Evolving threats: Leveraging C. elegans to decode the virulence profiles of highly related environmental Salmonella Newport isolates
Christina M. Ferreira, Mirae Choe, Bella Wayhs, Julie A. Haendiges, Robert Literman, Jianghong Meng, Arjuman Ghazi, Rebecca L. Bell

TL;DR
This study uses C. elegans to compare the virulence of different Salmonella Newport isolates, revealing that some environmental strains are more harmful than others.
Contribution
The study identifies specific environmental S. Newport isolates with higher virulence using a C. elegans model and genomic analysis.
Findings
Newport-1015 and Newport-61 isolates showed higher virulence in C. elegans survival assays.
The isolates differ by a 1.7 Mb genomic inversion, suggesting genetic factors influence virulence.
Environmental S. Newport strains can be as virulent as those from foodborne outbreaks.
Abstract
Salmonella enterica subspecies enterica, particularly serovar Newport, remains a leading cause of foodborne illnesses in the United States, implicated in numerous outbreaks associated with a diverse array of food products. This study systematically investigates the virulence of five distinct S. Newport isolates, characterized by varying patterns of pulse-field gel electrophoresis (PFGE) molecular-diagnostic subtyping, using the nematode Caenorhabditis elegans as a host model organism. We conducted viability assays on C. elegans to evaluate how these isolates affect nematode survival. The selected bacterial strains, chosen for their historical significance in foodborne outbreaks yet isolated form environmental sources, were previously sequenced to provide a comprehensive genomic framework. A notable focus of our research was on the nearly genetically identical PFGE types Newport-61 and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
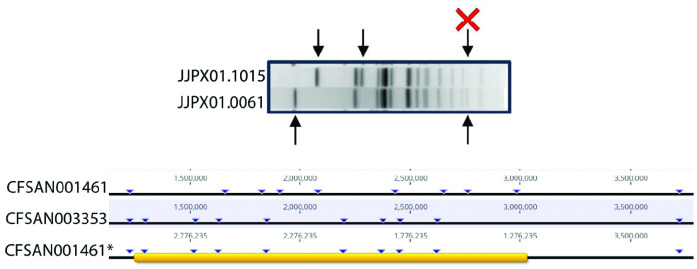 Fig 1
Fig 1 Fig 2
Fig 2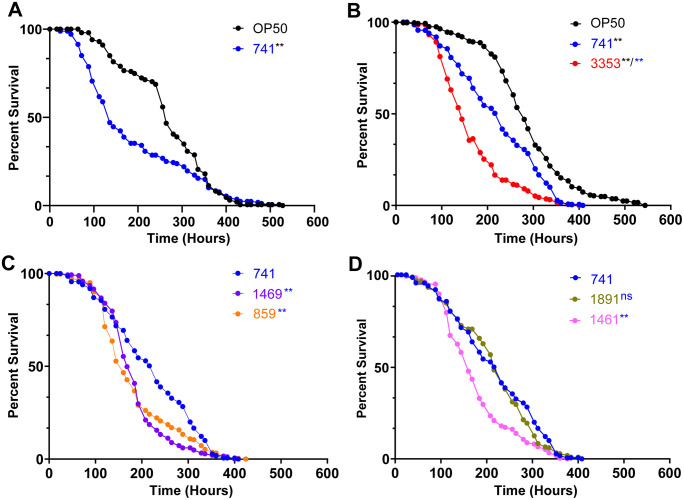 Fig 3
Fig 3- —http://dx.doi.org/10.13039/100000002National Institutes of Health
- —http://dx.doi.org/10.13039/100000002National Institutes of Health
- —http://dx.doi.org/10.13039/100000002National Institutes of Health
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsListeria monocytogenes in Food Safety · Salmonella and Campylobacter epidemiology · Vibrio bacteria research studies
Introduction
Salmonella enterica subspecies enterica is the leading cause of microbial foodborne illnesses in the United States, resulting in over 1 million infections and more than 400 deaths each year [1,2]. Among its serovars, S. enterica serovar Newport (S. Newport) ranks as the second most common culture-confirmed serovar associated with foodborne outbreaks in the country [3]. S. Newport has a broad host range, implicated in illnesses linked to a variety of foods, including tomatoes, cucumbers, papaya, onions, and ground beef [4–9]. From 2000 to 2020, there were 227 foodborne salmonellosis outbreaks attributed to S. Newport, with six of those being recurrent outbreaks over a 12-year period connected to tomatoes from the Virginia Eastern Shore (VES) [4,10,11]. These tomato outbreaks were linked to a clonal strain of S. Newport, identified through PFGE as “Pattern 61” (XbaI JJPX01.0061, Newport-61). This clone is now categorized by the CDC under REPJJP03, which also includes S. Newport Pattern 1015 (XbaI JJPX01.1015, Newport-1015).
