Hsp90 buffers behavioral variability by regulating Pdf transcription in clock neurons of Drosophila melanogaster
Angelica Coculla, Carlina Feldmann, Maite Ogueta, Sina Mews, Roland Langrock, Ralf Stanewsky

TL;DR
The protein HSP83 in fruit flies helps regulate circadian behavior by controlling the production of a key neuropeptide, which may allow flies to adapt to stressful conditions by expanding their active time.
Contribution
This study reveals a novel role for HSP83 in buffering behavioral variability by regulating the transcription of the circadian neuropeptide PDF in clock neurons.
Findings
HSP83 mitigates inter-individual behavioral variability in a subset of clock neurons.
Reduced HSP83 function leads to decreased levels of the circadian neuropeptide PDF.
Hsp83 mutants show disrupted synchronized oscillations of the clock protein PERIOD.
Abstract
Circadian rhythms are prevalent on Earth and temporally organize behaviour and physiology of organisms to occur in species-specific ‘temporal niches’. However, species differ in how strictly individuals are controlled by their circadian clock, suggesting that it may offer a selective advantage for an individual to extend its temporal niche under certain circumstances, for example during stressful environmental conditions. A potential mechanism controlling temporal niche adherence involves the evolutionary capacitor and chaperon protein HSP90, known to assist the proper folding of important signalling molecules. If HSP90 becomes rate limiting (e.g., under environmental stress) hidden genetic variation will be expressed, producing novel and potentially beneficial phenotypes for the individual. While this role of HSP90 is well established for morphological traits, we show here that it…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
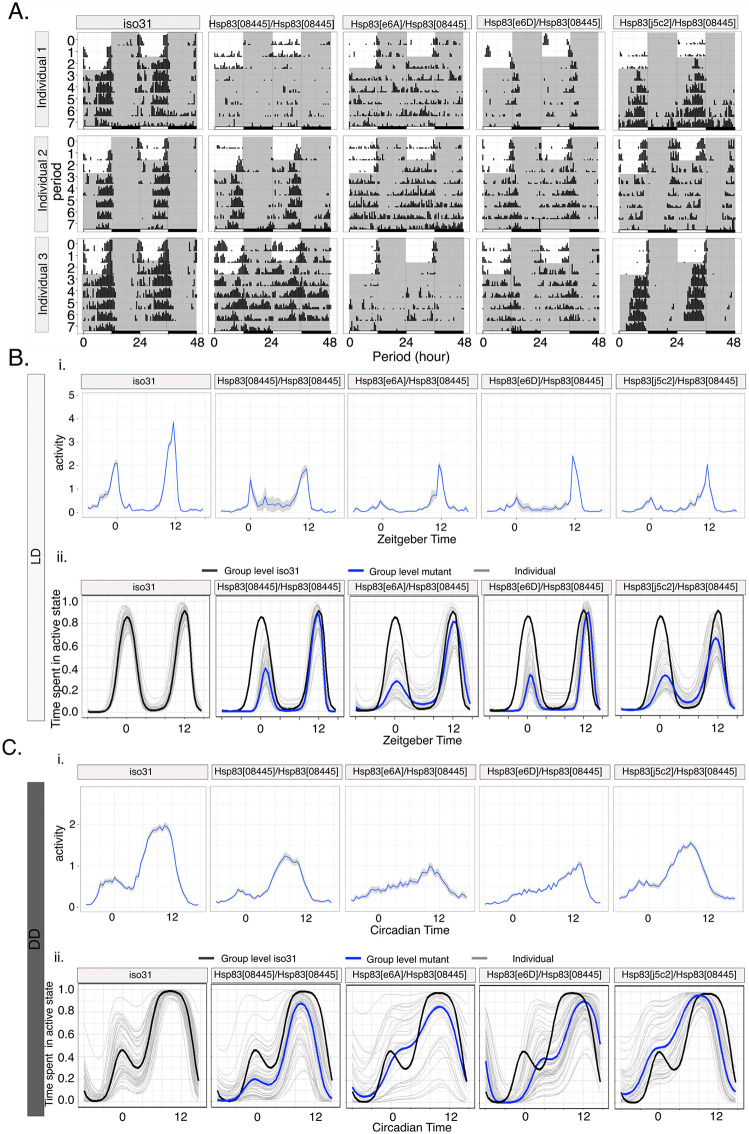 Fig 1
Fig 1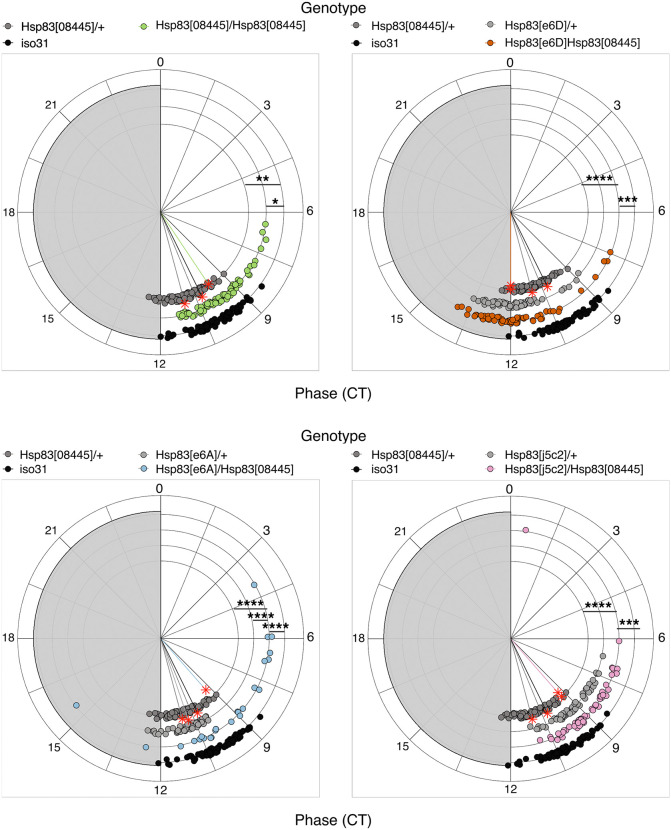 Fig 2
Fig 2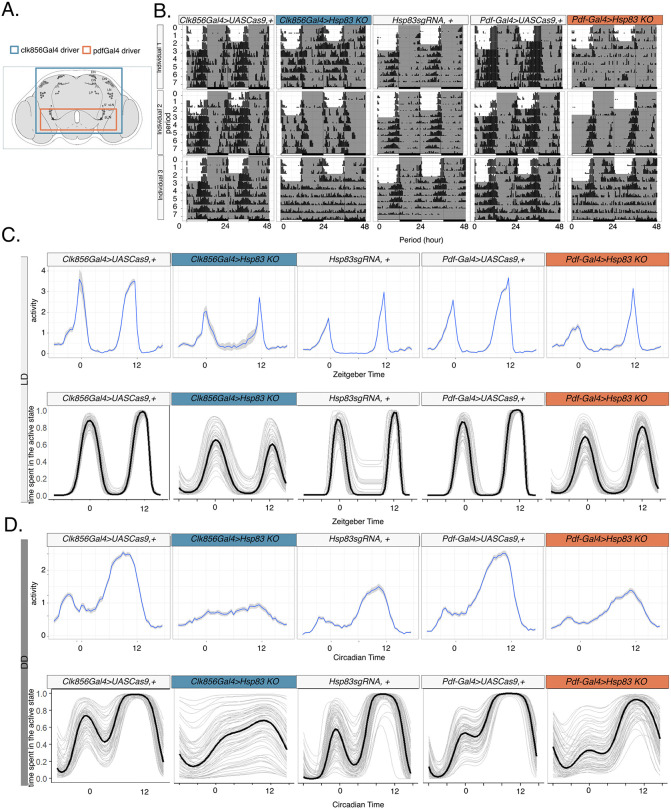 Fig 3
Fig 3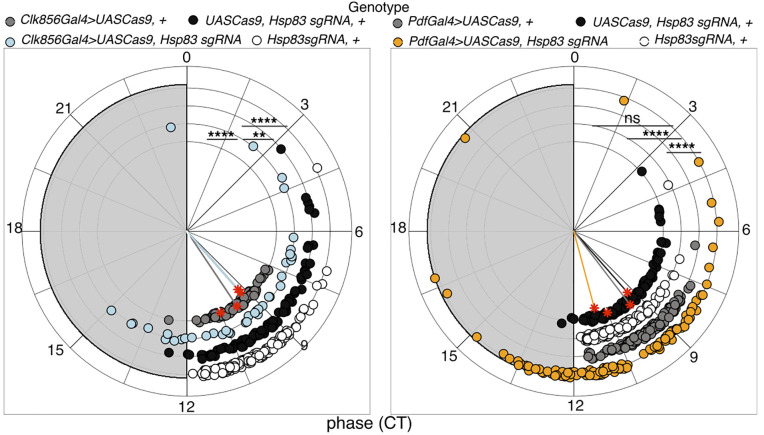 Fig 4
Fig 4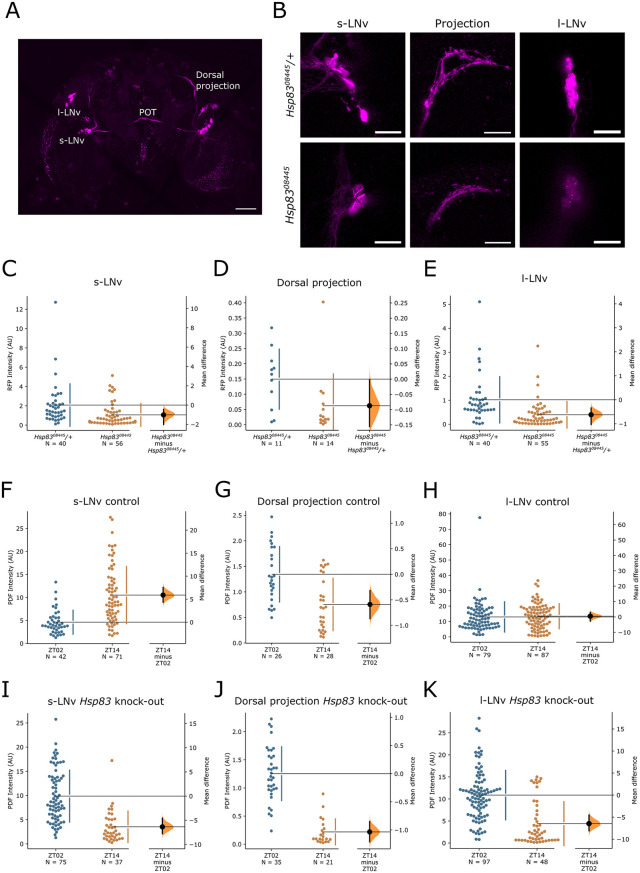 Fig 5
Fig 5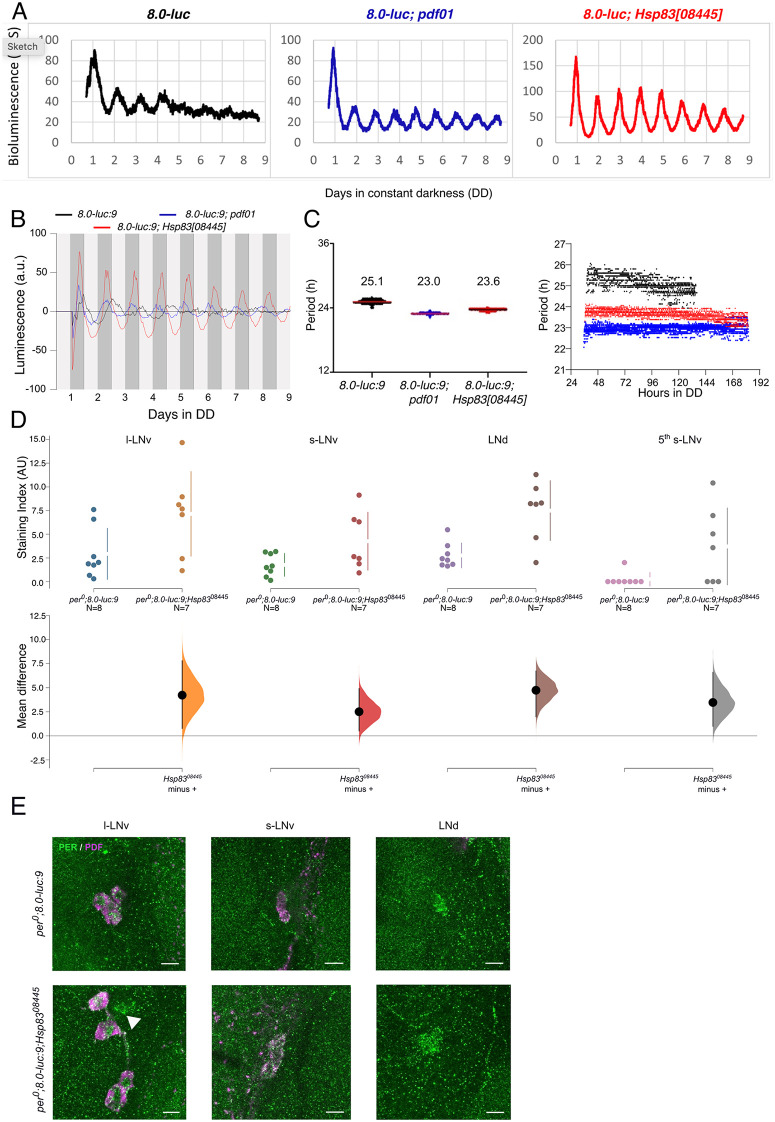 Fig 6
Fig 6- —http://dx.doi.org/10.13039/501100001659Deutsche Forschungsgemeinschaft
- —http://dx.doi.org/10.13039/501100001659Deutsche Forschungsgemeinschaft
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircadian rhythm and melatonin · Neurobiology and Insect Physiology Research · Genetics, Aging, and Longevity in Model Organisms
Introduction
