KPC-33 and ompK37 mutations: Unraveling the mechanism of ceftazidime/avibactam resistance in ST11 carbapenem-resistant Klebsiella pneumoniae
Shulong Zhao, Lin Ye, Shuang Song, Jingfang Sun, Jinfeng Xu, Fei Jiang, Haiquan Kang

TL;DR
This study identifies genetic changes in a type of antibiotic-resistant bacteria that help it resist a specific drug combination, highlighting the importance of genomic monitoring.
Contribution
The study reveals a novel resistance mechanism involving blaKPC-33 and ompK37 mutations in ST11 CRKP.
Findings
The isolate showed high-level resistance to ceftazidime-avibactam and carbapenems.
Resistance was linked to blaKPC-33 and mutations in the ompK37 gene.
The isolate remained susceptible to tigecycline, polymyxin B, and amikacin.
Abstract
Global surveillance indicates rising ceftazidime-avibactam resistance among carbapenem-resistant Klebsiella pneumoniae (CRKP), with sequence type 11 predominating in China. The contribution of blaKPC variants and porin alterations to high-level resistance remains significant. We employed whole-genome sequencing and functional analyses to characterise a CZA-resistant ST11 CRKP isolate recovered from a patient without prior exposure to ceftazidime-avibactam or carbapenems. The isolate harboured blaKPC-33 on a non-conjugative plasmid and multiple non-synonymous mutations in the porin gene ompK37, concomitant with high-level resistance to ceftazidime-avibactam and carbapenems while retaining susceptibility to tigecycline, polymyxin B and amikacin. blaKPC-33 coupled with OmpK37 alterations underpins dual resistance to ceftazidime-avibactam and carbapenems in ST11 CRKP, underscoring the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
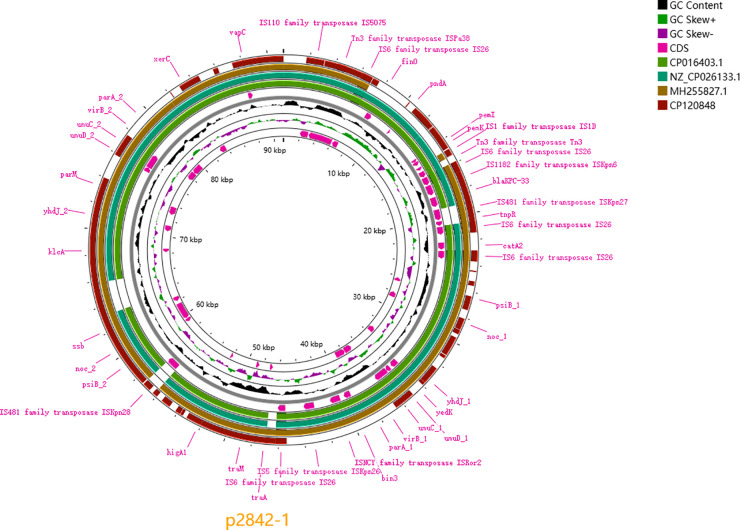 Fig 1
Fig 1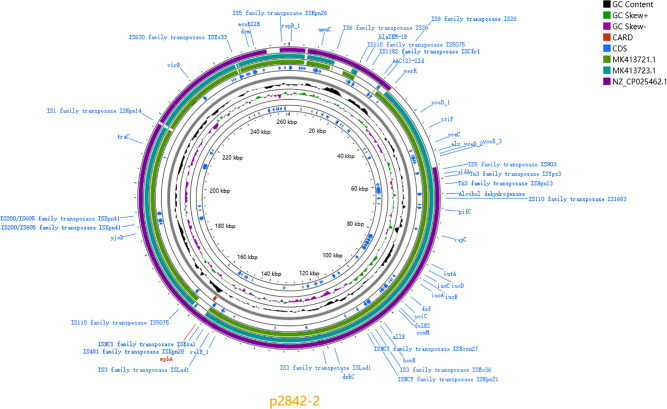 Fig 2
Fig 2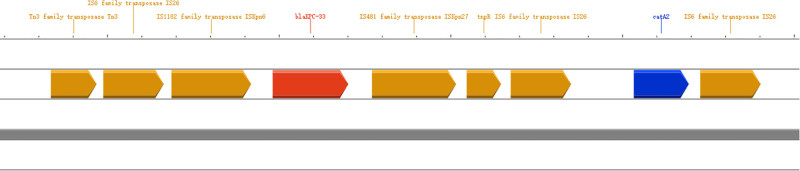 Fig 3
Fig 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Infections and bacterial resistance · Antibiotics Pharmacokinetics and Efficacy
1 Introduction
Carbapenem-resistant Klebsiella pneumoniae (CRKP) producing KPC enzymes have disseminated globally, leaving ceftazidime-avibactam (CZA) as one of the last viable options. Nevertheless, prospective surveillance shows that CZA resistance among CRKP has been on the rise in recent years. This emerging resistance poses a significant threat to clinical treatment outcomes, as it limits the already scarce therapeutic choices for infections caused by CRKP. The mechanisms underlying CZA resistance in CRKP are complex and multifaceted, involving various genetic alterations. Understanding these mechanisms is crucial for developing effective strategies to combat the spread of CZA-resistant CRKP strains. [1,2]. Molecular epidemiology indicates that blaKPC-2/blaKPC-3 variants coupled with porin loss (OmpK35/OmpK37) are the dominant resistance drivers [3]; however, the contribution of the recently described blaKPC-2/blaKPC-33 allele remains under-characterised, especially in China where sequence type 11 (ST11) constitutes the majority of CRKP isolates [4].