The nematode Caenorhabditis elegans has been established as a valuable model organism for studying the virulence mechanisms of human pathogens. A short lifespan (2–3 weeks) along with remarkable amenability to molecular, genetic and physiological interventions has led to discovery of fundamental innate immune mechanisms relevant to human infectious diseases [12,13]. Despite the absence of dedicated immune cells, worms deploy highly conserved immune response pathways such as the p38 MAPK response and TGF-beta pathways that have been studied extensively [14,15]. While C. elegans lacks the nuances of innate immunity present in mammalian immune systems, similarities in detecting infections and their resulting immune responses renders this model organism invaluable for studying pathogens that higher eukaryotes and humans are susceptible to as well as anti-microbial drug discovery [16,17]. Numerous pathogens, including S. enterica, colonize the host worm through their intestine following consumption and hence it is especially suitable for modeling immune mechanisms elicited by food-borne pathogens. Previously, worms exposed to S. enterica serovar Typhimurium (Typhimurium) strains were shown to have a significantly shortened lifespan with visible signs of infection in the intestinal tract. Notably, Salmonella strains that exhibit reduced virulence in mammals were shown to have similarly attenuated impact on the lifespan of C. elegans [18,19]. This indicates that bacterial genes important for pathogenic potency in vertebrate hosts are likely similarly required during C. elegans infection, highlighting its value to assess pathogenic virulence and host-pathogen interactions.
Newport-61 and Newport-1015 have been isolated from the surface water and sediment of the VES, with a seasonal recurrence of clonal isolates [20,21]. However, there are few documented cases, prior to the retirement of CDC PFGE PulseNet, of Newport-1015 causing human infection, suggesting overall less virulence capacity. In this study, we used C. elegans to evaluate the virulence of five S. Newport environmental isolates, each representing different PFGE patterns of historical significance related to foods. These isolates are closely related, with Newport-1015 and Newport-61 being particularly noteworthy, as they are nearly genetically identical except for a 1.7 Mb inversion [22].
Materials and methods
Bacterial isolates
Strains used in the C. elegans experiment included Escherichia coli OP50 (negative control) and six Salmonella enterica strains found in Table 1. The S. Typhimurium isolate (CFSAN000741) was used as the positive control for the C. elegans assays, and the S. Newport isolates (CFSAN000859, CFSAN001461, CFSAN001891, CFSAN003353, and CFSAN001469) were previously reported [22]. All strains were maintained as frozen stocks in Brain Heart Infusion Broth (BHIB, BD Difco) with 50% glycerol (v/v), and experiments were conducted with freshly grown cultures, streaked from frozen stock cultures onto Luria Bertani agar (LBA, BD Difco) and incubated at 35°C ± 2°C for 22 ± 2 hours [23]. Single colonies from these plates were inoculated into 3mL of Luria Bertani broth (LB, BD Difco) and incubated statically at 35°C ± 2°C for 22 ± 2 hours.
Table 1: Salmonella strains used for bioinformatic analysis and C. elegans assays.
Salmonella WGS comparative analysis
Strains used in this study were previously sequenced to provide complete, reference quality genomes using the PacBio [24]. The assemblies were uploaded to NCBI and annotated using the NCBI Prokaryotic Genome Annotation Pipeline (PGAP, v. 6.10) [25]. Roary (v. 3.13.0) was used to generate a pangenome alignment and to identify genes that were unique to the different PFGE patterns [26]. Visualization of the output from Roary were created using Phandango (v. 1.3.1) [27]. AMRFinderPlus (v. 4.0.3, with default settings and “plus genes”) and Phastest (v. 3.0 with deep annotation) were used to identify antimicrobial resistance (AMR), virulence, and phage genes [28–30]. BLASTN, hosted by NCBI with default settings, was used to identify the percent identity of the strains [31]. Geneious Prime (v 2025.1.2) was used for in silico restriction digestion of the strains with the enzyme XbaI to identify the difference in PFGE pattern comparison of JJPX01.0061 and JJPX01.1015.