Animals restrict their behavioral activities to specific times of day or night, and these behaviours are regulated by self-sustained circadian clocks. Environmental rhythms, like the daily light:dark and concomitant temperature changes synchronize the circadian clock with the external time. This allows animals to exert behaviours at optimal times of day, often by anticipating the actual environmental change. Circadian clocks therefore contribute to overall fitness, for example through minimizing energy expenditure, increasing mating probability, or avoiding predators [1]. In Drosophila melanogaster, daily activity rhythms are orchestrated by a network of ~240 clock neurons in the fly brain [2]. Within these neurons transcriptional and translational molecular feedback loops of key circadian clock genes generate ~ 24 h molecular oscillations, which are translated into neural activity rhythms, rhythmic neuropeptide release, as well as into rhythmic activation of clock-controlled output genes, ultimately regulating rhythmic behaviour and determining the crepuscular temporal niche of the fruit fly [3,4]. The core feedback loop is initiated by a heterodimer formed of the two transcription factors CLOCK (CLK) and CYCLE (CYC), which binds to E-box elements upstream of the period (per) and timeless (tim) genes, thereby activating their transcription. PER and TIM proteins gradually accumulate in the cytoplasm and eventually enter the nucleus to repress CLK-CYC activity and per/tim transcription. After degradation of PER and TIM, CLK-CYC can reinitiate per and tim transcription and the feedback loop repeats itself every 24 h. CLK and CYC are also part of an interlocked feedback loop, in which they activate the transcription of the genes encoding the basic-zipper transcription factors VRILLE (VRI) and PAR DOMAIN PROTEIN 1ε (PDP1ε), which repress, or activate Clk transcription, respectively [4]. Inactivating vri or Pdp1ε in the clock neurons abolishes locomotor activity rhythms without interfering with molecular clock function, demonstrating that the interlocked feedback loop is mainly responsible for clock output [5,6,7]. The neuropeptide pigment dispersing factor (PDF) is expressed in a subset of the clock neurons, four small as well as four large ventral lateral neurons (s-LNv and l-LNv) and required for maintaining behavioural rhythmicity in constant conditions [8]. While PDF is not required for the maintenance of molecular oscillations in the clock neurons, it does coordinate the phase and amplitude between different neuronal groups, which is essential for maintaining rhythmic behaviour in constant dark conditions [9,10]. In turn, PDF expression is post-transcriptionally regulated by the circadian clock via VRI, which is required for normal accumulation of Pdf RNA and peptide levels. As a consequence, the daily plasticity rhythms of the s-LNv terminals in the dorsal brain are abolished, most likely contributing to the behavioural arrhythmicity of vri mutant flies [6].
