kb fragment from the new Melanoconion sequences from this study contained COX1 (partial), COX2, ATP8, ATP6, COX3, ND3, ND5, ND4, ND4L, ND6, CYTB, and ND1. However, ATP8, ND3, and ND4L were not used because they were, with lengths 53, 117, and 98 AAs respectively, deemed too short. The first tree has a broad phylogenetic range and shows where samples from the Melanoconion subgenus fit into the Culicidae family (Fig. 7). The second tree was performed using all 23 Melanoconion sequences plus sequences from close groups (Fig. 8). The Genbank accession number of the sequences used in both phylogenetic trees is indicated after each species name in Figs 7-8.
overview phylogeny of Culicidae made using translations from nine mitochondrial protein coding genes, analysed with Phylobayes MPI V1.9, using two runs. The maxdiff convergence diagnostic was 0.08, showing good topological convergence. However, there was evidence for compositional heterogeneity over the taxa (p = 0.0).
phylogeny of the Melanoconion subgenus, using translations from nine mitochondrial protein coding genes, analysed with Phylobayes MPI V1.9, using two runs. The maxdiff convergence diagnostic was 0.07, showing good topological convergence. Compared to the Culicidae analysis shown in Fig. 7 there was less evidence for compositional heterogeneity across taxa here in these closely-related sequences (p = 0.3, 0.5)
Phylogenetic analysis generated two well-supported (100% posterior probability) monophyletic clades. A clade with lineages of Culex (Culex), Culex (Neoculex), and Culex (Lophoceraomyia) and one sister clade with Cx. (Mel.). Within the Cx. (Mel.) clade, the subclade with species from the Educator Group of the Melanoconion Section was well supported (100%) with the subclade sister of lineages from the Atratus Group of the Melanoconion Section (Fig. 8).
The analysis showed three possible putative species, two (Culex near vaxus Form 1 and Culex near vaxus Form 2) resulting from 2 specimens morphologically identified as Cx. near vaxus and one from the specimen Cx. near commevynensis (Fig. 8).
DISCUSSION
Recently, reviews of the Atratus and Educator Group of the Melanoconion Section of Culex (Mel.) have been published.10 , 11 New species were described, species distribution was updated and morphological identification keys for different mosquito life stages were provided, contributing to the identification of species in these Groups.
Species from these Groups are vectors of different parasites to humans, and are therefore important in public health.30 Previous studies have obtained and analysed mitochondrial and nuclear gene sequences, showing that gene fragments can be used as a tool in the identifying these species.14 , 19
In this study, the mitochondrial sequence of approximately 12 kb was obtained from 23 mosquitoes, comprising eight valid species and three putative species of Culex (Mel.). This region encompassed 12 PCGs, 15 transfer RNA genes and rrnL, with partial sequences of COX1 and rrnL. Although there was no difference in the order of PCGs or tRNAs in relation to other mitochondrial genomes of Culex,20 , 31 a difference in the length of the ND5 gene was observed. Variations in mitochondrial DNA length in insects are generally associated with evolutionary events in the control region32 , 33 , 34 and intergenic region.20 Liu and Beckenbach35 analysed COX2 sequences in 10 insect orders and found a variation in size (673 bp - 690 bp), resulting in between 226 and 229 amino acids. Internal insertions and deletions were observed, but variations length were more frequent in at or near the 3' end of gene. In the present study, all sequenced species showed deletion of a base near the 3' end of the ND5 gene, which resulted in the formation of a stop codon and, consequently, a lower number of amino acids. Little is known about the mitochondrial genes of Culex (Mel.), and therefore, further studies need to be carried out to verify whether this variation can (1) be characteristic of the studied Groups (Atratus and Educator), of the Melanoconion Section or of the subgenus Melanoconion, (2) have modified the functions and efficacy of the ND5 gene.
Partial stop codons (T__) present in COX1 and COX2 genes in this study were observed in others Culex species and insects.36 , 37 , 38 The presence of these codons is common in mitochondrial PCGs, which are added with adenine by polyadenylation.39 All species showed positive values for AT-skew and negative values for GC-skew. This result is in agreement with other studies with Culex, Haemagogus, and Anopheles.20 , 40 , 41
The importance of mitochondrial DNA in the respiratory chain process is well known, as is the high mutational pressure on it in metazoans.42 Previous studies have reported positive selection (Ka/Ks > 1) in insect mitochondrial PCGs, highlighting its role in adaptation to different environments. Analysis of the mitochondrial genomes of Anopheles stephensi and Anopheles dirus revealed evidence of positive selection in the ND2, ND4, and ND6 genes.43 Similarly, positive selection was detected in the ATP8 gene (Ka/Ks = 1.65) among hemipteran species.44 In addition, seven of the 13 mitochondrial PCGs in flying grasshopper lineages appear to be under positive selection, which may have facilitated adaptation to the high energy demands of sustained flight during periods of atmospheric oxygen reduction.45 In the current study, of the 12 mitochondrial PCGs, 11 presented Ka/Ks < 1 (negative selection), as observed in other studies with culicids.20 , 36 , 41 The ATP8 gene presented Ka/Ks > 1 only in the pairwise sequence comparisons Cx. longistriatus (PQ404, PQ390, PQ393) x Cx. angularis (PQ401, PQ403, PQ408). These specimens were recovered as sister species in the Bayesian topology (Fig. 8) and further studies need to be carried out to verify whether that selection was fundamental for the adaptation of the species to the environment.