Caenorhabditis elegans survival assay
C. elegans survival experiments were conducted at 20°C with the wild-type strain, N2, using standard techniques. Worms were maintained on nematode growth medium (NGM), a standard, nutrient-rich solid media used for C. elegans culture as described previously [32,33]. Briefly, NGM plates were seeded with 70 μL of the worms’ normal laboratory diet of E. coli strain OP50 (OP50) or one of the six Salmonella pathogenic strains. OP50 plates were allowed to dry for 24 hours before adding worms. Salmonella-seeded plates were used after 5 hours of drying; hence fresh Salmonella plates were prepared every 24 hours.
For the survival assays, healthy, gravid young adult worms were transferred to fresh OP50-seeded NGM plates, allowed to lay eggs for 1–2 days and removed. The eggs were reared at 20°C and age-matched L4-stage, pre-adult larvae were selected for the experiment and transferred to NGM plates seeded with either the OP50 control bacteria or various Salmonella strains. Thirty L4-stage worms were transferred to each plate, with a total of 150 worms tested per bacterial strain. Worm survival was scored twice daily by gently touching the head, tail, or midsection with a platinum wire pick; worms were considered dead if they failed to respond. Worms were transferred to fresh, corresponding plates every 24 hours. Survival monitoring continued until all original worms died, using 12-hour/12-hour or 8-hour/16-hour scoring intervals. Worms that exploded, bagged, crawled off the plate, or were otherwise unaccounted for were censored from analysis. All survival assays were conducted twice in two biological replicates. Survival statistics were plotted using the Kaplan–Meier method. Statistics were calculated using the nonparametric log-rank Mantel−Cox method on the OASIS2 platform (v.2.4.2) and subjected to multiplicity Bonferroni correction [34].
Results and discussion
Genomic analysis reveals a large-scale inversion distinguishing Newport-61 and Newport-1015 lineages
The strains presented in this study are all known to cause human infections to varying degrees and have been identified across the United States. Gene content analysis shows that these isolates demonstrate over 98% genetic identity to one another, with Newport-61 being more than 99% identical to Newport-1015 (Fig 1). Analysis of genetic content related to virulence, AMR or prophage presence did not show any differences among the S. Newport isolates in this study (Table 2 and S1 Table). It was previously reported that two strains, CFSAN001891 and CFSAN001461 (both JJPX01.1015), have a 1.7 Mbp inversion relative to the chromosome of CFSAN003353 (JJPX01.0061) [22]. As can be seen in Fig 1, the 1.7 Mbp inversion accounts for the difference in PFGE patterns between JJPX01.0061 and JJPX01.1015 as the location of the XbaI restriction enzyme site is altered (Fig 2, indicated by “X”).
Table 2: AMRFinder Plus identified virulence and antimicrobial resistance genes for Salmonella isolates in this study.
Phylogeny showing gene presence/absence of key Salmonella Newport isolates.The phylogenetic tree shows the relatedness of the S. Newport isolates to one another. The blue colored area represents the presence/absence of genes across the entirety of the genomes. The sequence types represented include 118 (magenta), 350 (yellow) and 5 (dark blue).