BlaKPC-33 differs from blaKPC-2 by a single Asp179Tyr substitution that restores carbapenem hydrolysis while conferring CZA resistance in vitro [5]. Clinical isolates carrying blaKPC-33 have been reported in adult and paediatric patients receiving CZA, suggesting that prior use of CZA may select for this variant [6]. Additionally, cumulative data imply that triple non-synonymous substitutions in OmpK37 (Ile70Met, Ile128Met, Asn230Gly) further reduce CZA uptake and potentiate the resistant phenotype [7]. Despite these insights, no study has integrated whole-genome analysis with detailed MIC data to dissect the synergistic effect of blaKPC-33 and OmpK37 mutations in ST11 CRKP.
Here, we employed hybrid long-read sequencing to characterise a CZA-resistant ST11 CRKP recovered from a patient without previous CZA or carbapenem exposure. Our findings delineate the genetic context of blaKPC-33 and quantify the impact of OmpK37 alterations on antimicrobial susceptibility, providing an evidence base for updated diagnostic and surveillance strategies in China.
2 Materials and methods
2.1 Bacterial isolation and identification
A carbapenemase-producing Klebsiella pneumoniae strain, designated 2842, was isolated from a sputum sample collected in April 2023 from a 62-year-old male patient with severe traumatic brain injury at the Affiliated Hospital of Xuzhou Medical University, Xuzhou, China. The patient had received ceftazidime (2 g q8 h, 9 days) and cefoperazone–tazobactam (2 g q12 h, 5 days) during the preceding two weeks, but no ceftazidime–avibactam or carbapenems. Species identification was performed by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS, Bruker Biotyper)
2.2 Antimicrobial susceptibility testing
MICs were determined by broth microdilution following CLSI M100-34th (2024) and EUCAST v14.0 (2024) breakpoints. Tigecycline MIC was determined by broth microdilution and interpreted using FDA breakpoints.Plates were incubated at 35 °C in ambient air for 16–20 h. Quality control was performed daily with Escherichia coli ATCC 25922 and Klebsiella pneumoniae ATCC 700603; all QC values fell within published ranges.
As only one clinical isolate was analysed, no inferential statistics were applied; MIC values are reported descriptively without median/IQR calculations or inter-group comparisons.
2.3 Whole-genome sequencing and analysis
Genomic DNA was extracted using a commercial kit (Novogene). Illumina MiSeq (2 × 150 bp) and PacBio Sequel II (HiFi) libraries were constructed and sequenced by Novogene (Beijing, China). Hybrid de novo assembly was performed with Unicycler v0.4.8 (default parameters); mean coverages were 220× (Illumina) and 65× (PacBio) for the chromosome, and 180–350× for the plasmids. High-quality reads (mean Phred score Q > 35) were retained to keep base error rate <0.05% [8]. MLST was assigned using the Pasteur scheme (https://bigsdb.pasteur.fr). Plasmid replicons and resistance genes were identified with PlasmidFinder and ResFinder 4.1 (Center for Genomic Epidemiology), respectively. Genome annotation was carried out using the NCBI PGAP pipeline. The complete chromosome and plasmid sequences were deposited under NCBI BioProject PRJNA1161147 (accessions CP170134–CP170136). Raw Illumina and PacBio reads are available in the Sequence Read Archive (SRR35805130,SRR35805131) under the same BioProject;
2.4 Conjugation experiment
A conjugation experiment was conducted to ascertain whether the blaKPC-33 gene was located on a conjugative plasmid. K. pneumoniae 2842 served as the donor, and E. coli J53 as the recipient. Transconjugants were selected on LB agar plates supplemented with sodium azide (100 µg/ml) and meropenem (0.3 µg/ml). The presence of the resistance gene blaKPC-33 in transconjugants was confirmed by PCR, following the methods described in the study.