Protein coding and non-coding sequences are used for studies involving Culex (Mel.) species. Torres-Gutierrez et al.19 used COX1 and nuclear genes to infer the phylogeny of species from the Spissipes and Melanononion Sections of Culex (Mel.) and verified that the phylogenetic signal of these genes is greater when analysed together than separately. Navarro and Weaver46 suggested a cryptic species similar to Culex pedroi by analysis of the internal transcribed spacer 2 (ITS2) region. In the present study, Bayesian analysis showed that the sequence of the analysed PCGs is highly informative for the separation not only of the Educator and Atratus Groups, but also for species of the others subgenera and genera. Furthermore, this analysis suggested the presence of three putative species, one most morphologically similar to Cx. commevynensis and the others to Cx. vaxus. The sequences generated may contribute to future studies of the Melanoconion subgenus of Culex.
Thus, Cx. dunni, Cx. ensiformis, Cx. theobaldi, Cx. trigeminatus, Cx. eknomios, Cx. zeteki, Cx. near commevynenis, Cx. longistriatus, Cx. angularis, and Cx. near vaxus showed the same pattern of PCGs, tRNA, and rRNA as species of the same genus. The PCGs used in the phylogenetic analyses provided good support for monophyletic clades. Future molecular studies will be needed to confirm whether the size difference in the ND5 gene is characteristic of the subgenus Melanoconion, and morphological studies will be essential for the analysis of the possible putative Culex species.
SUPPLEMENTARY MATERIALS
Supplementary material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ferro C Boshell J Moncayo AC Gonzalez M Ahumada ML Kang W Natural enzootic vectors of Venezuelan equine encephalitis virus in the Magdalena Valley, Colombia Emerg Infect Dis 20039149541253328110.3201/eid 0901.020136 PMC 2873762 · doi ↗ · pubmed ↗
- 2Cupp EW Zhang D Yue X Cupp MS Guyer C Sprenger TR Identification of reptilian and amphibian blood meals from mosquitoes in an eastern equine encephalomyelitis virus focus in central Alabama Am J Trop Med Hyg 200471327227615381805 PMC 1351276 · pubmed ↗
- 3Yanoviak SP Aguilar PV Lounibos LP Weaver SC Transmission of a Venezuelan equine encephalitis complex alphavirus by Culex (Melanoconion) gnomatos (Diptera: Culicidae) in northeastern Peru J Med Entomol 20054234044081596279410.1093/jmedent/42.3.404 · doi ↗ · pubmed ↗
- 4Turell MJ Dohm DJ Fernandez R Calampa C O'Guinn ML Vector competence of Peruvian mosquitoes (Diptera: Culicidae) for a subtype IIIC virus in the Venezuelan equine encephalomyelitis complex isolated from mosquitoes captured in Peru J Am Mosq Control Assoc 200622170751664632510.2987/8756-971X(2006)22[70:VCOPMD]2.0.CO;2 · doi ↗ · pubmed ↗
- 5López RH Soto SU Gallego-Gómez JC Evolutionary relationships of West Nile virus detected in mosquitoes from a migratory bird zone of Colombian Caribbean Virol J 201512802598990110.1186/s 12985-015-0310-8PMC 4445300 · doi ↗ · pubmed ↗
- 6Nunes Neto JP Reis LAM Freitas MNO do Nascimento BLS das Chagas LL da Costa HHM First isolation and genome sequence analysis of West Nile virus in mosquitoes in Brazil Trop Med Infect Dis 2023842373710436210.3390/tropicalmed 8040237 PMC 10143329 · doi ↗ · pubmed ↗
- 7Ramos B Carvalho V da Silva E Freitas M Barros LJ Santos M The First isolation of insect-specific alphavirus (Agua Salud alphavirus) in Culex (Melanoconion) mosquitoes in the Brazilian Amazon Viruses 202416913553933983210.3390/v 16091355 PMC 11436152 · doi ↗ · pubmed ↗
- 8Torres-Gutierrez C Sallum MA Catalog of the subgenus Melanoconion of Culex (Diptera: Culicidae) for South America Zootaxa 2015402811502662429510.11646/zootaxa.4028.1.1 · doi ↗ · pubmed ↗