Interestingly, despite the remarkable conservation of circadian clock genes and mechanisms in the animal kingdom, the degree of rhythmically behaving individuals within a certain species is surprisingly variable. While in D. melanogaster nearly each individual displays robust behavioural rhythms, we have recently shown that in the flour beetle Tribolium castaneum only 30–50% of the individuals behave rhythmically [11]. This difference between species may allow for adjustments of circadian clocks to the specific ecological demands of a particular species. For example, under environmental stress (e.g., drought, excessive heat), it could be advantageous not to be restricted to a narrow temporal niche, but to extend it, or even to be active at irregular times. In a population, these individuals would have a fitness advantage under certain environmental conditions, an evolutionary strategy termed as bet hedging [12]. On the other hand, even a largely homogenously rhythmic species like D. melanogaster could possess buffered standing genetic variation. Such cryptic genetic variation (CGV) would only emerge under stressful environmental conditions, leading to inter-individual behavioural variability. For morphological traits, the evolutionary capacitor HSP90 has been implicated in the release of such CGV, for example in fruit flies and cave fish [13,14]. Unlike HSP60 and HSP70 proteins, which are involved in normal protein folding, HSP90 is a chaperon protein assisting in the folding of important signalling molecules, such as transcription factors, kinases, and ubiquitin ligases [15]. Under non-stressful conditions, HSP90 is not rate-limiting, allowing correct protein folding, even in the presence of mild hypomorphic missense mutations in the genes encoding HSP90 target proteins. Protein folding is sensitive to environmental stress and HSP90 becomes rate limiting under these conditions, thereby unmasking such cryptic genetic polymorphisms, with potential beneficial phenotypes for the individual [13,14]. For example, reducing HSP90 function in Drosophila led to the appearance of morphological eye, bristle and wing phenotypes, which--following selection--persisted even after restoring HSP90 function [14]. There is also evidence that HSP83 acts as chaperon for epigenetic factors, notably proteins encoded by the trithorax group (TrxG) genes, thought to be involved in the maintenance of active chromatin states [16,17]. In support of this idea, it was shown that HSP90 depletion also resulted in morphological variability in isogenic Drosophila melanogaster and Arabidopsis thaliana plants, together suggesting an epigenetic mechanism for HSP90 capacitor function [17]. In contrast to morphological traits, it is questionable whether release of CGV or misfolding of epigenetic factors by limited availability of HSP90, also contribute to behavioural variability. An increase of behavioural variability under stressful environmental conditions by either mechanism would allow certain animals of a population to extend their temporal niche, potentially offering a selective advantage for these individuals. A previous study conducted in Drosophila pointed to a potential role of HSP90 (HSP83 in flies) as capacitor of behavioural variation in circadian locomotor activity [18]. Reduction of HSP83 resulted in a marked increase of inter-individual behavioural variation without apparent alterations of the underlying molecular clock oscillations [18]. Here, we aimed to identify the cellular substrates of circadian HSP83 function. Using an inducible HSP83 CRISPR allele, we show that behavioural variability increases when Hsp83 depletion is restricted to clock neurons only. Hsp83 is required for normal Pdf transcription and accumulation of PDF in the s-LNv cell bodies and projections. Strikingly, Hsp83 mutation resembles the effect of Pdf^01^ loss-of-function mutants on PER expression rhythms in subsets of PDF-negative neurons, indicating faulty synchronisation between different clock neuronal groups. Our results show that HSP83 is required for normal PDF levels throughout the day, and suggest that this chaperon is important for the proper folding of proteins that regulate PDF. Interestingly, it has recently been shown that DNA sequence differences in the non-coding, upstream region of the Pdf gene between two Drosophila species (D. sechellia and D. melanogaster) translate into different PDF peptide levels and distinct behavioural patterns [19]. Therefore, transcriptional regulation of a single circadian neuropeptide gene (Pdf), potentially contributes to behavioural circadian variation between different species and between individuals of a population.
Results
Hsp83 mutants exhibit inter-individual behavioral variability
To confirm that interference with Hsp83 function indeed increases behavioral variability of circadian behaviors, we analyzed locomotor activity of Hsp83 alleles previously reported to affect behavior [18]. We used the same three loss-of-function alleles (Hsp83^e6A^, Hsp83^e6D^, and Hsp83^j5c2^) and one hypomorphic allele (Hsp83^08445^). As expected, all loss-of-function alleles were homozygous lethal, and no transheterozygous offspring was obtained after crossing them to a deficiency removing the entire Hsp83 locus (S1 Table). In contrast Hsp83^08445^ flies are homozygous viable and generate viable transheterozygous offspring after crossing to the deficiency and the loss-of-function alleles ([18], S1 Table).
We tested the locomotor behavior of the homozygous and transheterozygous Hsp83 mutants and observed significant changes of the rhythmic locomotor activity patterns in the mutants compared to the controls (wild type flies and Hsp83^mut^/+) both in LD and DD (Figs 1A–C and S1). While control flies showed similar patterns in locomotor activity during entrainment and free running condition, the homozygous and trans-heterozygous offspring showed significant intra- and inter-individual differences (Figs 1A–C and S1). At the population level, mutants have less well-defined activity peaks compared to their respective controls in both LD and in DD (Figs 1A–C, S1A, and S1B).
Reducing Hsp83 function increases behavioral variability in Drosophila melanogaster.A) Representative actograms of Hsp83 mutant flies compared to iso31. White areas represent ‘lights on’ and gray areas ‘lights-off’. B, C) Population Activity (top) and HMM-implied percentage of time spent in the active state (bottom) of Hsp83 mutants (group and individual) and their controls (group) in LD (B) and DD (C). C) (experimental repeats: 4). See also Tables 1, 2, S2 and S3.
Table 1: Analysis of locomotor activity during LD.
Table 2: Analysis of locomotor activity in DD.
Hsp83 mutants increase behavioural variability during light:dark cycles
Inspection of the average activity plots showed that Hsp83 mutants display an amplitude reduction of the morning (M) and evening (E) peaks, indicating an impairment of synchronization to LD cycles, presumably caused by increased behavioural variability (Fig 1B). To quantify this behavior, we calculated an Entrainment index (EI), reflecting the ratio of M and E peak activity over the activity during the 12 hours flanking the light transitions in the morning and evening, respectively ([20], Materials and Methods). Compared to the wild type controls, M and E peak synchronization was reduced in all Hsp83 mutant combinations, particularly around lights on (Table 1). Moreover, in two heterozygous loss-of-function mutants (Hsp83^e6A^/ + , and Hsp83^e6D^/+) morning synchronization was reduced, which most likely is attributable to the generally higher M-peak variability, and indicating a dominant effect of Hsp83 depletion (Table 1). There was no significant difference in the rhythmic power of flies in LD (S2 Table). To investigate inter-individual variability more comprehensively, we analyzed LD and DD behavior separately using hidden Markov models (HMMs) as an alternative approach. An HMM is a statistical model that allows to identify not directly observable states (i.e., hidden states) based on the observable data, in our case locomotor activity counts [21]. These models are frequently applied in animal ecology to infer unobserved behavioral processes (e.g., active and inactive) from observed time series data (e.g., tracking data or locomotor activity) [22,21,23], and have been previously used with activity data in Drosophila to identify sleep stages [24]. Here, we used HMMs to distinguish two states: active and inactive (see Materials and Methods for details). Comparing average activity histogram plots with HMM analysis, which in contrast to total average activity reflects the proportion of flies in the active or inactive state, revealed the latter accurately reports the crepuscular behaviour of wild type flies displayed during LD cycles (Fig 1B, compare top and bottom graphs) [21]. Focusing on the model-implied percentage of time spent in the active state shortly before and after each environmental transition, we observed that overall Hsp83 mutants were more likely to be variable compared to the wild type (Figs 1B and S1A and Table 1). Interestingly, the time spent in the active state was more variable within the population also in the heterozygous mutant flies, with the exception of Hsp83^08445^/+ and Hsp83^e6A^/+ (Table 1). In summary, the HMM analysis revealed that inter-individual behavioral variability around the lights-on and lights-off transitions is increased in all Hsp83 mutants analyzed here. Moreover, at the population level this variability results in drastically reduced M-peak activity in Hsp83 mutants, suggesting that most of the mutant flies are not in the active state around lights-on (Figs 1B and S1A and Table 1).