As a positive control, K. pneumoniae ATCC BAA-1705 carrying blaKPC-2 on an IncFII plasmid was mated with E. coli J53, yielding transconjugants at a frequency of 10 ⁻ ⁵. Heat-killed donor cells (100 °C, 15 min) served as the negative control and produced no growth on selection plates. Strain 2842 gave no transconjugants (<10 ⁻ ⁹), consistent with the absence of tra genes on p2842-1.
2.5 Ethical approval
This study was approved by the Ethics Committee of the Affiliated Hospital of Xuzhou Medical University (XYFY2022-KL008–01) and informed consents were obtained from all patients, available upon written request.
3 Results and discussion
3.1 Resistance profile of K. pneumoniae 2842
Two weeks before the isolate was obtained, the patient had received ceftazidime (2 g q8h, 9 days) and cefoperazone/tazobactam (2 g q12h, 5 days), providing a plausible selective window for blaKPC-33 emergence.
Strain 2842 exhibited high-level resistance to ceftazidime-avibactam (MIC ≥ 32 mg/L), imipenem and meropenem (MIC ≥ 16 mg/L each), while remaining susceptible to tigecycline and polymyxin B (Table 1). A complete MIC panel for 21 antimicrobial agents is provided in S1 File.
Table 1: Minimum inhibitory concentrations (MICs) and interpretive categories of antimicrobial agents against Klebsiella pneumoniae strain 2842.
Although blaKPC-33 typically restores carbapenem susceptibility while conferring CZA resistance [5], 2842 retained full carbapenem resistance, creating a “camouflage” phenotype that can be missed by routine meropenem-based screening. Epidemiological data show that the patient had never received CZA, indicating that ceftazidime or cefoperazone/tazobactam can select blaKPC-33 in vivo—a scenario rarely documented previously.To place the present isolate in context, Table 2.
Table 2: Comparison of recently reported ST11 CRKP isolates exhibiting ceftazidime-avibactam resistance.
3.2 Whole-genome characteristics
Illumina clean reads (11,578,488; 150PE) and PacBio RS reads (120,568; mean 14.9 kb) were assembled into a single chromosome (5,451,441 bp; G + C 57.25%) and two plasmids: p2842-1 (91,212 bp; G + C 53.77%) and p2842-2 (261,134 bp; G + C 47.68%). Annotation (NCBI PGAP) revealed 5,561 CDS, 78 tRNAs, 8 rRNAs, 1 tmRNA and 129 pseudogenes. MLST assigned ST11, a high-risk clone endemic in China.A complete list of resistance determinants and key mutations detected in strain 2842 is provided in Table 3.
Table 3: Resistance determinants detected in Klebsiella pneumoniae 2842.
3.3 Plasmid analysis
p2842-1 (IncFII/IncR) carries blaKPC-33 and catA2 within an IS26-flanked 90.7-kb island (Tn3-IS26-ISKpn6-blaKPC-33-ISKpn27-tnpR-IS26-catA2-IS26) and shares 99.96% identity with pF5/pF5_1 (CP016403). Absence of tra genes concurs with negative conjugation.
p2842-2 (IncFIB/IncHI1B) harbours blaTEM-1B, AAC(3)-IId and mphA and is 99.87% identical to p504051-HI3/p721005-HI3. Both plasmids lack complete conjugative modules (Figs 1-2).The chromosomal context (±50 kb) surrounding the integrated blaKPC-33 module is illustrated in Fig 3.
Circular comparison of p2842-1 (91 kb) carrying blaKPC-33 with pF5/pF5_1, pSH9-KPC and pKPHRJ.Map generated with Proksee.