Inversion confirmation of CFSAN001461 and CFSAN1891 using in silico PFGE analysis.PFGE XbaI pattern of JJPX01.1015 compared to JJPX01.0061 (top), where the arrows highlight the differing bands in the PFGE patterns and the red “X” denotes the band that is not present in the 1015. The inversion identified in JJPX01.1015 accounts for the differences in the PFGE patterns due to changes in cut site locations within the genome. The locations of these cut site changes (bottom) due to the directionality of the inversion – by manually rotating the 1.7Mbp JJPX01.1015 inversion at the Gifsy-1 sites (denoted by CFSAN001461) and subsequently performing in silico digestion with XbaI, the cut sites revert to the same fragment sizes reflected in JJPX01.0061 (CFSAN003353) confirming the accuracy of inversion in the closed genome.*
Despite having short-read Illumina sequences available at NCBI, this inversion went undetected in these strains. To our knowledge, this inversion is the largest ever identified in Salmonella, surpassing the next largest known inversion of 1.62 Mbp found in S. Typhimurium [35]. While it has been noted that serovar Typhimurium tends to invert portions of its genome around ribosomal RNA (rRNA) regions, those segments are comparatively small, measuring only 70−80 kb, unlike the inversion observed in these isolates [36,37]. Furthermore, the flanking regions of this inversion do not appear to contain rRNA; instead, they are associated with a Gifsy-1 phage, which appears to have become grounded due to its inability to excise as evidenced by the absence of the anti-repressor gene in the prophage (S1 Table) [38]. With the S. Typhimurium inversion, the rRNA remain functional and the expression of the genes within the inversion remain intact, we did not investigate whether the inversion in Newport-1015 modified gene regulation in some way. Gammaproteobacteria have been shown to invert small sections of specific genes to phase variate in response to environment specific signals, and these variations are able to regulate both overall expression and modification of protein sequence [39]. Additionally, while these isolates were all collected from environmental sources, it is possible that the genetic inversion coupled with both the high sequence identity and presence of the flanking Gifsy-1 phage may be related to host-specific adaptations [40,41].
Closely related Salmonella strains exhibit distinct pathogenicity dynamics in a live C. elegans host
The Salmonella strains tested shortened worm survival as compared to animals on E. coli OP50. Importantly, these strains showed differential impacts on survival rates with significantly different time to death (TD_50_) for each, with trends that were similar between two independent trials (S2 Table). As a positive control, we exposed C. elegans to S. Typhimurium CFSAN000741 and found that, as previously reported, it shortens C. elegans mean survival by 15%−30% in independent trials (Fig 3A and Table 3) [18,19]. Upon testing the 6 strains used in this study, we found that the strongest, consistent pathogenic impact was seen upon exposure to CFSAN003353, which caused a significantly greater lifespan reduction as compared to S. Typhimurium (53% v 15%) (Fig 3B and Table 3). CFSAN001469 and CFSAN00859 also caused greater survival diminution compared to CFSAN000741, but their impacts were comparatively modest across trials (Table 3).
Table 3: Impact of Salmonella strains on C. elegans Survival.
Impact of Salmonella strains on survival of C. elegans.A: S. Typhimurium (741, blue) shortens the survival of C. elegans adults compared to the normal diet of OP50 (black). B: C. elegans lifespan is shortened significantly more upon infection with CFSAN003353 (3353, red) compared to 741. C: CFSAN001469 (1469, purple) and CFSAN00859 (859, orange) caused greater reduction in worm lifespan compared to 741. D: Despite exhibiting the same PFGE pattern with the preserved inversion, strains CFSAN001461 (1461, pink) and CFSAN001891 (1891, olive) shortened worm lifespan to different degrees. 1461 caused significantly greater reduction than the control 741, whereas 1891 had the same magnitude of impact as 741. Data shown is combined from two independent trials (Table 2) and represents mean survival of the population in hours (m) ± SEM. P < 0.0001 (**). ns = not statistically significant. Colors of asterisks indicate the strain being used for comparison.
Strikingly, we found that two of the strains, CFSAN001891 and CFSAN001461, which contain the large inversion and are > 99% identical to each other, exhibited distinctly different TD_50_s. In addition to the 15−30% shortening induced by CFSAN000741, CFSAN001461 exposure aggravated pathogen-driven survival diminution by an additional 25–30%, from approximately 236 hours to 150 hours, whereas CFSAN001891 had the same effect as CFSAN000741 (Figs 3C and 3D and Table 2). The underlying basis for the difference in virulence exhibited by the two strains, CFSAN001891 and CFSAN001461, is unclear from this study. However, these differences likely reflect variation in host–pathogen interactions driven by pathogen-specific features, host factors, or a combination of both, rather than a contradiction of their shared PFGE pattern or inversion status. In C. elegans, as in other species, virulence is determined not only by bacterial identity but also by how pathogen-associated features engage host immune pathways. Subtle differences in bacterial surface structures or secreted factors could differentially activate or evade host defense mechanisms, resulting in divergent survival outcomes even among closely related strains. Conversely, host-intrinsic factors may also contribute to strain-specific responses, including genetic variation that alters how the host perceives or responds to distinct pathogen strains. Consistent with this idea, strain-specific host responses to highly related pathogens such as Enterococcus faecalis and Bacillus thuringiensis (Bt) have been previously reported [42,43]. Notably, transcription factors AP-1 and ELT-2 have been implicated in mediating shared versus strain-specific host responses to Bt, respectively [43]. Future studies, such as tandem host–pathogen RNA sequencing, will be essential to elucidate the molecular mechanisms underlying the virulence differences observed here.