Hsp83 mutants increase within-group behavioral variability in constant darkness
Inspection of individual actograms also revealed variable activity patterns between Hsp83 mutant individuals in DD (Fig 1A). To quantify this variability, we first analyzed rhythmicity and period-length, because previous studies reported a significant reduction of rhythmicity in Hsp83 mutant males in DD [25,18]. In our hands, Hsp83 mutant males did not show a reduction in overall rhythmicity compared to the controls, except for Hsp83^e6A^/Hsp83^08445^ flies, which exhibited a ~ 20% reduction and reduced rhythmic strength (Table 2). Moreover, as previously reported, all Hsp83 mutant combinations showed normal free-running period lengths (Table 2, [25,18]), indicating that core clock function is not severely affected. Although overall rhythmicity appears not, or only mildly affected, the individual and group activity plots indicate substantial inter-individual variation of the DD activity pattern in Hsp83 mutants (Figs 1A, 1C and S1B). To determine if variability under constant conditions is indeed increased in Hsp83 mutants, we quantified the activity phase of each individual on two consecutive days. For wild type flies, activity peaks on consecutive days will occur around the same time, leading to a high degree of ‘phase coherence’ within the population. Based on the sometimes erratic behavior observed in individual actograms of Hsp83 mutants (Fig 1A), we expected to see a reduction in phase coherence in the mutants. Indeed, inspection of the circular phase plots revealed that all Hsp83 mutant combinations showed reduced phase coherence (length of the colored vector emerging from the center of each circle) (Fig 2 and S3 Table). Increased inter-individual variability was independently supported by the HMM analysis, because Hsp83 mutants exhibit increased variability of the model-implied percentage of time spent in the active state around the subjective evening peak (Figs 1C and S1B and S3 Table). Combined, the results show that Hsp83 mutant individuals display most of their activity at different times during the 24-hr day.
Hsp83 mutants decrease phase coherence between individuals in constant darkness.Circular phase plots showing the peak phase in DD, calculated using the second (DD2) and the third day (DD3) in constant darkness. Each Hsp83 mutant allele was compared with the corresponding parental control and with wild type (iso31). For all transheterozygous combinations, homozygous Hsp8308445 females were used as mothers (including Hsp8308445/+) (experimental repeats: 4). Arrhythmic flies were excluded from the peak analysis. Colored lines represent the median phase of each group, while the length of the vector represents the circular standard deviation of the corresponding group (1/circular SD) (i.e., the phase coherence) (S3 Table). Watson’s goodness of fit test and pairwise equal kappa (κ) test (S5 and S6 Tables). Note: * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
To investigate, if the increased behavioural variability is indeed a consequence of HSP83 depletion, and to home in on the cellular substrate for HSP83 function in circadian behaviour, we performed rescue experiments expressing Hsp83 in all clock neurons in homozygous Hsp83^08445^ mutant flies. Using the Clk856-Gal4 driver, which is expressed in all clock neuronal groups to drive UAS-Hsp83, restored phase coherence in Hsp83^08445^ mutant flies to the level of control flies (S2A Fig and S3, S5 and S6 Tables). This points to a role for HSP83 within central brain clock neurons for properly phased circadian behaviour (see below).
Hsp83 knock out in clock neurons increases behavioral variability
The PDF neuropeptide influences synchronization between different clock neuronal groups [9,10], raising the possibility that Hsp83 depletion affects PDF, which in turn would result in desynchronized behavior. We therefore spatially restricted the disruption of Hsp83 function, by expressing a ubiquitously expressed Hsp83-sgRNA line together with UAS-Cas9 in clock neurons, thereby generating clock-neuron-specific Hsp83 mutations [26]. Using the timeless-Gal4 driver, which is expressed in all peripheral clock cells in addition to clock neurons in the brain, resulted in no viable offspring (S1 Table). This indicates that the CRISPR/Cas9 meditated knock-out effectively disrupts Hsp83 gene function, resulting in lethality as in the Hsp83 loss-of-function mutants described above (S1 Table). Next, we restricted Hsp83 knock-out to all (Clk856-gal4), or to the PDF-expressing subset of clock neurons (Pdf-Gal4) (Fig 3A) [27,8], resulting in viable offspring in both cases, as expected (S1 Table).
Hsp83 knock-out in clock neurons increases behavioral variability.A) cartoon of the adult clock neurons in the fly brain and neurons expressing the indicated Gal4 drivers. B) Representative actograms of Hsp83 knock-out and control flies. White areas indicate times when the lights were on, and dark areas, times when the lights were off, respectively. C, D) Population Activity (top) and HMM-implied percentage of time spent in the active state (bottom) of flies with Hsp83 knock-out in all clock neurons (Clk856-Gal4 > UAS Cas9, Hsp83-sgRNA), or the PDF expressing subset only (Pdf-Gal4 > UAS Cas9, Hsp83-sgRNA) and their controls in LD (C) and DD (D) (see also Table 1 and S3 Table) (experimental repeats: 6).
Clock-neuron specific Hsp83 knock-out resulted in aberrant and variable temporal locomotor activity patterns in LD and DD conditions, similar as described for the Hsp83 mutants above (Fig 3B–3D). EI quantification and HMM analysis of the behavior during LD revealed that, synchronization of the M- and E-peaks was impaired, both after Hsp83 knock-out in all, or the PDF-clock neurons only (Fig 3C and Table 1). As for the mutants, there was no significant difference in the rhythmic power of flies in LD (S2 Table). Interestingly, in DD Hsp83 knock-out in all clock neurons reduced the rhythmicity by ~50% compared to control flies, while there was only a very subtle effect (~6% reduction) after Hsp83 knock-out in the PDF neurons (Table 2). As observed for the Hsp83 mutants, CRISPR-induced knockout in all, or the PDF neurons only, had no effect on period length (Table 2). Next, we analyzed if Hsp83 knock-out also affected phase coherence in DD, as indicated by the variable behavior in individual actograms (Fig 3B). Indeed, circular phase and HMM analysis revealed that Hsp83 knock-out in all, or only in the PDF subset of clock neurons significantly decreased phase coherence, demonstrating an increase of inter-individual behavioral variability (Figs 3D and 4 and S3 Table).