Circular comparison of p2842-2 (261 kb) carrying aac(3)-IId, mph(A) and blaTEM-1B with p504051-HI3/p721005-HI3 and p44-1.Map generated with Proksee.
The red arrow indicates blaKPC-33; yellow arrows, IS26/ISKpn6 elements; blue arrows, replication/partition proteins; grey shading.The diagram illustrates the chromosomal integration hotspot of the IS26-flanked resistance module. Map was generated with Proksee.
3.4 Discussion
We document a ST11 Klebsiella pneumoniae co-harbouring blaKPC-33 and triple OmpK37 substitutions (I70M+I128M + N230G) in Jiangsu Province, China. The strain displayed “camouflage” resistance—ceftazidime-avibactam (CZA) MIC ≥ 32 mg/L while carbapenem MICs remained ≥16 mg/L, contradicting previous reports that Asp179Tyr restores imipenem susceptibility [6]. This phenotypic inconsistency highlights the necessity of combining porin loss with enzyme characterization when interpreting MIC data.
This combination yields CAZ-AVI MIC ≥ 32 mg/L in the present ST11 isolate, whereas previously reported KPC-33-harbouring strains remained susceptible at 2–8 mg/L [5], demonstrating a phenotypic escalation beyond the enzyme-only effect.Asp179Tyr expands the Ω-loop of KPC-2, enlarging the acyl-enzyme pocket and increasing catalytic efficiency against avibactam 15-fold relative to KPC-2 [15]. The triple OmpK37 substitutions (I70M/I128M/N230G) narrow the porin channel and are associated with reduced outer-membrane permeability to carbapenems, cooperating with KPC-33 to confer ceftazidime-avibactam resistance [16,17], providing a physical barrier that keeps carbapenem MICs elevated even in the presence of a less efficient carbapenemase. Thus, enzyme kinetics plus reduced influx synergistically generate the dual resistance phenotype.
Routine meropenem-based screening failed to flag the isolate as a carbapenemase producer, consistent with recent observations that blaKPC-33 carriers are often missed by automated systems owing to low-level carbapenem resistance phenotypes.We therefore recommend (i) supplementing current algorithms with CZA gradient strips and (ii) adding a blaKPC-33-specific real-time PCR to the national AMR sentinel protocol. In particular, excessive or prolonged use of third-generation cephalosporins (e.g., ceftazidime, cefoperazone) exerts strong selective pressure in vivo and has been associated with a higher risk of colonisation or infection by carbapenem-resistant Enterobacterales, including blaKPC harbourers [18–21]; strict β-lactam stewardship—especially avoiding prolonged monotherapy—should therefore be prioritised in wards where ST11 CRKP is endemic.
Although therapeutic options against CRKP remain scarce, emerging gene-editing tools offer a potential breakthrough for halting the horizontal transfer of resistance determinants. CRISPR-Cas9 systems have been demonstrated to selectively excise blaKPC and blaNDM plasmids, curing carbapenem resistance and blocking conjugative transfer in Enterobacteriaceae [22]. Sublethal antibiotic stress increases IS26 transposition activity, and CRISPR-based or antisense RNA targeting of IS26 transposase can curb resistance island rearrangements in Klebsiella pneumoniae [23]. The IS26-flanked module carried by p2842-1 in the present study (Tn3-IS26-ISKpn6-blaKPC-33-ISKpn27-tnpR-IS26-catA2-IS26) constitutes an ideal target for such interventions. Future studies should verify in vitro (i) the efficacy of these strategies in selectively eliminating KPC-33–OmpK37 dual-resistant Klebsiella pneumoniae and (ii) their impact on the gut microbial ecosystem. Implementing these approaches could open new avenues for the precise control of ST11 CRKP in China.
3.5 Limitations
3.5.1 Single-isolate generalisability.
This study characterised only one clinical isolate, which limits the statistical power to estimate the prevalence of blaKPC-33 plus triple OmpK37 substitutions in China.
Impact: Prevalence figures cannot be extrapolated to provincial or national populations.
Mitigation: We will continue to monitor this evolving resistance pathway through ongoing multi-centre surveillance and targeted sequencing of ST11 CRKP isolates.
3.5.2 Lack of functional complementation.
Permeability and efflux contributions were inferred in silico; isogenic knock-ins or porin electrophoresis were not performed.