Conclusion
The C. elegans model has proven to be an invaluable tool for assessing a phenotype that might otherwise have gone unnoticed. Together, the experiments presented here underscored the feasibility of using C. elegans to measure the virulence capacity of environmental Salmonella strains as well as nuanced pathogenicity differences likely resulting from genomic events and their downstream host interactions. Given that there were fewer than ten confirmed illnesses linked to Newport-1015 (<10 from 2000−2018), the virulence phenotype discovered in this study is surprising and suggests that this genomovar may not be preferentially adapted to human hosts though sharing high sequence identity with strains known to have caused large illness outbreaks. The isolates studied here offer a unique opportunity to elucidate the underlying regulatory mechanisms that may contribute to not only the differences in virulence among Salmonella serovars but also host adaptive genetic markers. Moreover, due to the highly conserved nature of the genomes, distinguishing differences between the two genomovars (Newport-61 vs. Newport-1015) can currently only be achieved using PFGE or long-read sequencing. Comparative genomics alone is insufficient in drawing conclusions about the virulence potential of closely related strains of Salmonella. For this reason, expression studies will be required to identify the mechanisms underlying these phenotypes, along with genome-wide screening to understand the roles that specific genes play in the infection process.
Supporting information
S1 TableProphages and prophage-like elements in the Salmonella strains from these experiments.(XLSX)
S2 TableImpact of Salmonella strains on C. elegans Survival (Independent Trials).(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dewey-Mattia D, Kisselburgh H, Manikonda K, Silver R, Subramhanya S, Sundararaman P, et al. Surveillance for foodborne disease outbreaks–United States, 2016: annual report; 2018.10.15585/mmwr.ss 6710 a 1PMC 606196230048426 · doi ↗ · pubmed ↗
- 2Bishop R, Erdman M, Fields P, Fullerton K, Jackson K, Mahon B. National enteric disease surveillance: Salmonella surveillance overview. Georgia, USA: National Center for Emerging and Zoonotic Infectious Diseases; 2011.
- 3CDC Cf D Ca P. BEAM (Bacteria, Enterics, Ameba, and Mycotics) Dashboard Atlanta, Georgia: US Department of Health and Human Services; 2025. Available from: www.cdc.gov/ncezid/dfwed/BEAM-dashboard.html
- 4Greene SK, Daly ER, Talbot EA, Demma LJ, Holzbauer S, Patel NJ, et al. Recurrent multistate outbreak of Salmonella Newport associated with tomatoes from contaminated fields, 2005. Epidemiol Infect. 2008;136(2):157–65. doi: 10.1017/S 095026880700859 X 17475091 PMC 2870807 · doi ↗ · pubmed ↗
- 5Hernández-Reyes C, Schikora A. Salmonella, a cross-kingdom pathogen infecting humans and plants. FEMS Microbiol Lett. 2013;343(1):1–7. doi: 10.1111/1574-6968.12127 23488473 · doi ↗ · pubmed ↗
- 6CDC Cf D Ca P. National enteric disease surveillance: Salmonella annual report, 2016; 2016.
- 7Angelo KM, Chu A, Anand M, Nguyen T-A, Bottichio L, Wise M, et al. Outbreak of Salmonella Newport infections linked to cucumbers--United States, 2014. MMWR Morb Mortal Wkly Rep. 2015;64(6):144–7. 25695319 PMC 4584703 · pubmed ↗
- 8Whitney BM, Mc Clure M, Hassan R, Pomeroy M, Seelman SL, Singleton LN, et al. A series of papaya-associated Salmonella illness outbreak investigations in 2017 and 2019: a focus on traceback, laboratory, and collaborative efforts. J Food Prot. 2021;84(11):2002–19. doi: 10.4315/JFP-21-082 34265065 · doi ↗ · pubmed ↗