Hsp83 knock-out in clock neurons decreases phase coherence between individuals.Circular phase plots showing the peak phase in DD for each individual, calculated using the second (DD2) and the third day (DD3) in constant darkness. Phase of flies with Hsp83 knock-out in all clock neurons (Clk856-Gal4 > UAS Cas9, Hsp83-sgRNA) (left), or the PDF expressing subset only (Pdf-Gal4 > UAS Cas9, Hsp83-sgRNA) and their controls (experimental repeats: 4-9). Arrhythmic flies were excluded from the peak analysis. Colored lines represent the median phase of each group, while the length of the vector represents the circular standard deviation of the corresponding group (1/circular SD) (i.e., the phase coherence) (see also S3 Table). Watson’s goodness of fit test and pairwise equal kappa (κ) test (S5 and S6 Tables). Note: * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
HSP83 depletion impairs Pdf transcription and PDF accumulation in s-LNv neurons
Because Hsp83 depletion restricted to the PDF-expressing subset of clock neurons is sufficient to induce behavioural variability, we investigated if PDF expression is affected by HSP83. To analyze potential effects on Pdf transcription, we applied the Pdf-Red reporter in which a 0.6 kb Pdf promoter-fragment drives the expression of the monomeric and soluble red fluorescence protein mRFP1 [28]. Based on a previous report showing that a similar Pdf-tdTomato construct, which contains ~2 kb of additional 5’ regulatory genomic DNA sequences, reports Pdf transcription [29], Pdf-Red can be considered as transcriptional reporter. Although Pdf mRNA levels do not oscillate across the day, we dissected flies at ZT2 and quantified RFP intensity in the soma of the l-LNv, the s-LNv, and in the dorsal s-LNv projections. Homozygous Hsp83^08445^ mutant brains showed the typical arborization pattern of the PDF-expressing LNv in the dorsal brain and the optic lobes, indicating that HSP83 is not required for the development of these neurons and circuit formation (Fig 5A). However, compared to wild type and heterozygous mutant control flies, homozygous Hsp83^08445^ mutant flies exhibited reduced RFP signals in the s- and l-LNv soma, as well as in the dorsal s-LNv projections, indicating that Hsp83 depletion results in reduced transcription of the Pdf gene (Figs 5B–5E and S3A and S7 Table). To rule out the possibility that Hsp83 depletion impairs RFP stability or folding, we also tested the effects of Hsp83 depletion in flies expressing mRFP1 in the CCAP neurons (S3B–S3C Fig). The results show no difference in signal intensities within the CCAP soma and projections, indicating that HSP83 is not involved in RFP folding or stability (S3B–S3C Fig and S7 Table). To determine effects of HSP83 depletion on PDF peptide levels, we dissected brains of flies with Hsp83 knock-out in all clock neurons (Clk856 > Cas9, hsp83 sgRNA). Anti-PDF stainings revealed that Hsp83 disruption in the LNv did not interfere with the normal development of these clock neurons, nor with the s-LNv projections into the dorsal brain (S4 Table). Dorsal s-LNv projections of wild type flies show higher PDF levels in the early morning compared to the early evening [30] (Figs 5G and S3D and S7 Table). Interestingly, in Hsp83 knock-out flies this distribution was maintained and the amplitude of PDF oscillations was increased due to lower trough levels at ZT14 (Figs 5J and S3D). PDF levels in the s-LNv soma of wild type flies peak twice, once during the day (~ZT2-ZT8) and a second time during the night (~ZT14-ZT22), whereby the peak levels at night are higher compared to those during the day [30] (Figs 5F and S3D). Interestingly, in Hsp83 knock-out flies, peak PDF levels occur in the morning, and trough levels are observed at ZT14 (Figs 5I and S3D). Considering that PDF release from the dorsal projections occurs during the daytime, our results indicate that in Hsp83 knock-out flies--after its release--PDF is not restored to normal levels during the night, presumably due to reduced Pdf transcription (Figs 5C–5E and S3D). Consistent with this interpretation, after Hsp83 depletion we also observe lower PDF levels at ZT14 in the l-LNv soma compared to controls, which show equally high PDF levels during the day and night [30] (Figs 5H, 5K and S3D). Reduced Pdf transcription could also contribute to the weaker phenotypes observed after Pdf-Gal4 compared to Clk856-Gal4 mediated Hsp83 depletion (Table 2), because Pdf-Gal4 transcription would be equally affected as transcription of the endogenous Pdf gene. Finally, VRI is crucial for PDF accumulation in the dorsal s-LNv projections, and this regulation of PDF levels occurs at the posttranscriptional level [6]. To rule out that the reduced PDF levels we observed at ZT14 are due to altered VRI levels in Hsp83 mutants, we performed anti-VRI stainings in the LNv of Hsp83^08445^ and control brains at ZT20 when VRI expression is high in wild type flies [6] (S3E Fig). We observed no differences in VRI intensity between mutant and wild type LNv, further supporting a role for Hsp83 in the transcriptional regulation of Pdf (S3E Fig and S7 Table).
Hsp83 regulates Pdf transcription and PDF levels.A) Whole brain image of a Pdf-Red; Hsp8308455/Hsp8308455 fly, showing the s-LNv, l-LNv and the dorsal and contralateral s-LNv and l-LNv projections, respectively. POT: Posterior Optic Tract. Scale bar: 50µm. B) Representative images of the s-LNv soma, their dorsal projections and the l-LNv soma of the heterozygous and homozygous mutant Hsp8308445 flies. Scale bar: 20 µm. C-E) Quantification and Estimation Statistics (ES) of the RFP levels for heterozygous and homozygous mutant Hsp8308445 flies for the s-LNv (C), dorsal projections (D) and l-LNv (E) (see S3A Fig for a comparison to wild type flies). F-H) Quantification and ES of PDF expression in the control Hsp83-sgRNA/ + flies in the somas of the s-LNv (F), dorsal projections (G), and l-LNv (H). I-K) Quantification and ES of PDF expression in Hsp83 knock-out flies (Clk856-Gal4 > UAS Cas9, Hsp83-sgRNA) in the s-LNv (I), dorsal projections (J) and l-LNv (K) (Representative images in S3D Fig). n-numbers are indicated below each genotype. For the somas, data from single neurons are represented, and for the projections, n refers to hemispheres. In each of the panels (C-K), the left plot represents all the datapoints, with the mean (size of gap between vertical lines) and the standard deviation (length of the vertical bars) for each condition to the right. The right plot shows the mean difference of the data from the two conditions as a bootstrap 95% confidence interval (see Materials and Methods for details).
Hsp83 and Pdf interact genetically
The effect of Hsp83 depletion on Pdf described above, points to a specific interaction between both genes. To test this interaction genetically, we crossed Hsp83 mutant flies to Pdf^01^ and tested the behaviour of the transheterozygous F1 offspring. We found that Pdf^01^/Hsp83^08445^ and Pdf^01^/Hsp83^e6A^ transheterozygotes show reduced rhythmicity in DD (Table 2) and decreased phase coherence compared to heterozygous Pdf^01^/+ and Hsp83^08445^/+ or Hsp83^e6A^/ + controls (S2B Fig and S3, S5 and S6 Tables). Importantly, both transheterozygous combinations showed the same (Pdf^01^/Hsp83^e6A^), or more severe (Pdf^01^/Hsp83^08445^) phase dispersal compared to the respective homozygous Hsp83 mutant controls (S2B Fig and S3, S5, and S6 Tables). The results confirm both, a genetic interaction between Hsp83 and Pdf, and that the phenotypic expression depends on the severity of the Hsp83 allele and thereby its effects on Pdf. Interestingly, although heterozygous Pdf^01^ mutant flies are 100% rhythmic in DD, their phase coherence is reduced compared to heterozygous Hsp83 mutants and similar to that of homozygous Hsp83^08445^ mutants (S2B Fig and S3, S5, and S6 Tables), indicating a dominant effect of Pdf^01^ on phase coherence.