Impact: The relative contribution of each OmpK37 substitution to ceftazidime-avibactam MIC remains correlative.
Mitigation: We will conduct targeted experimental work, including allelic exchange and porin electrophoresis, to quantify the individual contribution of each OmpK37 substitution to ceftazidime-avibactam susceptibility.
3.6 Conclusions
This work documents high-level CZA and carbapenem resistance in ST11 K. pneumoniae driven by blaKPC-33 plus OmpK37 triple substitutions on a non-conjugative IncFII/IncR plasmid. The camouflage phenotype underscores the need to include CZA gradient tests or blaKPC-33 PCR in central-China AMR surveillance.
Supporting information
S1 FileMinimum inhibitory concentration (MIC) raw data for 21 antimicrobial agents.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Oliva A, Campogiani L, Savelloni G, Vitale P, Lodi A, Sacco F, et al. Clinical Characteristics and Outcome of Ceftazidime/Avibactam-Resistant Klebsiella pneumoniae Carbapenemase-Producing Klebsiella pneumoniae Infections: A Retrospective, Observational, 2-Center Clinical Study. Open Forum Infect Dis. 2023;10(7):ofad 327. doi: 10.1093/ofid/ofad 327 37476077 PMC 10354859 · doi ↗ · pubmed ↗
- 2Shen S, Tang C, Ding L, Han R, Yin D, Yang W, et al. Identification of KPC-112 from an ST 15 Klebsiella pneumoniae Strain Conferring Resistance to Ceftazidime-Avibactam. m Sphere. 2022;7(6):e 0048722. doi: 10.1128/msphere.00487-22 36374086 PMC 9769832 · doi ↗ · pubmed ↗
- 3Bianco G, Boattini M, Lupo L, Ambretti S, Greco R, Degl’Innocenti L, et al. In vitro activity and genomic characterization of KPC-producing Klebsiella pneumoniae clinical blood culture isolates resistant to ceftazidime/avibactam, meropenem/vaborbactam, imipenem/relebactam: an Italian nationwide multicentre observational study (2022-23). J Antimicrob Chemother. 2025;80(2):583–92. doi: 10.1093/jac/dkae 450 39699187 · doi ↗ · pubmed ↗
- 4Zhang N, Qi L, Liu X, Jin M, Jin Y, Yang X, et al. Clinical and Molecular Characterizations of Carbapenem-Resistant Klebsiella pneumoniae Causing Bloodstream Infection in a Chinese Hospital. Microbiol Spectr. 2022;10(5):e 0169022. doi: 10.1128/spectrum.01690-22 36190403 PMC 9603270 · doi ↗ · pubmed ↗
- 5Lai Y-C, Lin L-W, Lee Y-L. Emergence of ceftazidime-avibactam resistance through in-vivo bla KPC-2 to bla KPC-33 conversion during treatment of ST 11 Klebsiella pneumoniae associated infections. Int J Antimicrob Agents. 2024;64(2):107213. doi: 10.1016/j.ijantimicag.2024.107213 38795929 · doi ↗ · pubmed ↗
- 6Shi Q, Yin D, Han R, Guo Y, Zheng Y, Wu S, et al. Emergence and Recovery of Ceftazidime-avibactam Resistance in bla KPC-33-Harboring Klebsiella pneumoniae Sequence Type 11 Isolates in China. Clin Infect Dis. 2020;71(Suppl 4):S 436–9. doi: 10.1093/cid/ciaa 1521 33367577 · doi ↗ · pubmed ↗
- 7Gong G, Chen Q, Luo J, Wang Y, Li X, Zhang F, et al. Characteristics of a ceftadine/avibatam resistance KPC-33-producing Klebsiella Pneumoniae strain with capsular serotype K 19 belonging to ST 15. J Glob Antimicrob Resist. 2023;35:159–62. doi: 10.1016/j.jgar.2023.09.013 37751846 · doi ↗ · pubmed ↗
- 8Vereecke N, Yoon TB, Luo TL, Corey BW, Lebreton F, Mc Gann PT, et al. An open-source nanopore-only sequencing workflow for analysis of clonal outbreaks delivers short-read level accuracy. J Clin Microbiol. 2025;63(8):e 0066425. doi: 10.1128/jcm.00664-25 40679848 PMC 12345217 · doi ↗ · pubmed ↗