Hsp83 mutants affect PERIOD expression rhythms in PDF-negative clock neurons
In DD, high-amplitude PER oscillations in the 5^th^ sLNv and the LNd do not depend on PDF [10]. In fact, in Pdf^01^ mutants PER robustly oscillates in phase in all six LNd, while in wild type, PER cycles in opposite phase in the three CRY^+^ compared to the three CRY^-^ LNd [10]. These effects of PDF-absence are nicely reflected by the expression of the period-luciferase reporter 8.0-luc: This promoter-less PER encoding transgene contains intronic per regulatory sequences, directing PER expression to the three CRY^-^ LNd, the 5^th^-LNv, and the majority of the DN neurons (DN1-DN3) [31,10]. Strikingly, in a Pdf^01^ mutant background, 8.0-luc expression in the DN groups is strongly reduced, while strong expression is observed in all six LNd and the 5^th^ s-LNv [10]. Presumably this explains why in Pdf^01^ the 8.0-luc bioluminescence rhythms are more robust and of higher amplitude compared to the wild type background [10]. Due to the strong effects of Hsp83 mutants on Pdf transcription and PDF accumulation, we analyzed 8.0-luc expression in the Hsp83^08445^ mutant background (Fig 6). To determine bioluminescence expression, 8.0-luc males in wild type, Pdf^01^, or Hsp83^08445^ mutant background were measured for 9 days in a LumiCycle luminometer in DD at 25°C with high temporal resolution (4 min intervals, [32]). Strikingly, we observed a strong increase in the amplitude of 8.0-luc oscillations in Hsp^8308445^ mutant flies, which surpasses the comparatively milder effects of Pdf^01^ (Fig 6). As evident from the raw data plotted in Fig 6A, high-amplitude oscillations continue throughout the experiment in the background of both Pdf^01^ and Hsp83^08445^ mutant flies, consistent with the epistatic effects of Hsp83 on Pdf expression (Fig 5). Similar to Pdf^01^, Hsp83^08445^ also resulted in short-period 8.0-luc oscillations, reducing τ to 23.6 h--1.5 h shorter compared to the wild type background (Fig 6B–6C, [10]). To determine, if Hsp83 depletion has similar effects on spatial 8.0-luc expression as Pdf^01^, we performed anti-PER stainings on brains of per^01^; 8.0-luc; Hsp83^08445^ mutant flies. Indeed, as in Pdf^01^, anti-PER signals (reflecting 8.0-luc expression) were detected in all six LNd and the 5^th^-LNv (Fig 6D–6E and S7 and S8 Tables). Moreover, we observed additional signals in the s-LNv and l-LNv clock neurons, possibly explaining the enhanced expression level and amplitude of bioluminescence rhythms in Hsp83-depleted compared to Pdf^01^ flies. In summary, these results further support that Hsp83 positively regulates Pdf expression, and that mis-regulated PDF levels contribute to the the molecular and behavioral phenotypes of Hsp83 mutations. Because the effects of Hsp83 depletion on spatial and temporal 8.0-luc expression are more severe compared to Pdf^01^, they also indicate that Hsp83 has additional targets within the clock neurons.
Hsp83 depletion affects PERIOD-LUCIFERASE oscillations in subsets of clock neurons.Bioluminescence recordings from 8.0-luc flies in the genetic background indicated. 15 flies were measured for 9 days in DD at 25°C with 4 min time resolution. A) Raw bioluminescence data (counts per second, CPS). B) Detrended data (arbitrary units, a.u.). C) Period estimations using wavelet analysis averaged over 9 days (left, Median indicated by colored horizontal lines) and over time (right) (experimental repeats: 4). See text and Materials and Methods for details. D) Quantification of PER intensity in the different dorsal groups of per01; 8.0-luc flies either in a wild type background or in the background of the Hsp8308445 mutation. E) Representative images of PER (green) and PDF (magenta) staining in per01; 8.0-luc and per01;8.0-luc; Hsp8308445 brains in the different lateral clock neuron groups. Triangle marks the 5th s-LNv. Scale bar: 10 µm.
Discussion
We show here that the chaperon protein HSP83 buffers inter-individual behavioural variability. In contrast to previous studies [25,18], we did not observe significant effects of hypomorphic Hsp83 mutants on the strength of free-running behavioural rhythms, with the exception of Hsp83 depletion in all clock neurons (Table 2). While we have no explanation for this discrepancy, it is possible that differences in the release of CGV in the locally kept Hsp83 mutant strains, accumulated over many generations, contribute to the different results. In favor of this idea is the observation by Goda et al. [25], that rhythmicity was only reduced in Hsp83^e6D^/Hsp83^08445^ mutant males, while females were completely normal, indicating the release of one or several X-chromosomal hypomorphic alleles that compromise rhythmicity. In contrast, we were able to reproduce and refine the overall increased circadian variability. Importantly, we observed a significant decrease in phase coherence, both in Hsp83 mutants and after Hsp83 depletion in all clock neurons, or the PDF expressing subset only. Our results therefore pinpoint the clock neurons as cellular substrate for HSP83 function in buffering behavioural variation. For morphological traits, HSP83 depletion is thought to increase variation via the release of CGV, or through epigenetic mechanisms, although the underlying molecular mechanisms are not understood [13,14,17]. An epigenetic mechanism is usually characterized by a maternal effect phenotypically expressed in the heterozygous offspring [17]. Because we did not observe an increase of behavioural variability in heterozygous Hsp83^08445^/ + flies resulting from a cross of homozygous Hsp83^08445^/Hsp83^08445^ females to wild type (iso31) males (Fig 2 and Tables 1 and 2), we favour the idea that Hsp83 depletion releases CGV.
We show that Hsp83 genetically interacts with Pdf (S2A Fig), a neuropeptide encoding gene, required for normal locomotor behaviour of fruit flies [8]. Hsp83 depletion leads to reduced Pdf expression at the transcriptional level, presumably resulting in slower replenishment of PDF levels after PDF release in the s-LNv terminals in the dorsal brain (Figs 5, S3A, and S3D). The effects on PDF expression supply a reasonable explanation for the observed variation, and previous studies indeed showed that Pdf^01^ mutants show increased variability of their free-running period and in the ability to maintain rhythmicity in DD [8]. However, the behavioural phenotypes of Hsp83 depletion do not match those of Pdf^01^ mutants. Flies lacking PDF in the s-LNv display short-period behavioural rhythms in DD, which after a few days deteriorate to arrhythmicity. Moreover, PDF is required for morning anticipation (i.e., the behavioural activity increase before the lights come on in the morning), as well as for proper timing of the evening activity peak during LD [8,33]. We think that altered PDF-expression in Hsp83 mutants, as opposed to lack of PDF in Pdf^01^ mutants are the reason for this discrepancy. PDF exerts its function after release from dense core vesicles and binding to its receptor (PDFR), which is expressed in non-PDF clock neurons, including the three CRY-positive LNd and the 5^th^ s-LNv, the DN1a, six Cry-positive DN1p, a few DN3, and the PDF-expressing s-LNv themselves [34]. While PDF is required for synchronous and high-amplitude oscillations in most of the clock neurons (e.g., the s-LNv), other groups, like the six LNd and the 5^th^ s-LNv exhibit increased synchrony of clock gene expression and cycle with high-amplitude and short period in the absence of PDF [9,10]. These effects of Pdf^01^ on clock gene expression are accurately and conveniently reported by the 8.0-luc period-luciferase reporter whose expression is largely restricted to the PDF-negative PDFR clock neurons and the Cry-negative LNd [31,10] (Fig 6). We observed very similar effects of the Hsp83^08445^ mutant on 8.0-luc expression, including high-amplitude and short-period rhythms, further supporting HSP83 influence on PDF-signaling (Fig 6).
Our results are reminiscent of a recent study showing that transcriptional changes in Pdf expression contribute to circadian variability between different Drosophila species [19]. The equatorial species D. sechellia has lost the ability to adjust its activity to long photoperiods, which is causally linked to reduced Pdf expression caused by altered 5’-regulatory sequences compared to D. melanogaster [19]. Regulation of Pdf expression therefore appears to be an endpoint of various factors that influence circadian variation at the species (e.g., evolution of regulatory sequences) and individual (regulation by HSP83) level. Taken together our results provide compelling evidence for a role of HSP83 in regulating PDF expression.
However, based on the differences between Pdf^01^ and Hsp83 mutant phenotypes, and the more substantial behavioural effects of HSP83 depletion in all clock neurons compared to PDF neurons only (Table 1 and Figs 3 and 4), it seems likely that HSP83 has additional targets, either directly within the core molecular clock mechanism, or affecting other circadian neuropeptides. For example, in the mammalian clock system HSP90 stabilizes the CYC homologue BMAL1, and fly clock neurons express at least eleven different neuropeptides in addition to PDF [2,35]. Identifying PDF-misregulation as consequence of Hsp83 depletion and contribution to behavioural variability, our findings set the stage for addressing the underlying molecular mechanism of HSP83 behavioural capacitor function. Our results provide evidence for a role of HSP83 in the transcriptional regulation of Pdf. It is known that Pdf transcription is indirectly regulated by the transcription factor CLK, which activates the transcription of the genes stripe (sr) and Hormone receptor-like in 38 (Hr38) within the s-LNv, both of which encode Zinc Finger transcription factors [29]. Because both, SR and HR38 repress Pdf transcription [29] it is unlikely that these transcription factors (and CLK) are targets of HSP83 chaperon function. Transcription factors that positively regulate Pdf transcription have not been identified yet, but are possibly regulated by Hsp83 to increase behavioural variability under environmental stressful conditions.
Materials and methods
Fly strains and maintenance
Flies were raised on fly food containing 0.7% agar, 1.0% soya flour, 8.0% polenta/maize, 1.8% yeast, 8.0% malt extract, 4.0% molasses, 0.8% propionic acid, 2.3% nipagin) in 12 h:12 h LD cycle at 60% relative humidity at 25°C. All Hsp83 variants were obtained from the BDSC and w iso31 flies were used as controls [36]. Hsp83^e6D^ (BDSC 5696) is an antimorphic, homozygous lethal EMS-induced allele with a single amino-acid replacement (E317K) [37]. Hsp83^e6A^ (BDSC 36576) is a strong hypomorphic, homozygous lethal EMS-induced allele with the single amino-acid replacement S592F [37]. Hsp83^j5c2^ (BDSC 12064) is an amorphic homozygous lethal allele, induced by the insertion of the P{lacW} transposable element into the first non-coding Hsp83 exon, which is part of both Hsp83-RA and Hsp83-RB transcripts. Hsp83^08445^ (BDSC 11797) is a hypomorphic allele, induced by the insertion of the P{PZ} transposable element into the first non-coding exon of the Hsp83-RB transcript. Hsp83^08445^ is homozygous viable (presumably because the Hsp83-RA is not, or only mildly affected), and produces viable offspring when combined with the other three Hsp83 alleles (Table 1). The Hsp83sgRNA line (BDSC 80843) expresses Hsp83 specific sgRNA ubiquitously for mutagenesis of Hsp83 under UAS-Cas9 (BDSC 58985) control. tim-gal4:27 [38], Clk856-Gal4 [27] and Pdf-Gal4 [8], were used to drive Cas9 expression in all clock cells, clock neurons, or the PDF expressing LNv subset, respectively. Pdf-Red is a direct fusion of a 0.6 kb Pdf promoter region to mRFP1 [28]. UAS-Hsp83.Z (BDSC 58469) was used for rescue, and CCAP-Gal4 (BDSC 25685) and UAS-myr-mRFP1 (BDSC 7118) for stability experiments, respectively. 8*.0-luc:9* flies contain a promoter-less period-luciferase transgene on chromosome 2, encoding a PER-LUC fusion protein expressed in subsets of the PDF negative clock neurons DN1–3, LNd, and 5^th^ s-LNv [31,10]. 8.0-luc:9 was combined with the Pdf^01^ [10] and Hsp83^08445^ allele using standard genetic crosses.
Behavior
1 to 5 days old male flies were loaded individually into locomotor glass tubes with food (4% sucrose and 2% agar) at one end and a cotton cup at the other. The glass tubes were placed into DAM2 monitors of the Drosophila Activity Monitor (DAM) system (TriKinetics), to measure locomotor activity of the flies. DAM monitors were housed in programmable environmental incubators (Percival Scientific, USA). Flies were synchronized to 12 h: 12h light-dark (LD) cycle for at least 3 days and subsequently kept in constant darkness (DD) for at least 5 more days at 25°C. As controls for each genotype, mutants, Gal4 driver, and UAS lines were crossed w iso31, and heterozygous offspring was analyzed.
Behavioral analysis
We analyzed activity in 3 days in LD and 5 days in DD. Experimental repeats are listed under each figure. Period, percentage of rhythmic flies, rhythmic strength (RS) and phase analysis were computed using Flytoolbox in Matlab [39]. Phase was quantified over the second (DD2) and third (DD3) day in constant darkness. To prevent the algorithm from identifying artifactual peaks, arrhythmic flies were excluded from the phase analysis.
Actograms showed in the figures were computed using Rethomics in R [40], while phase graphs were created using a custom-made script in R (v4.1.2; R Core Team 2021). Entrainment index (EI) was computed in R (v4.1.2; R Core Team 2021), based on established protocols and correcting for the startle responses to light [20]. To calculate if flies concentrate most of their activity in anticipation of the light-dark transitions in the morning and evening, we summed up the activity during the 6 h before the respective transition, divided by the total activity 6 h before and the 6 h after each transition. In order to quantify the percentage of entrained individuals, we summed the entrained individuals who concentrate more than three quarters of their activity in the 6 hours preceding the light transitions, therefore who had an EI higher than 0.75.
To gain a better understanding of the inter-individual variability observed in our experiments, we used hidden Markov models (HMMs) to infer activity states from light barrier crossings [21]. We modelled data from the mutants and cell-specific knock-out separately, using 2-state HMMs to distinguish active and inactive behaviour. The (negative binomial) state-dependent distributions of locomotor activity are estimated separately for LD and DD, and were determined based on the controls only. Subsequently, we fixed the distributional parameters for fitting HMMs to the other genotypes to ensure comparability of the activity states in LD and DD between genotypes. To investigate temporal activity patterns, we modelled the state-switching probabilities as a function of time of day for each genotype, including random intercepts to account for individual differences. For reasons of parsimony, we restricted the random effects variances to be the same for the two state-switching probabilities. All models were implemented using functions from the R package LaMa (Koslik, 2025) [41]. Based on the fitted HMMs, we derived the model-implied percentage of time spent in each state as a function of the time of day [42] and calculated its variance between individuals at the morning and evening peaks (determined based on the control genotype iso31) for each genotype in LD and DD, respectively. Graphs were plotted in RStudio and then re-arranged, using the MacOS-based software Vectornator (Version 4.11.5, 2022, Linearity GmbH), also known as Linear Curve.
Bioluminescence assay
D-luciferin potassium salt (Biosynth) was mixed with standard fly food to a final concentration of 15 mM in Drosophila culture plates (Actimetrics). Luminescence of 15 males per plate was measured every 4 min for 7–8 days in DD at 25°C with a the LumiCycle 32 Color (Actimetrics). Actimetrics analysis software was used to normalize the exponential decay, data were exported into.csv files and custom python code was used to organize luminescence data into 30-minute bins (LABLv9.py; www.top-lab.org/downloads), and to quantify periods of oscillations using a Morlet wavelet fit (waveletsv4.py; www.top-lab.org/downloads). Data were plotted using Graphpad Prism 10. For details see [32]. and then re-arranged, using the MacOS-based software Vectornator (Version 4.11.5, 2022, Linearity GmbH), also known as Linear Curve.
Immunohistochemistry
For the staining of the cell-specific Hsp83 knock-out, we first entrained flies for 5 days in 12 hours light 12 hours dark (LD). After that, we fixed flies at ZT02and ZT14. Flies were fixed in 4% PFA for 2.5 h at room temperature (RT). After fixation, the samples were washed 6 times with 0.1 M phosphate buffer (pH 7.4) with 0.1% Triton X-100 (PBS-T) at RT. Brains were dissected in PBS, were then blocked with 5% goat serum in 0.1% PBS-T for 2 h at RT and stained with mouse anti-PDF C7 (DSHB, 1:500), rabbit anti-RFP (Merck, 1:500) or guinea pig anti-VRI (1:2000) [43] in 5% goat serum and 0.5% PBST for at least 48 h at 4⁰C. After washing 3 times in PBS-T, the samples were incubated at 4°C overnight with goat anti-mouse AlexaFluor 488 nm, anti-rabbit Alexa Fluor 550 or anti-guinea pig Alexa Fluor 568 (1:600) in PBS-T. Brains were washed 3 times in PBS-T before being mounted in Vectashield. Brains of 1–2 days old Pdf-Red flies were dissected in a physiological medium [44], mounted in ProLong Gold (Thermo Fisher Scientific), and imaged directly. PDF and Pdf-Red images were taken using a Leica SP8 confocal microscope, keeping the same settings for each experiment, all other images were generated using a Leica Stellaris microscope.
Immunostaining quantifications
For quantification of RFP and PDF intensity, pixel intensity of mean and background staining in each neuronal group was measured by ImageJ, FIJI [45]. Soma intensity was quantified drawing one vector in the center of the neuron and then we used the same vector to measure the corresponding background intensity. For PDF projections, we extracted a stack of the 10 Z slides where the s-LNv dorsal projections are showed at their maximum intensity. Then, we drew a vector at the center of the projection and used the same vector to measure the background intensity. Data were normalized by dividing the values by the length of the vector and consequently by subtracting the background signal. Quantification of PER (Fig 6D) and VRI (S3E Fig) intensity was performed as described in [46]. In brief, for each cell three measurements were taken, as well as 3 measurements of the background for the corresponding slice for background subtraction.
Statistical analysis
Descriptive statistics and the statistical tests used are listed in each figure and are summarized in S3 and S5–S7 Tables. Circular statistics for phase analysis (Figs 2 and 4) were computed in R (R Core Team, 2021), using the package circular [47]. As the goal of our study was to determine differences in variability between groups, we reported the median and the mu (µ) to understand population the central value, and the circular standard deviation (SD) and the concentration kappa (k) to show how spread the data are around the central value (S3 Table) [48]. Topics in Circular Statistics, Section 5.3, World Scientific Press, Singapore.). Because both the µ and k values are parameters for the Von Mises distribution, we applied the Watson’s goodness of fit test to determine whether the data met those assumptions (S5 Table) [48]. As all genotypes meet the Von Mises assumptions, we applied a pairwise equal kappa (κ) test and we adjusted the p-value using Bonferroni method (S6 Table) [48]. For staining intensity, we used estimation statistics [49] (S7 Table).
Supporting information
S1 FigHeterozygous Hsp83 loss of function does not affect behavior in LD and DD.A) Population Activity (top) and HMM-implied percentage of time spent in the active state (bottom) of Hsp83 heterozygous (Hsp83 mutant/+) flies in LD. B) Population Activity (top) and HMM-implied percentage of time spent in the active state (bottom) of Hsp83 heterozygous (Hsp83 mutant/+) flies in DD (repeats: 4). See also Tables 1, 2, S2 and S3.(DOCX)
S2 FigClock neuron-specific rescue of Hsp83 mutants and genetic interaction with *Pdf.*A) Circular phase plot comparing Hsp83^08445^ mutant flies with or without expression of UAS-Hsp83 in all clock neurons (repeats: 4–5). B) Circular phase plots of trans-heterozygous Pdf^01^/Hsp83^08445^ (left) and Pdf^01^/Hsp83^e6A^ (right) mutant flied and their respective controls./+) (repeats: 2). Arrhythmic flies were excluded from the peak analysis. Colored lines represent the median phase of each group, while the length of the vector represents the circular standard deviation of the corresponding group (1/circular SD). Watson’s goodness of fit test and pairwise equal kappa (κ) test **** p < .0001, *** p < .001, ** p < .01, * p < .05, ns p > .05.(DOCX)
S3 FigHsp83 depletion affects PDF mRNA and protein levels.A) Quantification of the RFP levels in Pdf-Red; + / + compared to Pdf-red; Hsp83^08445^/Hsp83^08445^ flies for the s-LNv, dorsal projection and l-LNv, using Estimation Statistics. B) Quantification of RFP levels in the cell bodies (left) and projections (right) of CCAP neurons (CCAP-Gal4 > UAS-myr-mRFP1) either in a wild type background or with the Hsp83^08445^ mutation C) RFP expression in the CCAP+ neurons using the CCAP-Gal4. Scale bar: 20μm. D) Representative images of anti-PDF staining of the s-LNv, dorsal projections, and l-LNv in Hsp83 knock out (Clk856-Gal4 > UAS-Cas9, hsp83 sgRNA) and control flies (hsp83 sgRNA/+). Scale bar: 20μm. E) Left: Representative images of anti-VRI staining in the l-LNv and s-LNv of iso31 and Hsp83^08445^/Hsp83^08445^ flies. Scale bar: 20μm. Right: Quantification of the intensity of VRI staining in the s-LNv and l-LNv in 8 hemispheres, using Estimation Statistics.(DOCX)
S1 TableViability of Hsp83 allelic combinations.(DOCX)
S2 TableRhythmic power of Hsp83 allelic combinations in LD.(DOCX)
S3 TableCircular Phase plot and HMM analysis reveal increased variability of evening activity phase after HSP83 depletion in constant darkness (DD).(DOCX)
S4 TableNumber of PDF projections, s-LNvs and l-LNvs neurons per hemisphere in cell-specific Hsp83 knock-out and parental control.(DOCX)
S5 TableWatson’s goodness of fit test to determine Von Mises distribution.(DOCX)
S6 TablePairwise equal kappa (κ) test with adjusted p-value using Bonferroni method.(DOCX)
S7 TableEstimation statistics signal intensity.(DOCX)
S8 TableNumber of PER^+^ lateral neurons per hemisphere in per^01^; 8.0-luc brains.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1van der Vinne V, Tachinardi P, Riede SJ, Akkerman J, Scheepe J, Daan S, et al. Maximising survival by shifting the daily timing of activity. Ecol Lett. 2019;22(12):2097–102. doi: 10.1111/ele.13404 31617283 PMC 6899458 · doi ↗ · pubmed ↗
- 2Reinhard N, Fukuda A, Manoli G, Derksen E, Saito A, Möller G, et al. Synaptic connectome of the Drosophila circadian clock. Nat Commun. 2024;15(1):10392. doi: 10.1038/s 41467-024-54694-0 39638801 PMC 11621569 · doi ↗ · pubmed ↗
- 3Helfrich-Förster C, Reinhard N. Mutual coupling of neurons in the circadian master clock: What we can learn from fruit flies. Neurobiol Sleep Circadian Rhythms. 2025;18:100112. doi: 10.1016/j.nbscr.2025.100112 39906412 PMC 11791320 · doi ↗ · pubmed ↗
- 4Tataroglu O, Emery P. The molecular ticks of the Drosophila circadian clock. Curr Opin Insect Sci. 2015;7:51–7. doi: 10.1016/j.cois.2015.01.002 26120561 PMC 4480617 · doi ↗ · pubmed ↗
- 5Benito J, Zheng H, Hardin PE. PDP 1epsilon functions downstream of the circadian oscillator to mediate behavioral rhythms. J Neurosci. 2007;27(10):2539–47. doi: 10.1523/JNEUROSCI.4870-06.2007 17344391 PMC 1828026 · doi ↗ · pubmed ↗
- 6Gunawardhana KL, Hardin PE. VRILLE controls PDF neuropeptide accumulation and arborization rhythms in small ventrolateral neurons to drive rhythmic behavior in Drosophila. Curr Biol. 2017;27(22):3442–3453.e 4. doi: 10.1016/j.cub.2017.10.010 29103936 · doi ↗ · pubmed ↗
- 7Zheng X, Koh K, Sowcik M, Smith CJ, Chen D, Wu MN, et al. An isoform-specific mutant reveals a role of PDP 1 epsilon in the circadian oscillator. J Neurosci. 2009;29(35):10920–7. doi: 10.1523/JNEUROSCI.2133-09.2009 19726650 PMC 2757269 · doi ↗ · pubmed ↗
- 8Renn SC, Park JH, Rosbash M, Hall JC, Taghert PH. A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell. 1999;99(7):791–802. doi: 10.1016/s 0092-8674(00)81676-1 10619432 · doi ↗ · pubmed ↗